
Abstract
Visceral adipose tissue (VAT) encases mesenteric lymphatic vessels and lymph nodes through which lymph is transported from the intestine and mesentery. Whether mesenteric lymphatics contribute to adipose tissue inflammation and metabolism and insulin resistance is unclear. Here we show that obesity is associated with profound and progressive dysfunction of the mesenteric lymphatic system in mice and humans. We find that lymph from mice and humans consuming a high-fat diet (HFD) stimulates lymphatic vessel growth, leading to the formation of highly branched mesenteric lymphatic vessels that ‘leak’ HFD-lymph into VAT and, thereby, promote insulin resistance. Mesenteric lymphatic dysfunction is regulated by cyclooxygenase (COX)-2 and vascular endothelial growth factor (VEGF)-C–VEGF receptor (R)3 signalling. Lymph-targeted inhibition of COX-2 using a glyceride prodrug approach reverses mesenteric lymphatic dysfunction, visceral obesity and inflammation and restores glycaemic control in mice. Targeting obesity-associated mesenteric lymphatic dysfunction thus represents a potential therapeutic option to treat metabolic disease.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary Information). Data that support the findings of this study are available from the corresponding authors upon request. The exception is lipidomic data, which will be available in the NIH Metabolomics Workbench repository at https://doi.org/10.21228/M8GH59. Source data are provided with this paper.
References
Alitalo, K. The lymphatic vasculature in disease. Nat. Med. 17, 1371–1380 (2011).
Bernier-Latmani, J. & Petrova, T. V. Intestinal lymphatic vasculature: structure, mechanisms and functions. Nat. Rev. Gastroenterol. Hepatol. 14, 510–526 (2017).
Trevaskis, N. L., Kaminskas, L. M. & Porter, C. J. From sewer to saviour—targeting the lymphatic system to promote drug exposure and activity. Nat. Rev. Drug Discov. 14, 781–803 (2015).
Petrova, T. V. & Koh, G. Y. Organ-specific lymphatic vasculature: from development to pathophysiology. J. Exp. Med. 215, 35–49 (2018).
Stacker, S. A. et al. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat. Rev. Cancer 14, 159–172 (2014).
Swartz, M. A. Immunomodulatory roles of lymphatic vessels in cancer progression. Cancer Immunol. Res. 2, 701–707 (2014).
Schwager, S. & Detmar, M. Inflammation and lymphatic function. Front. Immunol. 10, 1016–1028 (2019).
Maisel, K., Sasso, M. S., Potin, L. & Swartz, M. A. Exploiting lymphatic vessels for immunomodulation: rationale, opportunities, and challenges. Adv. Drug Deliv. Rev. 114, 43–59 (2017).
Louveau, A. et al. CNS lymphatic drainage and neuroinflammation are regulated by meningeal lymphatic vasculature. Nat. Neurosci. 21, 1380–1391 (2018).
Louveau, A. et al. Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337–341 (2015).
Da Mesquita, S. et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 560, 185–191 (2018).
Aspelund, A. et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 212, 991–999 (2015).
Ma, Q., Ineichen, B. V., Detmar, M. & Proulx, S. T. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat. Commun. 8, 1434 (2017).
Zhang, F. et al. Lacteal junction zippering protects against diet-induced obesity. Science 361, 599–603 (2018).
Blum, K. S. et al. Chronic high-fat diet impairs collecting lymphatic vessel function in mice. PLoS ONE 9, e94713 (2014).
Hespe, G. E. et al. Exercise training improves obesity-related lymphatic dysfunction. J. Physiol. 594, 4267–4282 (2016).
Torrisi, J. S. et al. Inhibition of Inflammation and iNOS improves lymphatic function in obesity. Sci. Rep. 6, 19817 (2016).
Weitman, E. S. et al. Obesity impairs lymphatic fluid transport and dendritic cell migration to lymph nodes. PLoS ONE 8, e70703 (2013).
Arngrim, N., Simonsen, L., Holst, J. J. & Bülow, J. Reduced adipose tissue lymphatic drainage of macromolecules in obese subjects: a possible link between obesity and local tissue inflammation? Int. J. Obes. 37, 748–750 (2012).
Greene, A. K., Grant, F. D. & Slavin, S. A. Lower-extremity lymphedema and elevated body-mass index. N. Engl. J. Med. 366, 2136–2137 (2012).
Klimontov, V. et al. The relationships between serum levels of adipokines, body fat distribution, and subcutaneous microvasculature in type 2 diabetic subjects. Diabetes 67, 2055-P (2018).
Boyages, J. et al. Liposuction for advanced lymphedema: a multidisciplinary approach for complete reduction of arm and leg swelling. Ann. Surg. Oncol. 22, S1263–S1270 (2015).
Dagenais, S. L. et al. Foxc2 is expressed in developing lymphatic vessels and other tissues associated with lymphedema-distichiasis syndrome. Gene Expr. Patterns 4, 611–619 (2004).
Sawane, M. et al. Apelin inhibits diet-induced obesity by enhancing lymphatic and blood vessel integrity. Diabetes 62, 1970–1980 (2013).
Harvey, N. L. et al. Lymphatic vascular defects promoted by Prox1 haploinsufficiency cause adult-onset obesity. Nat. Genet. 37, 1072–1081 (2005).
Dellinger, M. T., Hunter, R. J., Bernas, M. J., Witte, M. H. & Erickson, R. P. Chy-3 mice are Vegfc haploinsufficient and exhibit defective dermal superficial to deep lymphatic transition and dermal lymphatic hypoplasia. Dev. Dyn. 236, 2346–2355 (2007).
Lee, Y. et al. Enhanced lymphangiogenesis and lymphatic function protects diet-induced obesity and insulin resistance. FASEB J. 33, 662.25 (2019).
Lee, M.-J., Wu, Y. & Fried, S. K. Adipose tissue heterogeneity: implication of depot differences in adipose tissue for obesity complications. Mol. Aspects Med. 34, 1–11 (2013).
Liu, J. et al. Impact of abdominal visceral and subcutaneous adipose tissue on cardiometabolic risk factors: the Jackson Heart Study. J. Clin. Endocrinol. Metab. 95, 5419–5426 (2010).
Raajendiran, A., Tsiloulis, T. & Watt, M. J. Adipose tissue development and the molecular regulation of lipid metabolism. Essays Biochem. 60, 437–450 (2016).
Ji, Y., Sakata, Y. & Tso, P. Nutrient-induced inflammation in the intestine. Curr. Opin. Clin. Nutr. Metab. Care 14, 315–321 (2011).
Scallan, J. P., Hill, M. A. & Davis, M. J. Lymphatic vascular integrity is disrupted in type 2 diabetes due to impaired nitric oxide signaling. Cardiovasc. Res. 107, 89–97 (2015).
Zawieja, S. D. et al. Impairments in the intrinsic contractility of mesenteric collecting lymphatics in a rat model of metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 302, H643–H653 (2012).
Kuan, E. L. et al. Collecting lymphatic vessel permeability facilitates adipose tissue inflammation and distribution of antigen to lymph node-homing adipose tissue dendritic cells. J. Immunol. 194, 5200–5210 (2015).
Koenig, A. & Thaunat, O. Lymphoid neogenesis and tertiary lymphoid organs in transplanted organs. Front. Immunol. 7, 646 (2016).
Ruddle, N. H. Lymphatic vessels and tertiary lymphoid organs. J. Clin. Invest. 124, 953–959 (2014).
Escobedo, N. et al. Restoration of lymphatic function rescues obesity in Prox1-haploinsufficient mice. JCI Insight 1, e85096 (2016).
Donath, M. Y. Targeting inflammation in the treatment of type 2 diabetes: time to start. Nat. Rev. Drug Discov. 13, 465–476 (2014).
Kahn, S. E., Cooper, M. E. & Del Prato, S. Pathophysiology and treatment of type 2 diabetes: perspectives on the past, present, and future. Lancet 383, 1068–1083 (2014).
Su, J.-L. et al. Cyclooxygenase-2 induces EP1- and HER-2/Neu-dependent vascular endothelial growth factor-C up-regulation: a novel mechanism of lymphangiogenesis in lung adenocarcinoma. Cancer Res. 64, 554–564 (2004).
Hosono, K. et al. Roles of prostaglandin E2–EP3/EP4 receptor signaling in the enhancement of lymphangiogenesis during fibroblast growth factor-2-induced granulation formation. Arterioscler. Thromb. Vasc. Biol. 31, 1049–1058 (2011).
El-Bahrawy, H., Hegazy, S., Farrag, W. & Werida, R. Targeting inflammation using celecoxib with glimepiride in the treatment of obese type 2 diabetic Egyptian patients. Int. J. Diabetes Dev. Ctries 37, 97–102 (2017).
Hsieh, P.-S. et al. COX-2-mediated inflammation in fat is crucial for obesity-linked insulin resistance and fatty liver. Obesity 17, 1150–1157 (2009).
Karaman, S. et al. Blockade of VEGF-C and VEGF-D modulates adipose tissue inflammation and improves metabolic parameters under high-fat diet. Mol. Metab. 4, 93–105 (2014).
Karaman, S. et al. Transgenic overexpression of VEGF-C induces weight gain and insulin resistance in mice. Sci. Rep. 6, 31566 (2016).
Han, S. et al. Targeted delivery of a model immunomodulator to the lymphatic system: comparison of alkyl ester versus triglyceride mimetic lipid prodrug strategies. J. Control. Release 177, 1–10 (2014).
Hu, L. et al. Glyceride-mimetic prodrugs incorporating self-immolative spacers promote lymphatic transport, avoid first-pass metabolism, and enhance oral bioavailability. Angew. Chem. Int. Ed. 55, 13700–13705 (2016).
Adams, J. M. et al. Ceramide content is increased in skeletal muscle from obese insulin-resistant humans. Diabetes 53, 25–31 (2004).
Turpin, S. M. et al. Obesity-induced CerS6-dependent C16:0 ceramide production promotes weight gain and glucose intolerance. Cell Metab. 20, 678–686 (2014).
Holland, W. L. et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab. 5, 167–179 (2007).
Boon, J. et al. Ceramides contained in LDL are elevated in type 2 diabetes and promote inflammation and skeletal muscle insulin resistance. Diabetes 62, 401–410 (2013).
Anzai, K. et al. Increased lipid absorption and transport in the small intestine of Zucker obese rats. J. Clin. Biochem. Nutr. 45, 82–85 (2009).
Lu, W. J. et al. The regulation of the lymphatic secretion of glucagon-like peptide-1 (GLP-1) by intestinal absorption of fat and carbohydrate. Am. J. Physiol. Gastrointest. Liver Physiol. 293, G963–G971 (2007).
Miura, S. et al. Increased proliferative response of lymphocytes from intestinal lymph during long chain fatty acid absorption. Immunology 78, 142–146 (1993).
Harvey, N. L. The link between lymphatic function and adipose biology. Ann. NY Acad. Sci. 1131, 82–88 (2008).
Raajendiran, A. et al. Proteome analysis of human adipocytes identifies depot-specific heterogeneity at metabolic control points. Am. J. Physiol.-Endocrinol. Metab. 1, E1068–E1084 (2021).
Escobedo, N. & Oliver, G. The lymphatic vasculature: its role in adipose metabolism and obesity. Cell Metab. 26, 598–609 (2017).
Trevaskis, N. L. et al. Intestinal lymph flow, and lipid and drug transport scale allometrically from pre-clinical species to humans. Front. Physiol. 11, 458 (2020).
Chi, J. et al. Three-dimensional adipose tissue imaging reveals regional variation in beige fat biogenesis and PRDM16-dependent sympathetic neurite density. Cell Metab. 27, 226–236 (2018).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Trevaskis, N. L. et al. The mesenteric lymph duct cannulated rat model: application to the assessment of intestinal lymphatic drug transport. J. Vis. Exp. https://doi.org/10.3791/52389 (2015).
Trevaskis, N. L. et al. A mouse model to evaluate the impact of species, sex, and lipid load on lymphatic drug transport. Pharm. Res. 30, 3254–3270 (2013).
Tsiloulis, T. et al. Impact of endurance exercise training on adipocyte microRNA expression in overweight men. FASEB J. 31, 161–171 (2017).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC(T) method. Methods 25, 402–408 (2001).
Roy, D., Perreault, M. & Marette, A. Insulin stimulation of glucose uptake in skeletal muscles and adipose tissues in vivo is NO dependent. Am. J. Physiol. 274, E692–E699 (1998).
Stone, K. P., Wanders, D., Orgeron, M., Cortez, C. C. & Gettys, T. W. Mechanisms of increased in vivo insulin sensitivity by dietary methionine restriction in mice. Diabetes 63, 3721–3733 (2014).
Krebs buffer (10×, pH 7.2). Cold Spring Harb. Protoc. https://doi.org/10.1101/pdb.rec11255 (2007).
Weigmann, B. et al. Isolation and subsequent analysis of murine lamina propria mononuclear cells from colonic tissue. Nat. Protoc. 2, 2307–2311 (2007).
Nishimura, S. et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 15, 914–920 (2009).
Aurelio, L. et al. From sphingosine kinase to dihydroceramide desaturase: a structure–activity relationship (SAR) study of the enzyme inhibitory and anticancer activity of 4-((4-(4-chlorophenyl)thiazol-2-yl)amino)phenol (SKI-II). J. Med. Chem. 59, 965–984 (2016).
Creek, D. J., Jankevics, A., Burgess, K. E., Breitling, R. & Barrett, M. P. IDEOM: an Excel interface for analysis of LC–MS-based metabolomics data. Bioinformatics 28, 1048–1049 (2012).
Sumner, L. W. et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 3, 211–221 (2007).
Chong, J. et al. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 46, W486–W494 (2018).
Andrikopoulos, S., Blair, A. R., Deluca, N., Fam, B. C. & Proietto, J. Evaluating the glucose tolerance test in mice. Am. J. Physiol. Endocrinol. Metab. 295, E1323–E1332 (2008).
Lancaster, G. I. & Henstridge, D. C. Body composition and metabolic caging analysis in high fat fed mice. J. Vis. Exp. https://doi.org/10.3791/57280 (2018).
Acknowledgements
We thank the Monash Animal Research Platform; the Imaging and FACS core at the Monash Institute of Pharmaceutical Sciences, particularly S. Fung for technical support; the Melbourne Mouse Metabolic Phenotyping Platform, particularly V. Haynes for technical support; T. Karnezis, St. Vincent’s Institute, Melbourne, for providing human adipocytes and T. Tsiloulis, J.C.Y. Lo, D. Senyschyn and G. Chen for technical assistance. We sincerely thank surgeons G. Beban, N. Evenett, P. Johnston and A. Bartlett for their assistance with human mesenteric adipose tissue collection. This work was supported by project grants from the National Health and Medical Research Council of Australia (NHMRC grant 1100036 and NHMRC grant 1177084) and the Health Research Council of New Zealand (HRC 16-036) and the Hugo Charitable Trust (New Zealand).
Author information
Authors and Affiliations
Contributions
E.C. conducted the majority of the experimental work, wrote the main paper and designed and interpreted experiments; N.L.T. was the main supervisor of E.C., conducted some adipocyte studies, wrote the main paper and designed and interpreted the work; C.J.H.P. and M.J.W. substantially revised the paper, co-supervised E.C. and contributed to experimental design and interpretation; M.J.W. also assisted with some metabolic analyses; C.J.N. optimized imaging methods and analysis and revised sections of the manuscript and methods related to imaging analysis; T.Q. synthesized and helped design the celecoxib prodrug and wrote methods for prodrug synthesis; J.S.S. facilitated the design and synthesis of the celecoxib prodrug and revised methods for prodrug synthesis; L.H. co-supervised E.C. and contributed to experimental design and interpretation; S.A., G.G., V.D.M.F., H.C. and A.L. conducted analyses such as ELISA, flow cytometry and metabolic analyses; G.S. conducted light-sheet microscopy analysis of human adipose tissue samples; J.A.W., A.R.J.P., J.H., A.B.J.E. and K.L.P. collected human samples, assisted with data interpretation and revised the manuscript; N.L.H. assisted with design and interpretation of lymphatic analyses and revised the manuscript. A.S., D.J.C. and D.A. conducted the metabolomic analysis, assisted with interpretation and presentation of metabolomic data and revised sections of the manuscript and methods related to the metabolomic analysis.
Corresponding authors
Ethics declarations
Competing interests
C.J.H.P., N.L.T., T.Q., J.S.S., E.C. and M.J.W. are inventors of the lymph-directing glyceride prodrug technology described in the text (PCT/AU2020/050997). This technology has been patented and licensed via a commercial agreement with PureTech Health. PureTech Health has subsequently entered into a collaboration agreement with Boehringer Ingelheim to explore the technology in immune modulation. J.S.S. and T.Q. are currently employed by PureTech Health. The remaining authors declare no competing interests.
Additional information
Peer review information Nature Metabolism thanks Stephan Herzig, Joseph Rutkowski and the other, anonymous, reviewers for their contribution to the peer review of this work. Primary Handling Editor: Christoph Schmitt
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 HFD-associated changes to intestinal villi and lacteals, and adipocytes and FALC in VAT.
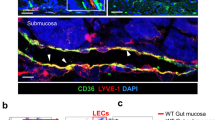
a, Representative immunofluorescence images of intestinal villi. Cell nuclei (Hoechst, grey), CD31 + blood vessels (pink) and LYVE-1+ lacteals (green). Scale bars, 100 µm. b-e, Quantification of the width and length of the intestinal villi and lacteals from immunofluorescence images. f, Representative immunofluorescence images of VAT tissue stained for lipid droplets (green, Bodipy C16 FA). Scale bar, 50 µm. g, Quantification of adipocyte size (µm2). Mean ± s.e.m, n = 4, 5 mice in (b-c), n = 4, 4 mice in (d-e) with 3-4 intestinal villi analysed per mouse, n = 3, 3 mice in (g). Statistical differences, *p < 0.05 from two-tailed Mann-Whitney test. h, Immunofluorescent images of podoplanin + (green) lymphatic vessels in mesenteric adipose tissue from lean patient 2 and 3 and obese patient 2. Images from lean patient 1 and obese patient 2 are in Fig. 1c. (i-m) Representative immunofluorescence images of lymphatic vessels (LYVE-1 + , pink), blood vessels (CD31 + , cyan), CD11b + myeloid cells (grey), CD4 + T cells (yellow), B220 + B cells (blue), and YFP particles (green) in FALCs from mice fed CFD or HFD for 32 weeks. i–j, Type 1 FALCs were rich in CD31 + blood vessels, T cells and B cells but lacked CD11b + and LYVE + cells (that is LECs and macrophages) and had relatively low uptake of YFP-nanoparticles. k, Type 2 FALCs were rich in T cells, B cells and LYVE-1+ cells (macrophages or LEC) but had no observable lymphatic vessels and moderate uptake of YFP-nanoparticles. l-m, Type 3 FALCs contained, and were in close proximity to, LYVE-1+ lymphatic vessels and CD31 + blood vessels. The lymphatic vessels appeared disorganised and lacked a clear orientation toward a collecting lymphatic vessel. Type 3 FALCs also contained high numbers of T cells, B cells and myeloid cells. The observed overlap in staining between the YFP nanoparticles and CD11b + myeloid cells is most likely due to phagocytosis of the particles by the cells. Scale bars, 100 µm (i-m). (i-m) Representative of images from 2-3 FALCs for n = 5 mice. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 2 An examination of mesenteric lymphatic dysfunction in 15-17 week old CFD fed db/db mice in comparison to C57BL/6 mice fed a CFD or HFD for 6, 15, 23 or 32 weeks from 6-7 weeks of age.
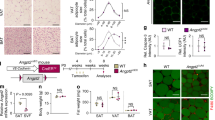
a, Total body weight. Mean ± s.e.m for n = 9, 9, 5. b, Representative immunofluorescence images of the mesenteric lymphatic vessels (LYVE-1 (green)) in VAT of db/db and C57BL/6 mice. Note white arrow indicates the direction of lymph flow, arrow heads indicate point of vessel branching. Scale bar, 500 μm. c, Quantification of mesenteric lymphatic vessel branching in VAT of mice. Mean ± s.e.m for n = 4 or n = 5 mice for week 6, n = 6 or n = 7 mice for week 15, n = 4 or n = 5 mice for week 23, n = 4 or n = 5 mice for week 32 and n = 5 for db/db mice. d, Representative images of Evans blue dye passage through mesenteric lymphatic vessels and leakage to VAT 10 min after intramucosal dye injection to CFD and HFD fed C57BL/6 mice and db/db mice. Scale bar, 5 mm. White circle highlights a site of lymph leakage. e, Area under the curve (AUC) of the Evan’s blue dye intensity plots. Mean ± s.e.m., n = 7, 7 and 5 mice. f, Total cells in mesenteric lymph of mice fed CFD or HFD for 6, 15, 23, 32 weeks or db/db mice fed CFD. Mean ± s.e.m for n = 5 or n = 4 mice for week 6, n = 8 or n = 9 mice for week 15, n = 6 or n = 6 mice for week 32 and n = 3 for db/db mice. g, Weight of subcutaneous adipose tissue (SAT, inguinal adipose tissue depot) and mesenteric adipose tissue (MAT). Note comparisons across CFD, HFD and db/db mice in (b, d, e, g) are made between C57BL/6 mice fed CFD or HFD for 23 weeks and db/db mice age 15-17 weeks. Statistical differences, *p < 0.05, **p < 0.01, ***p < 0.005, or ****p < 0.0001 from (a, e) one-way ANOVA, (c) two-tailed Mann-Whitney test for week 6, 23 and 32 data, two-tailed Student’s t test was used for week 15 data, one-way ANOVA was used for comparison between db/db mice and week 23 and week 32 data, (f) a two-tailed Student’s t-test (except one-way ANOVA was used for comparison between db/db mice vs week 6 data), (g) two-way ANOVA. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 3 Body weight, and composition and effects of mesenteric lymph from Sprague-Dawley rats fed CFD or HFD for 6-9 weeks.
a, Body weight and b, TG concentration. Mean ± s.e.m for n = 4, 4 (HFD) rats. c, Heatmap analysis of lipidomics data of lymph from HFD mice and rats, expressed as mean peak intensity relative to CFD mice and rats, respectively. Blue indicates decreased abundance, red indicates increase and yellow signifies no change. n = 4, 4 mice/rats. d-e, PGE2 and VEGFc concentration in mesenteric lymph of rats. Mean ± s.e.m, n = 4, 4 rats. Statistical differences, *p < 0.05 from two-tailed Mann-Whitney test. f, Real time PCR analysis of Fabp4, Lepr, Gapdh, Pparg, Pnpla2, and Il6 mRNA expression in 3T3-L1 adipocytes treated with control media (Ctrl) or 0.5 or 2% v/v CFD-lymph in media. Mean ± s.e.m. for n = 3 from N = 1 experiment. g, Lipolysis in mature 3T3-L1 adipocytes (from glycerol release) in basal and forskolin-stimulated conditions after treatment with control media (Ctrl) or 2% lymph chylomicron (CM), VLDL or lipid-free protein fraction (supernatant). Mean ± s.e.m. for n = 3 except Ctrl (n = 9 (basal) and n = 6 (forskolin)) from N = 1 experiment. Statistical differences, *p < 0.05, **p < 0.01, ***p < 0.005, ****p < 0.001 from Two-Way ANOVA. h, 14C-2-deoxyglucose (2DG) uptake into SPF3 human adipocytes in basal and insulin-stimulated conditions after treatment with control media or 2% HFD rat lymph. Mean ± s.e.m. for n = 8, 8 (basal) and n = 6, 7 (insulin) replicates from N = 3 experiments. Statistical differences, *p < 0.05 from two-way ANOVA. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 4 The impact of EP4 inhibition on HFD-lymph induced LEC migration, adipocyte lipolysis and adipocyte insulin sensitivity.
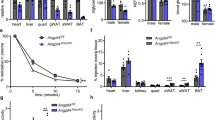
a, LEC migration over time on incubation with control media or 2% v/v HFD-lymph from mice ± EP4 antagonist, L-161,982. Mean ± s.e.m for n = 9, 7, 10 from N = 2 experiments. b, Lipolysis in mature 3T3-L1 adipocytes (from glycerol release) in basal and forskolin-stimulated conditions on incubation with control media or 2% v/v HFD-lymph from mice ± EP4 antagonist. Mean ± s.e.m for n = 8, 8, 8 (basal) and 6, 9, 8 (forskolin) replicates from N = 2 experiments. c, 14C-2-deoxyglucose (2DG) uptake into mature 3T3-L1 adipocytes in basal and insulin-stimulated conditions on incubation with control media or 2% v/v HFD-lymph from mice ± EP4 antagonist. Mean ± s.e.m. for n = 4 (basal) and n = 4 (insulin) replicates of N = 2 experiments. Statistical differences (a) *p < 0.05 from one-way ANOVA and (b-c) *p < 0.05, **p < 0.01 from two-way ANOVA. d-e, PGE2 and VEGFc concentration in plasma of mice fed CFD or HFD for 22-23 weeks. Mean ± s.e.m., n = 6, 6 (plasma VEGFc) and n = 5, 5 (plasma PGE2) mice. Statistical differences, ****p < 0.001 from two-tailed Student’s t-test. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 5 Immune cell accumulation in mesenteric lymph fluid and lymph nodes across prevention and treatment groups.
a-b, Total immune cells in mesenteric lymph fluid and lymph nodes. Mean ± s.e.m., (a) n = 5, 4 (week 6), n = 8, 9 (week 15), n = 6, 6 (week 32). (b) n = 5, 5 (week 6), n = 5, 5 (week 15), n = 6, 6 (week 32). c, Percent of CD45 + cells that were T cells (CD3 + ), B cells (CD19 + ), dendritic cells (CD11c + ), macrophages (F4/80 + ) and NK cells (NK1.1), and d, Percent of T cells that were Th1 (IFNγ + ), Th2 (IL-4 + ) and Th17 (IL-17 + ) cells in mesenteric lymph nodes of mice fed CFD or HFD for 6, 15 or 32 weeks. (c-d) all data at week 15 with n = 5 (CFD), 7 (HFD, except n = 5 for CD11c + and NK1.1+ cells). e, f, Total immune cells in mesenteric lymph fluid or node of mice fed with CFD, HFD or HFD plus Celecoxib (Cele) for 15 weeks (prevention study, see timeline Fig. 4e). Mean ± s.e.m., (e) n = 5, 5, 4. (f) n = 9, 10, 7. g-h, Percent of CD45 + cells that were T cells (CD3 + ), B cells (CD19 + ), dendritic cells (CD11c + ), macrophages (F4/80 + ) and NK cells (NK1.1) in mesenteric lymph fluid or nodes of mice fed with CFD, HFD or HFD plus celecoxib or Cele-Pro (treatment study, see timeline Fig. 5a). Mean ± s.e.m., (g) n = 6, 7, 4, 5 and (h) n = 5, 7, 6, 5 (except n = 4 for CFD NK1.1+ cells, n = 6 for HFD CD3 + cells and n = 6 HFD NK1.1+ cells). Statistical differences from two-tailed Student’s t-test (a-d) or one-way ANOVA (e-h), *p < 0.05, **p < 0.01, ***p < 0.005, ****p < 0.001. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 6 Systemic exposure of celecoxib in mice administered celecoxib (Cele) or celecoxib prodrug (Cele-Pro).
a, Plasma concentrations of celecoxib over time in individual mice, and b, Area under the curve (AUC) of celecoxib plasma concentration versus time profile from time 0-5 h following oral gavage of 29 mg/kg Cele or 29 mg/kg Cele-Pro to fasted mice in a lipid based formulation. Mean of n = 2.
Extended Data Fig. 7 Lipid metabolites in mesenteric lymph fluid obtained from mice fed a CFD, HFD, HFD plus celecoxib or celecoxib prodrug (Cele-Pro).
a-b, PGI2 and 6 keto-PGF1a concentrations in mesenteric lymph from CFD, HFD, HFD + Cele and HFD + Cele-Pro fed mice. Mean ± s.e.m for n = 5, 6, 6, 7. c, Free fatty acid (FFA), (d) cholesterol (Ch), (e) phospholipid (PL) and (f) glucose concentrations in mesenteric lymph. Mean ± s.e.m., n = 4, 8, 6, 7 or 4. g, Lipidomic analysis heatmap of mesenteric lymph from mice in the prevention study (Fig. 4e,g timeline) or treatment study (Fig. 5a timeline). h, Fold change in concentration of significant lipid metabolites in lipidomic analysis including sphingolipids, fatty amides and acyls, glycolipids, sterols, glycerophosphoglycerols, glycerophosphocholines, glycosylphosphatidylinositol, glycerophosphoethanolamines, glycerophosphoinositols in mesenteric lymph from mice in the treatment study (Fig. 5a timeline). h, Mean ± s.e.m. of peak intensities from treatment samples multiplied by the fold change values compared to control. n = 4 (CFD), 7 (HFD), 5 (Cele), 3 (Cele-Pro) mice. P values and details of the statistical testing are provided as source data.
Extended Data Fig. 8 Dose-dependent changes to blood glucose and plasma insulin in mice treated with celecoxib or celecoxib prodrug (Cele-Pro).
Blood glucose (a) and plasma insulin (b) area under the concentration-time curve (AUC) after an oral glucose challenge in mice that were fed with CFD, HFD, HFD + celecoxib (~3, 10, 30 mg/kg) or HFD + Cele-Pro (~3, 10, 30 mg/kg, celecoxib equivalent). Study outline is as described in Fig. 5a. Mean ± s.e.m for CFD (12), HFD (8), HFD + celecoxib (~3 (n = 6), 10 (n = 8), 30 (n = 7) mg/kg) or HFD + Cele-Pro (~3 (n = 7), 10 (n = 6), 30 (n = 8) mg/kg) in (a). CFD (10), HFD (11), HFD + celecoxib (~3 (n = 7), 10 (n = 8), 30 (n = 6) mg/kg) or HFD + Cele-Pro (~3 (n = 7), 10 (n = 6), 30 (n = 8) mg/kg) in (b). Statistical differences, **p < 0.01, ***p < 0.005, or ****p < 0.0001 from one-way ANOVA compared to CFD. P values and details of the statistical testing are provided as source data.
Supplementary information
Supplementary Information
Supplementary Tables 1–5 and Figs. 1–7
Supplementary Video 1
Mesenteric lymphatic vessels in VAT of human patient 1 who was lean.
Supplementary Video 2
Mesenteric lymphatic vessels in VAT of human patient 1 with obesity.
Source data
Source Data Fig. 1
Statistical source data for Fig 1.
Source Data Fig. 2
Statistical source data for Fig 2.
Source Data Fig. 3
Statistical source data for Fig 3.
Source Data Fig. 4
Statistical source data for Fig 4.
Source Data Fig. 5
Statistical source data for Fig 5.
Source Data Fig. 6
Statistical source data for Fig 6.
Source Data Extended Data Fig. 1
Statistical source data for Extended Data Fig 1.
Source Data Extended Data Fig. 2
Statistical source data for Extended Data Fig 2.
Source Data Extended Data Fig. 3
Statistical source data for Extended Data Fig 3.
Source Data Extended Data Fig. 4
Statistical source data for Extended Data Fig 4.
Source Data Extended Data Fig. 5
Statistical source data for Extended Data Fig 5.
Source Data Extended Data Fig. 6
Statistical source data for Extended Data Fig 6.
Source Data Extended Data Fig. 7
Statistical source data for Extended Data Fig 7.
Source Data Extended Data Fig. 8
Statistical source data for Extended Data Fig 8.
Rights and permissions
About this article

Cite this article
Cao, E., Watt, M.J., Nowell, C.J. et al. Mesenteric lymphatic dysfunction promotes insulin resistance and represents a potential treatment target in obesity. Nat Metab 3, 1175–1188 (2021). https://doi.org/10.1038/s42255-021-00457-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-021-00457-w
This article is cited by
-
Lymphatic vessels in the age of cancer immunotherapy
Nature Reviews Cancer (2024)
-
The lymphatic vascular system: much more than just a sewer
Cell & Bioscience (2022)
-
Fixing lymphatics improves glucose metabolism
Nature Metabolism (2021)
-
Lymph leakage promotes insulin resistance in obesity
Nature Reviews Endocrinology (2021)
