
Abstract
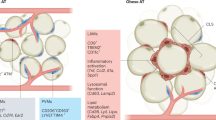
Adipose tissue macrophages (ATMs) display tremendous heterogeneity depending on signals in their local microenvironment and contribute to the pathogenesis of obesity. The phosphoinositide 3-kinase (PI3K) signalling pathway, antagonized by the phosphatase and tensin homologue (PTEN), is important for metabolic responses to obesity. We hypothesized that fluctuations in macrophage-intrinsic PI3K activity via PTEN could alter the trajectory of metabolic disease by driving distinct ATM populations. Using mice harbouring macrophage-specific PTEN deletion or bone marrow chimeras carrying additional PTEN copies, we demonstrate that sustained PI3K activity in macrophages preserves metabolic health in obesity by preventing lipotoxicity. Myeloid PI3K signalling promotes a beneficial ATM population characterized by lipid uptake, catabolism and high expression of the scavenger macrophage receptor with collagenous structure (MARCO). Dual MARCO and myeloid PTEN deficiencies prevent the generation of lipid-buffering ATMs, reversing the beneficial actions of elevated myeloid PI3K activity in metabolic disease. Thus, macrophage-intrinsic PI3K signalling boosts metabolic health by driving ATM programmes associated with MARCO-dependent lipid uptake.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
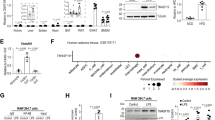

Data availability
The data that support the findings of this study are available from the corresponding authors upon request. The lipidomic datasets generated and analysed during the current study are available at https://github.com/menchelab/Marco. Microarray data of Fig. 5a and Extended Data Fig. 4h that support the findings of this study are available in Gene Expression Omnibus under accession number GSE8831 (ref. 10). Source data are provided with this paper.
Code availability
All code used in the metabolomic differential analyses and subsequent integration is available at https://github.com/menchelab/Marco.
References
Virtue, S. & Vidal-Puig, A. Adipose tissue expandability, lipotoxicity and the Metabolic Syndrome–an allostatic perspective. Biochim. Biophys. Acta 1801, 338–349 (2010).
Lackey, D. E. & Olefsky, J. M. Regulation of metabolism by the innate immune system. Nat. Rev. Endocrinol. 12, 15–28 (2016).
Russo, L. & Lumeng, C. N. Properties and functions of adipose tissue macrophages in obesity. Immunology 155, 407–417 (2018).
Xu, H. et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 112, 1821–1830 (2003).
Arkan, M. C. et al. IKK-beta links inflammation to obesity-induced insulin resistance. Nat. Med. 11, 191–198 (2005).
Jaitin, D. A. et al. Lipid-associated macrophages control metabolic homeostasis in a Trem2-dependent manner. Cell 178, 686–698 (2019).
Hill, D. A. et al. Distinct macrophage populations direct inflammatory versus physiological changes in adipose tissue. Proc. Natl Acad. Sci. USA 115, E5096–E5105 (2018).
Li, C. et al. Single-cell transcriptomics based-MacSpectrum reveals novel macrophage activation signatures in diseases. JCI Insight 5, e126453 (2019).
Kratz, M. et al. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophages. Cell Metab. 20, 614–625 (2014).
Xu, X. et al. Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation. Cell Metab. 18, 816–830 (2013).
Coats, B. R. et al. Metabolically activated adipose tissue macrophages perform detrimental and beneficial functions during diet-induced obesity. Cell Rep. 20, 3149–3161 (2017).
Cinti, S. et al. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 46, 2347–2355 (2005).
Sharif, O., Brunner, J. S., Vogel, A. & Schabbauer, G. Macrophage rewiring by nutrient associated PI3K dependent pathways. Front Immunol. 10, 2002 (2019).
Flaherty, S. E. 3rd et al. A lipase-independent pathway of lipid release and immune modulation by adipocytes. Science 363, 989–993 (2019).
Boutens, L. et al. Unique metabolic activation of adipose tissue macrophages in obesity promotes inflammatory responses. Diabetologia 61, 942–953 (2018).
Saltiel, A. R. & Kahn, C. R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 414, 799–806 (2001).
Winnay, J. N. et al. PI3-kinase mutation linked to insulin and growth factor resistance in vivo. J. Clin. Invest. 126, 1401–1412 (2016).
Foukas, L. C. et al. Critical role for the p110α phosphoinositide-3-OH kinase in growth and metabolic regulation. Nature 441, 366–370 (2006).
Ciraolo, E. et al. Phosphoinositide 3-kinase p110β activity: key role in metabolism and mammary gland cancer but not development. Sci. Signal. 1, ra3 (2008).
Morley, T. S., Xia, J. Y. & Scherer, P. E. Selective enhancement of insulin sensitivity in the mature adipocyte is sufficient for systemic metabolic improvements. Nat. Commun. 6, 7906 (2015).
Horie, Y. et al. Hepatocyte-specific Pten deficiency results in steatohepatitis and hepatocellular carcinomas. J. Clin. Invest. 113, 1774–1783 (2004).
Wijesekara, N. et al. Muscle-specific Pten deletion protects against insulin resistance and diabetes. Mol. Cell. Biol. 25, 1135–1145 (2005).
Wang, L. et al. Deletion of Pten in pancreatic ss-cells protects against deficient β-cell mass and function in mouse models of type 2 diabetes. Diabetes 59, 3117–3126 (2010).
Kosteli, A. et al. Weight loss and lipolysis promote a dynamic immune response in murine adipose tissue. J. Clin. Invest. 120, 3466–3479 (2010).
Lee, Y. S. et al. Inflammation is necessary for long-term but not short-term high-fat diet-induced insulin resistance. Diabetes 60, 2474–2483 (2011).
Ortega-Molina, A. et al. Pten positively regulates brown adipose function, energy expenditure and longevity. Cell Metab. 15, 382–394 (2012).
Cucak, H., Grunnet, L. G. & Rosendahl, A. Accumulation of M1-like macrophages in type 2 diabetic islets is followed by a systemic shift in macrophage polarization. J. Leukoc. Biol. 95, 149–160 (2014).
Boni-Schnetzler, M. & Meier, D. T. Islet inflammation in type 2 diabetes. Semin Immunopathol. 41, 501–513 (2019).
Wang, Y., Viscarra, J., Kim, S. J. & Sul, H. S. Transcriptional regulation of hepatic lipogenesis. Nat. Rev. Mol. Cell Biol. 16, 678–689 (2015).
Schabbauer, G. et al. Myeloid PTEN promotes inflammation but impairs bactericidal activities during murine pneumococcal pneumonia. J. Immunol. 185, 468–476 (2010).
Wentworth, J. M. et al. Pro-inflammatory CD11c+CD206+ adipose tissue macrophages are associated with insulin resistance in human obesity. Diabetes 59, 1648–1656 (2010).
Lancaster, G. I. et al. Evidence that TLR4 is not a receptor for saturated fatty acids but mediates lipid-induced inflammation by reprogramming macrophage metabolism. Cell Metab. 27, 1096–1110 (2018).
Vangapandu, H. V., Jain, N. & Gandhi, V. Duvelisib: a phosphoinositide-3 kinase δ/γ inhibitor for chronic lymphocytic leukemia. Expert Opin. Investig. Drugs 26, 625–632 (2017).
Madisen, L. et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 13, 133–140 (2010).
Moore, K. J. & Freeman, M. W. Scavenger receptors in atherosclerosis: beyond lipid uptake. Arterioscler Thromb. Vasc. Biol. 26, 1702–1711 (2006).
Canton, J., Neculai, D. & Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 13, 621–634 (2013).
Martin, D. et al. Regulation of heme oxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. J. Biol. Chem. 279, 8919–8929 (2004).
Ren, D. et al. Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc. Natl Acad. Sci. USA 108, 1433–1438 (2011).
Patrucco, E. et al. PI3Kγ modulates the cardiac response to chronic pressure overload by distinct kinase-dependent and -independent effects. Cell 118, 375–387 (2004).
Huang, S. C. et al. Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nat. Immunol. 15, 846–855 (2014).
Chavez, J. A. & Summers, S. A. A ceramide-centric view of insulin resistance. Cell Metab. 15, 585–594 (2012).
Lumeng, C. N., Bodzin, J. L. & Saltiel, A. R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest. 117, 175–184 (2007).
Fujisaka, S. et al. Regulatory mechanisms for adipose tissue M1 and M2 macrophages in diet-induced obese mice. Diabetes 58, 2574–2582 (2009).
Persson, J., Nilsson, J. & Lindholm, M. W. Interleukin-1beta and tumour necrosis factor-alpha impede neutral lipid turnover in macrophage-derived foam cells. BMC Immunol. 9, 70 (2008).
Maya-Monteiro, C. M. et al. Leptin induces macrophage lipid body formation by a phosphatidylinositol 3-kinase- and mammalian target of rapamycin-dependent mechanism. J. Biol. Chem. 283, 2203–2210 (2008).
Pal, A. et al. PTEN mutations as a cause of constitutive insulin sensitivity and obesity. N. Engl. J. Med. 367, 1002–1011 (2012).
Dror, E. et al. Postprandial macrophage-derived IL-1β stimulates insulin, and both synergistically promote glucose disposal and inflammation. Nat. Immunol. 18, 283–292 (2017).
Mauer, J. et al. Myeloid cell-restricted insulin receptor deficiency protects against obesity-induced inflammation and systemic insulin resistance. PLoS Genet. 6, e1000938 (2010).
Ortega-Molina, A. et al. Pharmacological inhibition of PI3K reduces adiposity and metabolic syndrome in obese mice and rhesus monkeys. Cell Metab. 21, 558–570 (2015).
Han, S. et al. Macrophage insulin receptor deficiency increases ER stress-induced apoptosis and necrotic core formation in advanced atherosclerotic lesions. Cell Metab. 3, 257–266 (2006).
Liang, C. P. et al. Increased CD36 protein as a response to defective insulin signaling in macrophages. J. Clin. Invest. 113, 764–773 (2004).
Kzhyshkowska, J., Neyen, C. & Gordon, S. Role of macrophage scavenger receptors in atherosclerosis. Immunobiology 217, 492–502 (2012).
Arredouani, M. et al. The scavenger receptor MARCO is required for lung defense against pneumococcal pneumonia and inhaled particles. J. Exp. Med. 200, 267–272 (2004).
Dahl, M. et al. Protection against inhaled oxidants through scavenging of oxidized lipids by macrophage receptors MARCO and SR-AI/II. J. Clin. Invest. 117, 757–764 (2007).
Yoshimatsu, M. et al. Induction of macrophage scavenger receptor MARCO in nonalcoholic steatohepatitis indicates possible involvement of endotoxin in its pathogenic process. Int. J. Exp. Pathol. 85, 335–343 (2004).
Cavallari, J. F. et al. Targeting macrophage scavenger receptor 1 promotes insulin resistance in obese male mice. Physiol. Rep. 6, e13930 (2018).
Hirano, S. & Kanno, S. Macrophage receptor with collagenous structure (MARCO) is processed by either macropinocytosis or endocytosis-autophagy pathway. PLoS ONE 10, e0142062 (2015).
Rogers, N. J. et al. A defect in Marco expression contributes to systemic lupus erythematosus development via failure to clear apoptotic cells. J. Immunol. 182, 1982–1990 (2009).
Wermeling, F. et al. Class A scavenger receptors regulate tolerance against apoptotic cells, and autoantibodies against these receptors are predictive of systemic lupus. J. Exp. Med. 204, 2259–2265 (2007).
Jung, S. B. et al. Reduced oxidative capacity in macrophages results in systemic insulin resistance. Nat. Commun. 9, 1551 (2018).
Mukhopadhyay, S. et al. MARCO, an innate activation marker of macrophages, is a class A scavenger receptor for Neisseria meningitidis. Eur. J. Immunol. 36, 940–949 (2006).
Itoh, K. et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 236, 313–322 (1997).
Brandon, A. E. et al. Minimal impact of age and housing temperature on the metabolic phenotype of Acc2−/− mice. J. Endocrinol. 228, 127–134 (2016).
Ayala, J. E., Bracy, D. P., McGuinness, O. P. & Wasserman, D. H. Considerations in the design of hyperinsulinemic-euglycemic clamps in the conscious mouse. Diabetes 55, 390–397 (2006).
Krzywinski, M. et al. Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645 (2009).
Acknowledgements
We thank H. Paar, A. Hladik and M. Salzmann for technical assistance. We thank E. Hirsch (Department of Molecular Biotechnology and Health Sciences, Molecular Biotechnology Center, University of Torino) for the Pi3kγKD/KD mice, M. Serrano (IRB Barcelona, Barcelona Institute of Science and Technology) for the PtenTg/+ mice and C. Tsatsanis (Laboratory of Clinical Chemistry, University of Crete Medical School) for the Akt1loxP/loxPAkt2 loxP/loxPLysM-Cre+/− mice. This research was supported by FWF 30026 and 31106 to G.S. and 31568 to O.S. and the Christian Doppler Laboratory for Arginine Metabolism in Rheumatoid Arthritis and Multiple Sclerosis to G.S. J.S.B., A.V. and A.L. were supported by a DOC fellowship of the Austrian Academy of Sciences. A.B. received funding from the European Research Council under the European Union’s Seventh Framework Programme and Horizon 2020 research and innovation programme (grant agreement no. 677006; CMIL). M.A.F. is supported by a Senior Principal Research Fellowship from the National Health and Medical Research Council of Australia (APP1116936).
Author information
Authors and Affiliations
Contributions
J.S.B., A.V., O.S. and G.S. conceived and designed the study. J.S.B., A.V., A.L., A.K., M.P., M.H., A.H., M. Kieler, L.Q.G., M. Kerndl, M. Kuttke, A.E.B., E.E. and C.L.E. performed experiments. M.C. analysed lipidomic data and performed bioinformatic analysis. I.M. analysed liver steatosis. M.W.G., M. Kulik, P.M.D. and M.S. provided intellectual input. K.K. performed lipidomics analysis. J.S.B., A.V. and A.L. analysed the data. F.G., J.M., A.B., T.W. and M.A.F. provided key resources. J.S.B., A.V., O.S. and G.S. wrote the manuscript. All authors read, revised and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Primary Handling Editors: Pooja Jha; George Caputa. Nature Metabolism thanks Yi-bin Feng and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Disruptions of adipose homeostasis and metabolic stress ablate myeloid PI3K/AKT signalling.
a, Serum insulin levels of animals subjected to ST-HFD (n = 4 ND, n = 5 ST-HFD animals), 16 weeks HFD (n = 5 ND, n = 4 HFD animals) or acute lipolysis (n = 6 animals per group) (***P = 0.0006; two-tailed student’s t-test). b, Percentage of viable FB (CD45+CD11b+F4/80+) macrophages and in the adipose tissue of mice post 2 or 16 weeks HFD feeding and acute lipolysis (n = 5 animals; **P = 0.0072, ****P < 0.001, **P = 0.0026; two-tailed student’s t-test). c, Corresponding p-AKT MFI of FB-ATMs shown in (b) (*P = 0.027, ***P = 0.0001, **P = 0.0029; two-tailed student’s t-test). d, p-AKT MFI of macrophages post adipocyte conditioned media (ACM) treatment for 3 h (n = 5 samples; ***P = 0.001; two-tailed student’s t-test). Data are mean ± SEM and representative of one (a) or two (b-d) independent experiments.
Extended Data Fig. 2 Adaptive immune cell PTEN and p-AKT, hyperinsulinemic-euglycemic clamps, pancreatic macrophages and weights of BMT animals.
a, PTEN and p-AKT expression in adaptive immune cells from blood of WT (n = 3) or PtenTg/+ (n = 2) animals. Cells were pre-gated on viable CD45+CD3+B220+. b, Weights, fat and lean mass of Pten+/+>WT (n = 12) or PtenTg/+>WT (n = 10) mice as determined using Echo MRI. c, Blood glucose levels and glucose infusion rate during clamping of Pten+/+>WT (n = 10) or PtenTg/+>WT (n = 8) mice. d, Basal and insulin-stimulated whole-body glucose disposal rates and hepatic glucose production of mice in (c). e, Pancreatic CD68 expression in Pten+/+>WT or PtenTg/+>WT 16 weeks post HFD (n = 9 animals per genotype; ***P = 0.0003; two-tailed student’s t-test). f, Weights of mice shown in Fig. 1f-i. Data are mean ± SEM and representative of one (a-e) or two (f) independent experiments.
Extended Data Fig. 3 In vivo effects of modulating myeloid PI3K activity are independent of adiposity differences.
a, PTEN and p-AKT expression in adaptive immune cells from blood of PtenWT (n = 7) or PtenΔmyel (n = 6) mice. Cells were gated on viable CD45+CD3+B220+. Isotype control samples depicted are identical to Extended Data Fig. 2a. b, oGTT and AUC of PtenWT (n = 7 ND, n = 16 HFD) or PtenΔmyel (n = 9 ND, n = 16 HFD) mice, 16 weeks post feeding (**P = 0.0057, **P = 0.0031; two-way ANOVA, Bonferroni’s correction, **P for AUC = 0.009; two-tailed student’s t-test). c, Serum insulin levels post glucose bolus (n = 9 PtenWT, n = 7 PtenΔmyel animals). d, Weights of HFD mice shown in Fig. 2b,c stratified for low weight (LW) and high weight (HW) (n = 4 PtenWT LW, n = 10 PtenWT HW, n = 8 PtenΔmyel LW, n = 9 PtenΔmyel HW). e, Insulin tolerance test (ITT) and area under the curve (AUC) post 16 weeks feeding of PtenWT (n = 4 LW, n = 10 HW) or PtenΔmyel (n = 8 LW, n = 9 HW) mice stratified for low weight (LW) and high weight (HW) (*P = 0.0216, *P = 0.0151, **P = 0.0037; two-way ANOVA, Bonferroni’s correction; **P for AUC = 0.0051; two-tailed student’s t-test). f, Serum AST and ALT levels post 16 weeks HFD feeding of PtenWT (n = 12 HFD, n = 4 LW, n = 8 HW) or PtenΔmyel (n = 10 HFD, n = 4 LW, n = 6 HW) mice stratified for low weight (LW) and high weight (HW) (*P = 0.0185, *P = 0.0126, *P = 0.0293; two-tailed student’s t-test). Data are mean ± SEM and representative of one (a, c) or are pooled from two (d-f) or three (b) independent experiments.
Extended Data Fig. 4 Gating strategies and FBC206-ATMs are a large, lipid laden, lysotracker, TREM-2high population, mostly of resident origin post ST-HFD.
a, Gating strategy of ATMs. b, Gating strategy of monocytes. c, Gating strategy of hepatic myeloid-cells. d, Representative heatmap of BODIPY MFI in relation to cell size (FSC) of cells isolated from eWAT post 16 weeks HFD. e, Representative percentages of FBC and FBC206 populations within FSC low, intermediate and high low cells are depicted. Cells were pre-gated on live CD45+CD11b+F4/80+. f, Cell size quantification according to forward scatter (FSC) of FB-, FBC- and FBC206-ATMs (n = 8 animals; ***P = 0.0001, ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). g, TREM-2 and CD9 MFI of FB-, FBC- and FBC206-ATMs post ST-HFD (n = 5 animals; ****P < 0.0001, *P = 0.013; one-way ANOVA, Bonferroni’s correction). h, Lysosomal gene expression based on KEGG pathway (mmu04142) of microarray data of obese FBs vs FBCs from C57Bl/6 J Lepob/ob mice (GSE8831). Data depicted as fold change relative to FB condition. i, Lysotracker MFI of the FBC206-ATM population post 16 weeks HFD (n = 7 animals; **P = 0.011; one-way ANOVA, Bonferroni’s correction). j, Post ST-HFD, percentages of GFP- FBC- vs GFP+ FBC-ATMs and GFP- FBC206- vs GFP+ FBC206-ATMs were analysed in eWAT of Ccr2GFP/+ animals. Cells were pre-gated on CD45+CD11b+F4/80+ (n = 5 animals; ****P < 0.0001; two-way ANOVA, Bonferroni’s correction). Data are mean ± SEM and representative of one (g, i), two (j) or four (d-f) independent experiments.
Extended Data Fig. 5 FBC206 generation of naïve macrophages stimulated with the indicated cytokines, adipokines and lipids.
a, FBC206/FBC ratio of macrophages post stimulation with glucose, insulin and ACM for 24 hours (n = 5 samples; ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). b, FBC206/FBC ratio of macrophages post stimulation with various cytokines and ACM for 24 hours (n = 3 samples; ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). c, FBC206/FBC ratio of macrophages post stimulation with various lipids and ACM for 24 hours (n = 4 samples; ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). d, FBC206/FBC ratio of macrophages post stimulation with glucose, insulin and MMe for 24 hours (n = 5 samples). e, FBC206/FBC ratio and BODIPY MFI of naïve macrophages treated with Resatorvid and ACM for 24 hours (n = 4 samples). Data are mean ± SEM and representative of two (a-e) independent experiments.
Extended Data Fig. 6 MARCO and p-AKT post ST-HFD and acute lipolysis in the different ATM populations, where PTEN is deleted equally.
a, Percentages of Tomato+ and PTEN+ ATMs represented as percentage of their respective parent (FB-, FBC- or FBC206-ATMs) post ST-HFD in PtenWTR26tdTomato (cre-) and PtenΔmyelR26tdTomato (cre+) mice (n = 3 animals). b/c, MARCO MFI in macrophages post ST-HFD (b) or acute lipolysis (c). Cells were pre-gated on viable FBs (CD45+CD11b+F4/80+) (n = 5 animals; ****P < 0.0001, ***P = 0.0003; one-way ANOVA, Bonferroni’s correction). d, p-AKT MFI of FBC- and FBC206-ATMs of mice post ST-HFD or after acute lipolysis (n = 5 ST-HFD, n = 4 CL316,243 animals; **P = 0.0025, ***P = 0.0001; two-tailed student’s t-test). Data are mean ± SEM and representative of one (a) or two (b-d) independent experiments.
Extended Data Fig. 7 Impaired lipid buffering of Marco-/- mice is not associated with adiposity differences and weights of animals in Fig. 8d,e.
a, Weight of mice in Fig. 6c. b, Fat depot distribution of lean Marco+/+ and Marcon mice presented as percent of bodyweight (n = 4 mice per genotype). RP: retroperitoneal, eWAT: epididymal, VAT: visceral, SAT: subcutaneous, white adipose tissue depots. c, Marco transcript levels measured in macrophages (n = 5 samples) and adipocytes (n = 4 samples). d, NEFA and glycerol levels of adipose tissue isolated from lean Marco+/+ and Marco-/- mice, stimulated with isoproterenol (ISO) for 2 hours (n = 5 animals; **P = 0.0022, **P = 0.0099, ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). e, Weight of mice in Fig. 6d. f, Fat mass and adipose depot distribution of Marco+/+ and Marco-/- mice post 16 weeks HFD. RP: retroperitoneal, eWAT: epididymal, VAT: visceral, SAT: subcutaneous, white adipose tissue depots (n = 11 mice per genotype). g, Weights of mice shown in Fig. 6e. h, Macrophage content and FBC206/FBC ratios of Marco+/+ (n = 5) and Marco-/- (n = 7) mice prior to HFD feeding. i, Corresponding weights of mice in Fig. 8d (***P = 0.0009, ****P < 0.0001; one-way ANOVA, Bonferroni’s correction). j, Corresponding weights of mice in Fig. 8e (*P = 0.0181; one-way ANOVA, Bonferroni’s correction). Data are mean ± SEM and representative of one (b, c, g, h), two (a, d), or are pooled from two (f, j), four (e) or six (i) independent experiments.
Extended Data Fig. 8 PI3K-dependent differences in lipid uptake and catabolism are independent of mitochondrial numbers and mitochondrial respiratory chain complex expression of Marco-/- macrophages.
a, AcLDL uptake of naïve macrophages from PtenWT (n = 10) and PtenΔmyel (n = 13) mice depicted as fold change (FC) increase (**P = 0.0025; two-tailed student’s t-test). b, Spare respiratory capacity (SRC) of data in Fig. 7e (*P = 0.0184; two-tailed student’s t-test). c, Mitochondrial numbers in naïve PtenWT and PtenΔmyel macrophages as determined using Mitotracker (n = 4 samples per genotype). d, mRNA levels of Pten and selected mitochondrial marker genes in naïve PtenWT (n = 8, for Pparg n = 7 samples) and PtenΔmyel (n = 8 samples) macrophages (****P < 0.0001, **P = 0.0091, *P = 0.443; two-tailed student’s t-test). e, Extracellular acidification rate (ECAR) of data in Fig. 7e (***P = 0.0007; two-tailed student’s t-test). f, SRC and ECAR of data in Fig. 7f (two-tailed student’s t-test). g, SRC of data in Fig. 7g (*P = 0.0487, *P = 0.0173, *P = 0.0111; two-tailed student’s t-test). h, SRC of data in Fig. 7h (**P = 0.0063, *P = 0.0171; two-tailed student’s t-test). i, mRNA levels of selected mitochondrial complex genes in naïve Marco+/+ (n = 8 samples) and Marco-/- (n = 9 samples) macrophages. The mitochondrial complex (C) represented by each gene is indicated. Data are mean ± SEM and representative of two (b-h), or are pooled from two (a, i) independent experiments.
Supplementary information
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Rights and permissions
About this article

Cite this article
Brunner, J.S., Vogel, A., Lercher, A. et al. The PI3K pathway preserves metabolic health through MARCO-dependent lipid uptake by adipose tissue macrophages. Nat Metab 2, 1427–1442 (2020). https://doi.org/10.1038/s42255-020-00311-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-020-00311-5
This article is cited by
-
The application of MARCO for immune regulation and treatment
Molecular Biology Reports (2024)
-
Aqueous extract of Platycodon grandiflorus attenuates lipopolysaccharide-induced apoptosis and inflammatory cell infiltration in mouse lungs by inhibiting PI3K/Akt signaling
Chinese Medicine (2023)
-
Targeting macrophages in hematological malignancies: recent advances and future directions
Journal of Hematology & Oncology (2022)
-
Early macrophage response to obesity encompasses Interferon Regulatory Factor 5 regulated mitochondrial architecture remodelling
Nature Communications (2022)
-
Adipoclast: a multinucleated fat-eating macrophage
BMC Biology (2021)
