
Abstract
Habitat selection studies facilitate assessing and predicting species distributions and habitat connectivity, but habitat selection can vary temporally and among individuals, which is often ignored. We used GPS telemetry data from 96 Gray wolves (Canis lupus) in the western Great Lakes region of the USA to assess differences in habitat selection while wolves exhibited resident (territorial) or non-resident (dispersing or floating) movements and discuss implications for habitat connectivity. We used a step-selection function (SSF) to assess habitat selection by wolves exhibiting resident or non-resident movements, and modeled circuit connectivity throughout the western Great Lakes region. Wolves selected for natural land cover and against areas with high road densities, with no differences in selection among wolves when resident, dispersing, or floating. Similar habitat selection between resident and non-resident wolves may be due to similarity in environmental conditions, when non-resident movements occur largely within established wolf range rather than near the periphery or beyond the species range. Alternatively, non-resident wolves may travel through occupied territories because higher food availability or lower human disturbance outweighs risks posed by conspecifics. Finally, an absence of differences in habitat selection between resident and non-resident wolf movements may be due to other unknown reasons. We recommend considering context-dependency when evaluating differences in movements and habitat use between resident and non-resident individuals. Our results also provide independent validation of a previous species distribution model and connectivity analysis suggesting most potential wolf habitat in the western Great Lakes region is occupied, with limited connectivity to unoccupied habitat.
Similar content being viewed by others

Introduction
Understanding how animals select habitat is necessary to explain and predict species distributions, facilitating population management and species conservation1,2. Characterizing species-habitat relationships can inform where populations can establish3 and identify linkages between habitat patches suitable for dispersal4. Whereas habitat selection and associated connectivity studies are valuable2,4, processes underlying habitat selection are often poorly understood5. Drivers of habitat selection can differ among life stages or individuals and understanding these differences can improve our understanding of habitat selection and connectivity5,6.
Mismatches between landscape connectivity analyses and species ecology can be mitigated by accounting for behavioral aspects that can influence movement6,7. Processes underlying animal movements are relevant to connectivity analyses as they influence the behavior and movements of dispersing animals8. Human landscape disturbances, including high human population densities and associated activities, can alter large carnivore movements9,10, but avoidance of human disturbance can be lower for non-resident individuals than for residents11. This can affect accuracy of connectivity models, as studies generally consider habitat selection across individuals as equivalent12,13.
Reduced avoidance of human disturbance by non-resident animals has been documented for dispersing red wolves (Canis rufus;14) and lions (Panthera leo;15), which avoided areas near roads and with higher human population densities less strongly than residents. Non-resident gray wolves (C. lupus) similarly displayed reduced selection against human disturbance compared to resident wolves16,17. Alternatively, dispersers may not avoid areas of higher human disturbance at all; resident brown bears (Ursus arctos) avoided public roads and resident Iberian lynx (Lynx pardinus) avoided low-traffic roads, while dispersers did not18,19.
Gray wolves historically occupied the Northern Hemisphere north of 11–20° N, though by 1970 wolves were extirpated from most of their historical range in the contiguous USA20. Following federal protection in 1974, wolves recolonized additional areas of Minnesota, and former range in Wisconsin21 and the Upper Peninsula of Michigan22. The western Great Lakes population appears to have stabilized at around 4200 wolves23. Unoccupied habitat within former wolf range has been identified in the eastern USA, with apparent limited connectivity to current wolf range in the Great Lakes region24. However, estimates of habitat availability and connectivity should further consider factors underlying habitat selection including potential differences between resident and non-resident movements.
Gray wolves are territorial, though most disperse from their natal territory and establish or become residents of different territories25. Other wolves do not establish new territories or join existing territories and exhibit nomadic (or floating) movements, constrained by conspecific territories20,26. Wolves also make extraterritorial excursions (i.e., predispersal movements) of varying distance and duration27,28. Greater use of human-disturbed areas by wolves when dispersing or floating could result from avoiding existing wolf territories in less human-disturbed areas or decreased site familiarity that reduces their ability to avoid human disturbances, compared to when they are resident of a territory8,29. Alternatively, disturbances such as roads may facilitate efficient travel for non-residents30, while areas with high livestock abundance may provide food when lower site familiarity or prey abundance limits acquisition of wild prey29.
We investigated habitat use by gray wolves in the western Great Lakes region exhibiting resident (territorial) or non-resident (dispersing or floating) movements relative to human disturbance. We predicted wolves would select for areas with greater natural land cover and against areas of greater human disturbance as indexed by road densities and proportions of agricultural land cover, with stronger selection during resident than non-resident movements. We also quantified habitat selection and connectivity throughout the western Great Lakes region and evaluated these results against an existing connectivity map for wolves in the eastern USA. We expected a strong correlation between a previous connectivity map developed using winter track surveys24 and one resulting from this habitat selection analysis based on telemetry data.
Methods
Study area
The study area (Fig. 1) included the area representing the western Great Lakes distinct population segment of gray wolves (hereafter, western Great Lakes region;31), including Minnesota (220,185 km2), Wisconsin (145,593 km2), and Michigan (151,279 km2), and parts of North Dakota (108,193 km2), South Dakota (93,571 km2), Iowa (99,971 km2), and Illinois (27,190 km2).

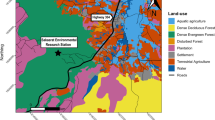
Top panel: land cover within the western Great Lakes distinct population segment of gray wolves (Canis lupus), USA and southern Ontario and Manitoba, Canada. Bottom panel: Circuit connectivity for the western Great Lakes distinct population segment of gray wolves (Canis lupus), USA and southern Ontario and Manitoba, Canada, 2017–2021. Figure based on a step-selection function (main figure) and circuit connectivity map derived from the same study area based on snow track data (24; inset). Figures were created using ArcGIS Pro 3.0.0 (https://www.esri.com).
The study area also included southern Ontario, Canada (515,966 km2), delineated by the Area of the Undertaking (the area in Ontario under forest management;32), and southern Manitoba (84,920 km2). The climate is predominantly humid continental, with warm summers and cold winters33. Average summer (June–September) minima are 7–17 ° C and maxima are 17–30 °C while average winter (December–March) minima are – 25 to − 6 °C and maxima are – 10 to 4 °C34. Elevations are 30–757 m above sea level35. The study area, excluding the Great Lakes, contains 46% natural land cover (primarily various forest types and wetlands) and 18% water, while agricultural and urban areas comprise 32% and 4%, respectively36. The primary prey of wolves in the study area is white-tailed deer (Odocoileus virginianus), in addition to American beaver (Castor canadensis), moose (Alces alces, where available), and other mammals37. Average wolf mid-winter pack size across Minnesota, Wisconsin, and Michigan is 2.7–5.6 individuals38,39,40, and the combined population is about 4200 wolves23.
Data collection and processing
We used gray wolf GPS telemetry data collected during 2017–2021 by state, federal, and tribal agencies of Minnesota, Wisconsin, and Michigan. Animal capture and handling for data collection during original research and monitoring were approved by the respective state, tribal, and federal agencies. Use of these data for this study was approved by the Michigan State University Institutional Animal Care and Use Committee. We excluded the first five days of post-capture data from each wolf to reduce potential capture effects3. We created a dataset of 96 wolves (51 males, 39 females, 6 unknown) collared in Michigan (44), Wisconsin (31), and Minnesota (21), with 13- or 16-h relocation intervals. We compared two preliminary models using datasets with 13- or 16-h relocation intervals, found no notable differences, and pooled these datasets for analysis.
We separated resident (territorial) from non-resident (dispersing or floating) annual wolf movements by calculating relative net squared displacement (rNSD), which represents the squared Euclidian distance between consecutive locations41. For each wolf during each biological year (starting 15 April), we used the MigrateR package42 in program R43 to fit data to three a priori non-linear models representing resident, dispersing, and floating (named ‘nomadic’ in the MigrateR package) movements41. The rNSD assigns data to the model with the most similar net-squared displacement curve. For example, a curve whereby rNSD values are low and stable, exponentially increase, and thereafter stabilize at high rNSD values is classified as a disperser, as this is a curve typical of an animal that was resident to a territory, then dispersed, and thereafter settled into a new territory with a high relative distance from the first location in the dataset. After fitting data to each of the a priori curves, the best supported movement type is assigned based on the lowest AIC score44.
When data could not be assigned to a movement type because wolves displayed multiple movement types within a biological year, we split data between multiple movement categories based on visual inspection of the rNSD-plots and raw GPS data42. Visual inspection of movement data to confirm rNSD classifications is recommended to override rNSD classifications when suspected to be incorrect42. We then calculated 90% bivariate normal kernel utilization distributions to approximate annual range size using the ‘kernelUD’ function in AdehabitatHR package45. State reports during 2017–2021 were used as an independent source to set the maximum annual range size for wolves to be considered resident: from these reports we extracted the maximum territory size (561 km2) found across Minnesota, Wisconsin, and Michigan46. We classified movements within annual ranges ≤ 561 km2 as resident and reclassified movements within larger annual ranges initially classified as resident movements as floating movements. We classified extraterritorial movements between a territory and a non-overlapping territory as dispersal movements, from the first movement beyond the initial territory to the last movement before entering the subsequent territory. We included potential predispersal movements27,28 as dispersal movements by including extraterritorial movements leaving from and returning to the same territory, with a duration ≥ 10 days, based on visual inspection.
Modeling landscape use
We used step-selection functions (SSF), linking consecutive animal locations and contrasting each observed step with three random available steps47. To obtain random steps, we pooled individual movements by movement type47 then randomly sampled the length and angle of random steps from the distribution of observed steps for each type. We used road density, proportion of natural land cover, and proportion of agricultural cover as continuous variables, whereby proportional land covers were calculated as the percentage of respective land cover types within a cell. We achieved this by assigning the value ‘1’ to the land cover of interest and ‘0’ to other land covers at the original raster resolution (30-m), after which we summed all values in an aggregated raster with 300-m resolution. We used road data from TIGER/line shapefiles (50-m resolution;48) and the Canadian National Road Network (5-m resolution;49), the most comprehensive road databases for these countries including categories ranging from highways to service roads, and roads only accessible by four-wheel drive vehicles. We used the North American land change monitoring system (NALCMS; 30-m resolution)36 to calculate proportional land cover. We reclassified land covers as natural (managed and unmanaged ‘forest’ classes, ‘shrubland’, ‘grassland’, ‘barren land’, and ‘wetland’), agricultural (class ‘cropland’), urban, and water (Fig. 1). We resampled rasters to 300-m resolution to reduce spatial mismatch between species and environmental data50, and rescaled continuous variables (− 1 to 1) to facilitate effects comparisons.
We fit the SSF using a conditional Poisson regression model, which yields equivalent estimates to the conditional logistic regression model typically used for SSFs51. We included random slopes for the continuous variables51 to account for individual variation among wolves. We fit the model using the glmmTMB package52 in program R. We used variance inflation factors (VIF) and pairwise correlations to test for multicollinearity of variables with thresholds of 10 and 0.70, respectively53. We selected from two candidate models based on the lowest AIC, or the competing model (ΔAIC < 2) with fewer terms44: one model contained road density and proportion of natural land cover, and another model contained these variables interacting with movement type. We created used-habitat calibration plots (UHC) to visualize how well model predictions characterize used locations by plotting the distribution of an explanatory variable at used locations and overlaying this with the distribution of explanatory variable values predicted by the model54. As a measure of ecological importance of statistical estimates, we spatially predicted relative strength of selection (RSS;55) throughout the study area by calculating RSS as the probability of selecting a given point over a point with average variable values in our study area, scaling probabilities from 0 to 1.
Modeling connectivity
We used Circuitscape software to assess landscape-level connectivity without the assumption of animals having landscape knowledge56. We inverted the RSS surface raster to obtain an estimate of movement resistance57 then replaced each cell from the movement resistance surface with nodes connected by resistors, translating connectivity to ‘current flow’. We limited connectivity analysis to non-resident movements if the results of our SSF indicated significant differences (α < 0.05) in habitat selection for resident and non-resident wolf movements. We incorporated part of Indiana (8795 km2) into the study area to avoid a spatial interruption that would bias the circuit connectivity model. We assigned 154 points at about 40-km intervals along the perimeter of the study area and calculated connectivity between all pairs of points, providing an omnidirectional connectivity map for animals moving randomly through the landscape56. Connectivity between perimeter points is prone to edge effects as connectivity increases near these points. We therefore placed perimeter points at the midpoint of a 15-km buffer bordering the study area edge, filled cells within this buffer with the average movement resistance value of the study area, and removed this buffer after analysis57. To assess how the connectivity map compared to a previous species distribution model (SDM) and circuit connectivity analysis for gray wolves in the eastern USA and southern Canada24, we resampled our RSS and circuit connectivity maps to 1-km resolution and calculated Pearson’s correlation coefficients between the RSS and SDM rasters and their respective circuit connectivity rasters.
Results
We retained 24,540 steps, with a median of 878 steps per wolf (range = 156–3811 steps). Of these steps, 16,668 were classified as resident, 1656 as dispersing, and 6216 as floating movements, with median step-lengths of 609, 1011, and 655 m for resident, dispersing, and floating movements, respectively. Resident, dispersing, and floating movements were found for 72, 20, and 24 wolves, respectively, and 17 of 96 wolves displayed multiple movement types. We used a maximum annual range size of 561 km2 for resident wolves to support movement type classification using the NSD-method and visual inspection, though average estimated annual range sizes were smaller for wolves classified as resident (mean = 195 km2, StDev = 116) than for dispersers (9294 km2, StDev = 15,065) or floaters (8608 km2, StDev = 11,119). Short-distance dispersal events occurred (n = 16, median = 54 km), with the longest dispersal being 615 km. Proportions of natural (VIF = 3.36) and agricultural (VIF = 3.03) cover were correlated (r = − 0.80), and we retained proportion of natural cover as it is inverse to the combined proportions of agricultural and urban cover, thus a stronger proxy of human disturbance.
The model retaining movement type interacting with proportion of natural land cover and road density indicated no habitat selection differences among resident, dispersing, and floating wolves (Table 1; Fig. 2). Our final model included road density and proportion of natural land cover without interactions with movement type, and had a lower AIC value than the model including these interactions (ΔAIC = 5.2). This final model suggested wolves avoided areas with greater road density and selected for areas with greater proportions of natural cover (Fig. 3).

Characteristics of used and available steps for road density (km/km2, top panel) and proportion of natural cover (0–100, bottom panel) for 96 Gy wolves (Canis lupus) in the western Great Lakes region, USA, and southern Ontario and Manitoba, Canada, 2017–2021. Circles represent average selection of wolves by movement type, and error bars are 95% confidence intervals of these averages.

Predictions from a step-selection function for gray wolves (Canis lupus) in the western Great Lakes region, USA, and southern Ontario and Manitoba, Canada, 2017–2021. Predicted values are probabilities of selection relative to the average variable value of used and available steps (dashed lines, average proportion of natural land cover = 90.42%, average road density = 0.83 km/km2).
Calibration of our top-ranked model was successful for road density and reasonable for proportions of natural cover, based on visual inspection of overlap between predicted and used values in UHC plots (Appendix A).
Areas previously estimated suitable that were not selected by wolves included parts of the Lower Peninsula of Michigan, and isolated and fragmented areas east of the Missouri river in North and South Dakota. Non-resident movements in our dataset were limited to established wolf range. Because we detected no differences in habitat selection between resident and non-resident movements, we retained all data for connectivity analysis. Our circuit connectivity map indicated highest connectivity for wolves in the northern and eastern parts of Ontario and was positively correlated (r = 0.75) with the circuit connectivity raster of the SDM (Fig. 1). The resampled RSS raster (Appendix B) was positively correlated (r = 0.78) with a previous habitat suitability raster24.
Discussion
Our estimate of gray wolf habitat selection in the western Great Lakes region of the USA and southern Ontario and Manitoba, Canada, supported our prediction that wolves avoid areas with high road densities and select for areas with high proportions of natural cover. Contrary to our prediction, we found no differences among habitat selection of wolves exhibiting resident, dispersing, or floating movements. We also confirmed unoccupied habitat in the western Great Lakes region is limited, and connectivity between occupied and unoccupied habitat is constrained by the Great Lakes, areas of extensive agriculture, and urban areas.
Habitat selection of resident and non-resident wolves in our study was similar, though previous studies found it can differ by disturbance type. Gray wolves in Portugal displayed increased tolerance toward roads and settlements during dispersal, but not towards areas with higher livestock densities or windfarms11. The absence of increased tolerance for high road densities or proportions of urban or agricultural land cover during non-residency may be due to non-resident movements in our study occurring within established wolf range, causing high similarity or overlap in habitat characteristics to which residents and non-residents are exposed. Only four dispersing wolves exceeded estimated mean dispersal distances (range = 29–148 km) for the western Great Lakes region58. The prevalence of short-distance dispersals (median = 54 km), and dispersal and floating movements being largely limited to established wolf range, may be caused by increased opportunities for non-residents to join existing packs due to high pack densities59,60. Alternatively, with little unoccupied habitat available, non-residents may traverse conspecific territories when higher prey availability or lower human disturbance in occupied habitat outweighs risks of encountering conspecifics17.
The relative selection strength (RSS) map was positively correlated with a habitat suitability map for the same area based on wolf winter track surveys24, while derived connectivity maps were also positively correlated. This similarity provides validation of current wolf range and habitat connectivity throughout the western Great Lakes region from independent data. Our results also suggest that accounting for differences between resident and non-resident movements using data transformations61, or by limiting connectivity analyses to non-resident movement data15,16, may be unnecessary when non-resident movements are largely limited to established range or otherwise similar to resident individuals.
We note several limitations to our study. Similar to how movement resistance rasters are calculated from habitat suitability maps24, we calculated a resistance raster by inverting the RSS map resulting from our SSF62. This is not ideal because RSS values are conditional probabilities55, though no alternative approach is available. Additionally, we used a traditional step-selection function that builds a habitat selection analysis upon estimation of movement, which can introduce a bias in habitat selection because habitat selection to an extent may depend on the movement capability of animals63. A fully mechanistic approach to classifying movement types is unavailable, and the migrateR package was developed primarily for migratory animals, so classification depends in part on visual interpretation of data42. Also, wolves can swim up to 2 km64 but can cross larger waterbodies during freeze-over65. Our approach resulted in waterbodies having above average resistance to movement due to low natural, terrestrial cover, but as the Great Lakes are roadless they have a lower resistance than areas with low natural cover and high road density. Year-round estimates of connectivity are imperfect due to seasonal changes in movement resistance of water. Using GPS locations collected at shorter intervals could be used to assess finer-scale wolf movements, and may reveal differences in habitat selection among movement types11. Analysis including non-resident movements beyond established range also is needed to confirm whether differences in habitat use between resident and non-resident wolves depend on differences in the range of conditions they occur in. Finally, wolves generally avoid higher road densities but can select for minor, lower traffic roads for efficient travel66. The road databases used here generally group unpaved rural roads, that wolves are known to use, with roads in suburban areas that wolves would likely avoid, thus testing the response to road densities classified by road type was not possible.
We suggest potential for further recolonization of the western Great Lakes region is low, as unoccupied habitat and habitat connectivity are limited. The Straits of Mackinac can connect current range in the Upper Peninsula of Michigan with the Lower Peninsula during freeze-over, though recent crossings of the straits have been too infrequent for population establishment67. Recolonization of potential habitat in North and South Dakota is limited by low dispersal frequencies and high anthropogenic mortality68, and connectivity with current range may be higher through Manitoba than through Minnesota.
Conclusions
We offer further support that gray wolves in the western Great Lakes region select for areas with high proportions of natural cover, and against human disturbance as indexed by road densities. We found no differences in habitat selection among wolves that were resident, dispersing, or floating. The need to limit connectivity analyses to non-resident movements, or to apply transformations to data of primarily resident wolf movements, will depend on the magnitude of differences in habitat characteristics experienced by resident and non-resident individuals. As most wolf habitat in the western Great Lakes region appears occupied and there is limited habitat connectivity between currently occupied range and limited unoccupied range in the USA part of the western Great Lakes region, further recolonization appears most likely through Canada to connect with wolf habitat in North Dakota, and across the Straits of Mackinaw to connect with habitat in the Lower Peninsula of Michigan. Interjurisdictional cooperation will be important to improve landscape connectivity for gray wolves between Canada and the USA. If recolonization of areas beyond current wolf range in the Great Lakes region is desired, promoting human-wolf co-existence in areas most likely to be recolonized is pertinent, though further natural recolonization within and beyond the Great Lakes region appears limited by the dominance of urban and agricultural areas surrounding current range.
Data availability
Data supporting the conclusions of this article are not publicly available as the subject species (gray wolf) is federally protected under the Endangered Species Act, and subject to poaching within the study area. Data are available upon request from co-authors affiliated with the Department of Natural Resources of Minnesota (john.erb@state.mn.us), Wisconsin (david.macfarland@wisconsin.gov), and Michigan (petroeljet@michigan.gov).
References
Rodríguez, J. P., Brotons, L., Bustamante, J. & Seoane, J. The application of predictive modelling of species distribution to biodiversity conservation. Divers. Distrib. 13, 243–251. https://doi.org/10.1111/j.1472-4642.2007.00356.x (2007).
Guisan, A. et al. Predicting species distributions for conservation decisions. Ecol. Lett. 16, 1424–1435. https://doi.org/10.1111/ele.12189 (2013).
Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 34, 487–515. https://doi.org/10.1146/132419 (2003).
Correa Ayram, C. A., Mendoza, M. E., Etter, A. & Salicrup, D. R. P. Habitat connectivity in biodiversity conservation: A review of recent studies and applications. Prog. Phys. Geogr. 40, 7–37. https://doi.org/10.1177/0309133315598713 (2016).
McLoughlin, P. D., Morris, D. W., Fortin, D., Vander Wal, E. & Contasti, A. L. Considering ecological dynamics in resource selection functions. J. Anim. Ecol. 79, 4–12. https://doi.org/10.1111/j.1365-2656.2009.01613.x (2010).
Benz, R. A. et al. Dispersal ecology informs design of large-scale wildlife corridors. PLoS One 11, e0162989. https://doi.org/10.1371/journal.pone.0162989 (2016).
Chetkiewicz, C. B., St-Clair, C. C. & Boyce, M. S. Corridors for conservation: Integrating pattern and process. Annu. Rev. Ecol. Evol. Syst. 37, 317–342. https://doi.org/10.1146/annurev.ecolsys.37.091305.110050 (2006).
Laundré, J. W., Hernandez, L. & Ripple, W. J. The landscape of fear: Ecological implications of being afraid. Open Ecol. J. 3, 1–7. https://doi.org/10.2174/1874213001003030001 (2010).
Oriol-Cotterill, A., Valeix, M., Frank, L. G., Riginos, C. & Macdonald, D. W. Landscapes of coexistence for terrestrial carnivores: The ecological consequences of being downgraded from ultimate to penultimate predator by humans. Oikos 124, 1263–1273. https://doi.org/10.1111/oik.02224 (2015).
Smith, J. B., Nielsen, C. K. & Hellgren, E. C. Suitable habitat for recolonizing large carnivores in the midwestern USA. Oryx 50, 555–564. https://doi.org/10.1017/S0030605314001227 (2016).
Rio-maior, H., Nakamura, M., Álvares, F. & Beja, P. Designing the landscape of coexistence: Integrating risk avoidance, habitat selection and functional connectivity to inform large carnivore conservation. Biol. Conserv. 235, 178–188. https://doi.org/10.1016/j.biocon.2019.04.021 (2019).
Vasudev, D., Fletcher, R. J., Goswami, V. R. & Krishnadas, M. From dispersal constraints to landscape connectivity: Lessons from species distribution modeling. Ecography 38, 967–978. https://doi.org/10.1111/ecog.01306 (2015).
Abrahms, B. et al. Does wildlife resource selection accurately inform corridor conservation?. J. Appl. Ecol. 54, 412–422. https://doi.org/10.1111/1365-2664.12714 (2017).
Hinton, J. W. et al. Space use and habitat selection by resident and transient red wolves (Canis rufus). PLoS One 11, e0167603. https://doi.org/10.1371/journal.pone.0167603 (2016).
Elliot, N. B., Cushman, S. A., Macdonald, D. W. & Loveridge, A. J. The devil is in the dispersers: Predictions of landscape connectivity change with demography. J. Appl. Ecol. 51, 1169–1178. https://doi.org/10.1111/1365-2664.12282 (2014).
Dondina, O., Meriggi, A., Bani, L. & Orioli, V. Decoupling residents and dispersers from detection data improve habitat selection modelling: The case study of the wolf in a natural corridor. Ethol. Ecol. Evol. 34(6), 617–635. https://doi.org/10.1080/03949370.2021.1988724 (2022).
Morales-González, A., Fernández-Gil, A., Quevedo, M. & Revilla, E. Patterns and determinants of dispersal in grey wolves (Canis lupus). Biol. Rev. 97, 466–480. https://doi.org/10.1111/brv.12807 (2022).
Gastón, A. et al. Response to agriculture by a woodland species depends on cover type and behavioural state: Insights from resident and dispersing Iberian lynx. J. Appl. Ecol. 53, 814–824. https://doi.org/10.1111/1365-2664.12629 (2016).
Thorsen, N. H. et al. Movement and habitat selection of a large carnivore in response to human infrastructure differs by life stage. Mov. Ecol. 10, 1–14. https://doi.org/10.1186/s40462-022-00349-y (2022).
Boitani, L. Wolf conservation and recovery. In (eds. Mech, M. & Boitani, L.) Wolves: Behavior, Ecology, And Conservation 317–341 (University of Chicago Press, 2003). https://doi.org/10.14430/arctic540
Wydeven, A. P. et al. History, population growth, and management of wolves in Wisconsin. In Recovery of Gray Wolves in the Great Lakes region of the United States (eds Wydeven, A. P. et al.) 87–105 (Springer, 2009). https://doi.org/10.1007/978-0-387-85952-1_6.
Beyer, D. E., Peterson, R. O., Vucetich, J. A. & Hammill, J. H. Wolf population changes in Michigan. In Recovery of Gray Wolves in the Great Lakes region of the United States (eds Wydeven, A. P. et al.) 65–85 (Springer, 2009). https://doi.org/10.1007/978-0-387-85952-1_5.
USFWS (U.S. Fish and Wildlife Service). Gray wolf biological report: Information on the species in the lower 48 United States. US Government Printing Office, Washington, DC (2020).
van den Bosch, M. et al. Identifying potential gray wolf habitat and connectivity in the eastern USA. Biol. Conserv. 273, 109708. https://doi.org/10.1016/j.biocon.2022.109708 (2022).
Mech, L. D. & Boitani, L. 2003. Wolf social ecology. In (eds. Mech, M. & Boitani, L.) Wolves: Behavior, Ecology, and Conservation 317–341 (University of Chicago Press, 2003). https://doi.org/10.14430/arctic540
Mancinelli, S., Falco, M., Boitani, L. & Ciucci, P. Social, behavioural and temporal components of wolf (Canis lupus) responses to anthropogenic landscape features in the central Apennines, Italy. J. Zool. 309, 114–124. https://doi.org/10.1111/jzo.12708 (2019).
Messier, F. Solitary living and extraterritorial movements of wolves in relation to social status and prey abundance. Can. J. Zool. 63(2), 239–245. https://doi.org/10.1139/z85-037 (1985).
Mech, L. D. Unexplained patterns of grey wolf Canis lupus natal dispersal. Mamm. Rev. 50, 314–323. https://doi.org/10.1111/mam.12198 (2020).
Imbert, C. et al. Why do wolves eat livestock. Factors influencing wolf diet in northern Italy. Biol. Conserv. 195, 156–168. https://doi.org/10.1016/j.biocon.2016.01.003 (2016).
Hill, J. E., DeVault, T. L. & Belant, J. L. A review of ecological factors promoting road use by mammals. Mamm. Rev. 51, 214–227. https://doi.org/10.1111/mam.12222 (2021).
USFWS (U.S. Fish and Wildlife Service). Post-delisting monitoring plan for the western Great Lakes distinct population segment of the gray wolf. U.S. Fish and Wildlife Service, Twin Cities Field Office and Midwest Region, Bloomington, MN and Ft. Snelling, MN (2008).
Ouellette, M., Remmel, T. K. & Perera, A. H. A spatial database of historical wildfire and timber harvesting in the boreal Area of the Undertaking of Ontario: The methodological framework. Ontario MNRF, TR 37 (2020).
Beck, H. E. et al. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 5, 180214. https://doi.org/10.1038/sdata.2018.214 (2018).
Scott, R. W. & Huff, F. A. Impacts of the Great Lakes on regional climate conditions. J. Great Lakes Res. 22, 845–863. https://doi.org/10.1016/S0380-1330(96)71006-7 (1996).
United States Geological Survey. USGS EROS archive: Digital elevation global 30 arc-second elevation (GTOPO30) (1996). https://doi.org/10.5066/F7DF6PQS.
Homer, C. et al. Developing a new North American land cover product at 30m resolution: Methods, results and future plans. In AGU Fall Meeting Abstracts Vol. 2017 GC52C-01 (2017).
DelGiudice, G. D., McCaffery, K. R., Beyer Jr, D. E. & Nelson, M. E. Prey of wolves in the Great Lakes region. In (eds. Wydeven, A. P. et al.) Recovery of Gray Wolves in the Great Lakes Region of the United States 155–204 (Springer, 2009). https://doi.org/10.1007/978-0-387-85952-1_10.
Simpson, T. L., Thiel, R. P., Sailer, D. T., Reineke, D. M. & Thomsen, M. Demographics of Gray Wolf (Canis lupus) packs recolonizing variable habitats in central Wisconsin. Northeast. Nat. 30, 75–98. https://doi.org/10.1656/045.030.0108 (2023).
Potvin, M. J. et al. Monitoring and habitat analysis for wolves in upper Michigan. J. Wildl. Manag. 69, 1660–1669. https://doi.org/10.2193/0022-541X(2005)69[1660:MAHAFW]2.0.CO;2 (2005).
Erb, J. D. & Humpal, C. Minnesota wolf population update 2019. Minnesota Department of Natural Resources, St. Paul (2020).
Powell, R. A. & Proulx, G. Trapping and marking terrestrial mammals for research: Integrating ethics, performance criteria, techniques, and common sense. ILAR J. 44, 259–276. https://doi.org/10.1093/ilar.44.4.259 (2003).
Bunnefeld, N. et al. A model-driven approach to quantify migration patterns: Individual, regional and yearly differences. J. Anim. Ecol. 80, 466–476. https://doi.org/10.1111/j.1365-2656.2010.01776.x (2011).
Spitz, D. B., Hebblewhite, M. & Stephenson, T. R. ‘MigrateR’: Extending model-driven methods for classifying and quantifying animal movement behavior. Ecography 40, 788–799. https://doi.org/10.1111/ecog.02587 (2017).
R Core Team. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria (2021). https://www.R-project.org/.
Burnham, K. P. & Anderson, D. R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach (Springer, 2004). https://doi.org/10.1007/b97636.
Calenge, C. Home range estimation in R: The adehabitatHR package. Office national de la classe et de la faune sauvage: Saint Benoist, Auffargis, France (2011).
Thurfjell, H., Ciuti, S. & Boyce, M. S. Applications of step-selection functions in ecology and conservation. Mov. Ecol. 2, 4. https://doi.org/10.1186/2051-3933-2-4 (2014).
United States Census Bureau. TIGER/Line shapefiles 2020: Roads (2020). https://www.census.gov/geographies/mapping-files/time-series/geo/tiger-line-file.html.
Statistics Canada. Road Network File, Catalogue no. 92-500-X (2020). https://www150.statcan.gc.ca/n1/en/catalogue/92-500-X.
Guisan, A. & Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 8, 993–1009. https://doi.org/10.1111/j.1461-0248.2005.00792.x (2005).
Muff, S., Signer, J. & Fieberg, J. Accounting for individual-specific variation in habitat-selection studies: Efficient estimation of mixed-effects models using Bayesian or frequentist computation. J. Anim. Ecol. 89, 80–92. https://doi.org/10.1111/1365-2656.13087 (2020).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. https://doi.org/10.3929/ethz-b-000240890 (2017).
Dormann, C. F. et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. https://doi.org/10.1111/j.1600-0587.2012.07348.x (2013).
Fieberg, J. R. et al. Used-habitat calibration plots: A new procedure for validating species distribution, resource selection, and step-selection models. Ecography 41, 737–752. https://doi.org/10.1111/ecog.03123 (2018).
Avgar, T., Lele, S. R., Keim, J. L. & Boyce, M. S. Relative selection strength: Quantifying effect size in habitat-and step-selection inference. Ecol. Evol. 7, 5322–5330. https://doi.org/10.1002/ece3.3122 (2017).
McRae, B. H. & Shah, V. B. Circuitscape user's guide. The University of California, Santa Barbara, CA (2009). http://www.circuitscape.org.
Buchholtz, E. K., Stronza, A., Songhurst, A., McCulloch, G. & Fitzgerald, L. A. Using landscape connectivity to predict human-wildlife conflict. Biol. Conserv. 248, 108677. https://doi.org/10.1016/j.biocon.2020.108677 (2020).
Treves, A., Martin, K. A., Wiedenhoeft, J. E. & Wydeven, A. P. Dispersal of gray wolves in the Great Lakes region. In (eds. Wydeven, A. P. et al.) Recovery of Gray Wolves in The Great Lakes Region of the United States 191–204 (Springer, 2009). https://doi.org/10.1007/978-0-387-85952-1_12.
Brainerd, S. M. et al. The effects of breeder loss on wolves. J. Wildl. Manag. 72, 89–98. https://doi.org/10.2193/2006-305 (2008).
Jimenez, M. D. et al. Wolf dispersal in the Rocky Mountains, Western United States: 1993–2008. J. Wildl. Manag. 81, 581–592. https://doi.org/10.1002/jwmg.21238 (2017).
Keeley, A. T., Beier, P. & Gagnon, J. W. Estimating landscape resistance from habitat suitability: Effects of data source and nonlinearities. Landsc. Ecol. 31, 2151–2162. https://doi.org/10.1007/s10980-016-0387-5 (2016).
Osipova, L. et al. Using step-selection functions to model landscape connectivity for African elephants: Accounting for variability across individuals and seasons. Anim. Conserv. 22, 35–48. https://doi.org/10.1111/acv.12432 (2019).
Avgar, T., Potts, J. R., Lewis, M. A. & Boyce, M. S. Integrated step selection analysis: Bridging the gap between resource selection and animal movement. Methods Ecol. Evol. 7, 619–630. https://doi.org/10.1111/2041-210X.12528 (2016).
Darimont, C. T. & Paquet, P. C. Gray wolves, Canis lupus, of British Columbia’s Central and North Coast: Distribution and conservation assessment. Can. Field-Nat. 116, 416–422 (2002).
Orning, E. K. et al. Emigration and first-year movements of initial Wolf translocations to Isle Royale. Northeast. Nat. 27, 701–708. https://doi.org/10.1656/045.027.0410 (2020).
Zimmermann, B., Nelson, L., Wabakken, P., Sand, H. & Liberg, O. Behavioral responses of wolves to roads: Scale-dependent ambivalence. Behav. Ecol. 25(6), 1353–1364. https://doi.org/10.1093/beheco/aru134 (2014).
Stricker, H. K., Gehring, T. M., Donner, D. & Petroelje, T. Multi-scale habitat selection model assessing potential gray wolf den habitat and dispersal corridors in Michigan, USA. Ecol. Modell. 397, 84–94. https://doi.org/10.1016/j.ecolmodel.2018.12.021 (2019).
Licht, D. S. & Fritts, S. H. Gray wolf (Canis lupus) occurrences in the Dakotas. Am. Midl. Nat. 132, 74–81. https://doi.org/10.2307/2426202 (1994).
Acknowledgements
We thank individuals from the respective government agencies and the public that assisted with data collection. We thank the U.S. Geological Survey and L.D. Mech for data contributions to this research. Primary funding for manuscript preparation was provided by the U.S. Fish and Wildlife Service through the Great Lakes Fish and Wildlife Restoration Act and Boone and Crockett Program at Michigan State University.
Funding
Funding was provided by the U.S. Fish and Wildlife Service through the Great Lakes Fish and Wildlife Restoration Act and Boone and Crockett Program at Michigan State University.
Author information
Authors and Affiliations
Contributions
M.V.D.B. was responsible for formal data analysis, visualization and original draft writing. M.V.D.B., K.F.K., M.G.G., B.R.P. and J.L.B. developed the methodology. D.E.B., J.D.E., D.M.M., D.C.N., J.L.P.T., and J.L.B. were responsible for funding acquisition, and J.L.B. was responsible for project administration and supervision. All co-authors were involved with conceptualization of the research, data curation as well as review and editing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van den Bosch, M., Kellner, K.F., Gantchoff, M.G. et al. Habitat selection of resident and non-resident gray wolves: implications for habitat connectivity. Sci Rep 13, 20415 (2023). https://doi.org/10.1038/s41598-023-47815-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-47815-0
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
