
Abstract
Diabetes is a common metabolic disorder that has become a major health problem worldwide. In this study, we investigated the role of rutin in attenuating diabetes and preventing diabetes-related colon lesions in mice potentially through regulation of gut microbiota. The rutin from tartary buckwheat as analyzed by HPLC was administered intragastrically to diabetic mice, and then the biochemical parameters, overall community structure and composition of gut microbiota in diabetic mice were assayed. The results showed that rutin lowered serum glucose and improved serum total cholesterol, low-density lipoprotein, high-density lipoprotein, triglyceride concentrations, tumor necrosis factor-α, interleukin-6, and serum insulin in diabetic mice. Notably, rutin obviously alleviated colon lesions in diabetic mice. Moreover, rutin also significantly regulated gut microbiota dysbiosis and enriched beneficial microbiota, such as Akkermansia (p < 0.05). Rutin selectively increased short-chain fatty acid producing bacteria, such as Alistipes (p < 0.05) and Roseburia (p < 0.05), and decreased the abundance of diabetes-related gut microbiota, such as Escherichia (p < 0.05) and Mucispirillum (p < 0.05). Our data suggested that rutin exerted an antidiabetic effect and alleviated colon lesions in diabetic mice possibly by regulating gut microbiota dysbiosis, which might be a potential mechanism through which rutin alleviates diabetes-related symptoms.
Similar content being viewed by others

Introduction
Diabetes, classified as type 1 diabetes mellitus (T1DM) and type 2 diabetes mellitus (T2DM), is a metabolic disorder accompanied by insufficient insulin production or insulin resistance1. To date, although there have been some antidiabetic drugs in clinical practice, there is still a general lack of effective approaches to prevent the initiation and development of diabetes and diabetes-related diseases2. In T2DM patients, the incidence of colorectal cancer (CRC) is higher than that in nondiabetic populations3. Hyperinsulinemia, hyperglycemia and chronic inflammation are common risk factors for CRC and liver cancer4. Colon lesions appear in an initial phase of diabetes-induced CRC. Due to the lack of understanding of the CRC development process, there are currently no effective methods to prevent the development of colon lesions. Studies have shown that the incidence of diabetes is closely related to intestinal flora which plays an important role in host metabolism, immune regulation, the development of inflammation-derived disease and chronic inflammation5,6,7. The composition of the gut microbiota is also significantly correlated with obesity and diabetes8,9,10. Although an increasing number of drugs are being used to treat diabetes and related diseases, the side effects of many drugs cannot be ignored11. Therefore, it is of great value to develop new effective and nontoxic drugs for the treatment of diabetes and its related diseases. At present, increasing attention has been paid to the development of natural products due to their low cost and minimal side effects in diabetes treatment.
Rutin, a type of flavonoid glycoside, is mainly found in plants, such as apple, black tea, vegetables and buckwheat12,13. It was reported that rutin had antioxidant activity and promoted intestinal absorption14. Rutin has attracted extensive attention due to its various biological activities, such as antioxidant15, antiinflammatory16,17, antidiabetic13,18,19,20,21,22, and anticancer activities23,24,25. Recent studies have indicated that the regulation of gut microbiota might be one of the potential mechanisms by which natural products achieve antidiabetic effects26. However, whether rutin from tartary buckwheat could improve diabetes and colon lesions by modulating gut microbiota is still not clear. Here, we investigated the effect of rutin on reducing the diabetes pathological state, diabetes-related colon lesions, overall community structure and composition of gut microbiota in diabetic mice and their potential relationship. Our discovery of rutin reducing colon lesions and modulating the gut microbiota composition in diabetic mice provided evidence for the therapeutic potential of rutin to treat diabetes-related symptoms possibly by promoting intestinal health.
Materials and methods
Chemicals and reagents
Control diet (10% lipids, 19% protein, 71% carbohydrates, Cat. AIN-93) was obtained from Jiangsu-Xietong, Inc, Nanjing, China. A high-fat diet (HFD) (60% fat, Cat. D12492) was from Research Diets Incorporated Company, New Brunswick, New Jersey, USA. Streptozotocin (STZ, Cat. 18883-66-4) and saline (Cat. 231-598-3) were purchased from Sigma-Aldrich Co., Ltd (St. Louis, MO, USA). Tartary buckwheat powder (Zhongku No. 3) was obtained from Weining Dongfang Shengu Co., Ltd. (Guizhou, China). The AB-8 macroporous resins (Cat. M0042) were purchased from Solarbio Technology Ltd (Beijing, China). All other chemical reagents used in this study were of analytical grade.
Rutin preparation from tartary buckwheat
The rutin used in this study was obtained by ethanol extraction from tartary buckwheat powder, the purity of rutin prepared by us reached more than 96% and stored in our laboratory as previously described27.
Establishment and treatment of the diabetic mouse model
Forty male C57BL/6 J mice (8 weeks old) were obtained from Shanghai Slac Laboratory Animal Co., Ltd (Shanghai, China). All mice were raised in separate cages under conditions in which they could freely ingest available solid feed and water. The mice were caged at a room temperature of 23 ± 1 °C, humidity of 60% and a 12 h light–dark cycle. The mice were provided with ad libitum access to food and water for 1 week. The STZ-induced T1DM mouse model and HFD/STZ-induced T2DM mouse model were established as previously described28,29. Briefly, the mice received intraperitoneal injections of STZ (45 mg/kg/day) for five consecutive days. Four weeks after diabetes induction, fasting blood glucose level of 16.7 mM and above were considered diabetic. The HFD/STZ-induced T2DM mouse model were established, the ND group was fed with a normal diet (10% lipids, 19% protein, 71% carbohydrates) sustainably, the other three groups of mice were fed the HFD (45% lipids, 19% proteins and 36% carbohydrates, research diets incorporated company, New Brunswick, New Jersey, USA) for 8 weeks. After 8 weeks of HFD feeding, the mice were fasted for 12 h overnight. The HFD groups were received intraperitoneal injections of STZ (35 mg·kg − 1, dissolved at 0.1 mol/L cold citrate buffer, pH 4.4) daily for three consecutive days. The fasting serum glucose was sampled from the tail vein and determined by an enzymatic colorimetric assay using a modified glucose oxidase–peroxidase method (Roche Diagnostics, Mannheim, Germany) and a glucose analyzer (Roche-Hitachi 917). Diabetes was defined as abdominal serum glucose ≥ 16.7 mmol/L for two consecutive days30,31,32. In a follow-up experiment, the successfully established diabetes mice were used in the experiment, and the failed models were excluded. The dosage of rutin was optimized according to previous reports33. Rutin or saline was administered intragastrically in T1DM and T2DM mice groups ( 5 mice per group) once a day for 4 weeks as follows: ND group (normal diet mice with daily administration of saline), T1DM group (T1DM mice with daily administration of saline), T1DM-Rutin100 group (mice with daily administration of 100 mg Rutin/kg body weight), and T1DM-Rutin200 group (mice with daily administration of 200 mg Rutin/kg body weight), T2DM control group (mice with daily administration of normal saline), T2DM-Rutin100 group (mice with daily administration of 200 mg Rutin/kg body weight), T2DM-Rutin200 group (mice with daily administration of 200 mg Rutin/kg body weight). Four weeks after rutin treatment, fresh fecal samples were collected under a sterile environment and frozen at − 80 °C for subsequent analysis. Serum samples were collected and centrifuged at 3000 rpm for 15 min at 4 °C. Colon tissues and intestinal contents were collected and frozen in liquid nitrogen and stored at − 80 °C. All animal experiments were approved by the Wenzhou University Animal Care and Use Committee (Wenzhou, China) with approval number WZU-2020-010. All methods were performed in accordance with the relevant guidelines and regulations. This study is reported in accordance with ARRIVE guidelines (Animal Research: Reporting of in vivo Experiments; https://arriveguidelines.org).
Biochemical analysis
Commercial detection kits (Jiancheng Bioengineering Institute, Nanjing, China) were used to measure total serum cholesterol (TC) (Cat. A111-1-1), triglyceride (TG) (Cat. A110-1-1), low-density lipoprotein cholesterol (LDL-C) (Cat. A113-1-1) and high-density lipoprotein cholesterol (HDL-C) (Cat. A112-1-1). Commercial detection kits (MultiSciences Biotech Co., Ltd., Hangzhou, China) were used to measure serum TNF-α (Cat. EK282/4-01) and IL-6 (Cat. 70-EK206/3-96) levels. The fasting serum insulin levels were measured using a Mouse INS(Insulin) ELISA Kit (Cat. D721159-0096, Sangon Biotech Co., Ltd., Shanghai, China).
Haematoxylin & Eosin (H&E) staining
Colon tissues were fixed with 4% paraformaldehyde for 48 h and all samples were dehydrated with ethanol and embedded in paraffin wax. Tissues were sliced into 5 μm-thick Sections (3–5 sections/specimen) and stained with H&E. Samples were sealed with neutral resin and observed under optical microscope (× 400 magnification) for evaluation. The histopathological score of colonic lesions was measured using the following parameters: the degree of inflammation, crypt damage, number of vacuoles and arrangement of villi. The final pathological scores were described as previously reported34.
Immunohistochemistry
For the immunohistochemistry assays, the colon tissues were dissected, fixed in 4% paraformaldehyde for 12 h at 4 °C, dehydrated overnight with 30% sucrose at 4 °C, embedded in optimal cutting temperature compound and immediately frozen at − 80 °C. Samples sectioned at a thickness of 10 μm were washed with PBS and permeabilization solution with 0.5% Triton X-100 in turn. Samples were then blocked in 3% horse serum in PBS for 30 min and incubated sequentially with the following primary antibodies: anti-collagen I (Cat. ab260043, Abcam Ltd., UK). After incubation with corresponding secondary antibodies (Cat. A11006, Invitrogen, Carlsbad, CA, USA) at room temperature for 1 h, sample sections were treated with a peroxidase substrate DAB kit (Cat. ab64238, Abcam Ltd., UK) and counterstained with hematoxylin.
16S rRNA gene sequencing
Total bacterial genomic DNA of fecal samples was extracted using a rapid DNA spin extraction kit (Cat. 117033600, MP Biomedicals, Santa Ana, CA, USA) in accordance with the manufacturer’s instructions. The DNA content was then detected by agarose gel electrophoresis and Nanodrop method (Thermo Scientific, NC2000). The DNA was amplified with specific bacterial primers targeting the 16S rRNA gene containing the V3-V4 region using universal primers 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’). PCR amplification was purified using Agencourt AMPure Beads (Beckman Coulter, Indianapolis, IN). The PCR amplification products were quantified using the fluorescence reagent Quant-iT PicoGreen dsDNA Assay Kit (Cat. P11496, Invitrogen, Carlsand, CA, USA) and a quantitative microplate reader (BioTek, FLx800). Sequencing was carried out using the Illumina MiSeq platform and MiSeq-pe250 in Shanghai Pesino Biotechnology Co., Ltd (Shanghai, China). All sample sequences were clustered according to the distance between the sequences using Uparse software (v7. 0. 1001), and were divided into operational classification units (OTUs) by plotting the relationship graph between the changes in OTUs and the clustering similarity value. A 97% similarity value was selected for OTU analysis and taxonomic analysis. QIIME 2.0 was then applied to calculate the α-diversity index, including the Chao1 index, Shannon index and Simpson index. UniFrac distance measurement and principal coordinate analysis (PCoA) were selected for β-diversity analysis of the gut microbiota structure, and the gplots package from R software (R 3.4.0) was used for heatmap analysis. Linear discriminant analysis (LDA) of the effect size (LEfSe) was performed with LEfSe software (Version 1.0) and applied to calculate the OTU abundance to determine the differences between groups. Heatmap analysis was performed using R Statistical Software (R version 3.4.0: Foundation for Statistical Computing, Vienna, Austria).
Statistical analysis
First, the normality of the distribution of dates was tested using the Shapiro–Wilk normality test. Second, statistical comparisons of different groups were evaluated by unpaired Student's t test or one-way ANOVA followed by Tukey’s multiple comparisons. If the data were nonnormally distributed, the Kruskal–Wallis test followed by the Mann–Whitney U test was used. Spearman’s correlation analysis was performed using R Statistical Software (R version 3.4.0: Foundation for Statistical Computing, Vienna, Austria)35, and a clustering heatmap of correlation coefficients was calculated by Ward's hierarchy. GraphPad Prism 6.00 (GraphPad Prism Software, San Diego, California, USA, www.graphpad.com) software was used for statis-tical analysis. All values were expressed as the means ± standard deviation (SD). A value of p < 0.05 was considered statistically significant.
Results
Rutin improved glucose and lipids metabolism in diabetic mice
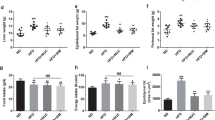
To investigate the effect of rutin on glucose and lipids in diabetic mice, the STZ-induced T1DM mouse model and HFD/STZ-induced T2DM mouse model were successfully established, followed by administration of 100 or 200 mg rutin per kg body weight for 4 weeks. The metabolic performances of diabetic mice showed that T1DM mice featured decreased body weights and HDL-C levels, increased fasting serum glucose, increased concentrations of TC, TG, and LDL-C, increased secretion of proinflammatory cytokines TNF-α and IL-6, and less secretion of fasting insulin (Fig. 1, Supplementary Table 1) compared with normal mice. Similar metabolic performances occurred in T2DM mice except for fasting serum insulin, which was upregulated in T2DM mice (Fig. 2, Supplementary Table 2). However, rutin alleviated the symptoms that are typical disorders related to glucose and lipid metabolism in T1DM and T2DM mice. Collectively, these observations suggest that rutin could effectively improve glucose and lipids metabolism in diabetic mice.

Effects of rutin on serum glucose and lipids in STZ-induced diabetic mice. (A) Fasting serum glucose. (B–I) Serum TC, TG, LDL-C, HDL-C, TNF-α, IL-6, and insulin levels. Values are means ± SD (n = 5): *p < 0.05, **p < 0.01, ***p < 0.001.

Effects of rutin on serum glucose and lipids in HFD/STZ-induced diabetic mice. (A) Fasting serum glucose. (B–I) Serum TC, TG, LDL-C, HDL-C, TNF-α, IL-6, and insulin levels. Values are means ± SD (n = 5): *p < 0.05, **p < 0.01, ***p < 0.001.
Rutin alleviated colon lesions in diabetic mice
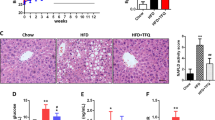
To investigate the effect of rutin on colonic lesions in diabetic mice, colon tissues were stained with H&E. Figure 3 and Supplementary Table 3 show that the villi of colon tissues in the T1DM and T2DM groups were arranged in a disorderly manner and were easy to break, along with many vacuolar changes and damaged crypts, compared to those in the ND group. Treatment with rutin exhibited significant protection against diabetes-related colon lesions. In addition, the collagen I protein level was increased in diabetic mice, which was reversed by rutin treatment. These results indicated that rutin could ameliorate the progression of intestinal fibrosis diseases in diabetic mice. Furthermore, the greater the amount of rutin used, the better the colon lesions were improved.

Rutin improved colon lesions in diabetic mice. (A) H&E staining of colon tissues in ND, T1DM, T1DM-Rutin100 and T1DM-Rutin200 mouse groups (scale bar: 20 μm). Red arrows: crypts; black arrows: villi. (B) Histopathological scores of colon tissues in T1DM mice using H&E staining. (C) H&E staining of colon tissues in ND, T2DM, T2DM-Rutin100 and T2DM-Rutin200 groups. Red arrows: vacuole; black arrows: villi (scale bar: 20 μm). (D) Histopathological scores of colon tissues in T2DM mice by H&E staining. (E) Immunohistochemistry indicated that collagen I protein levels were increased in diabetic mice, which was recovered by rutin treatment (scale bar: 20 μm). Values are means ± SD (n = 5): *p < 0.05, **p < 0.01, ***p < 0.001.
Rutin changed the overall community structure of the gut microbiota in diabetic mice
The V3-V4 region of bacterial 16S rRNA in fecal samples from the experimental mice was classified and sequenced to explore the effects of rutin on the overall community structure of gut microbiota in diabetic mice. For the experiment with T1DM mice, a total of 2,672,620 original sequences were obtained and an average of 127,267 ± 17,666 sequences were acquired for each sample. After quality screening, 2,119,396 sequences were generated with an average of 100,923 ± 13,879 sequences for each fecal sample. The abundance grade curves and species accumulation curves indicated that most of the gut microbial diversity was captured with the current sequencing depth in each sample (Fig. S1). Based on the α-diversity analysis, the results showed that the Chao1, Shannon and Simpson indexes reflected both the abundance and diversity of microbial communities. The Chao1 index was significantly downregulated in T1DM mice. This change was reversed by rutin treatment (Fig. 4A and Supplementary Table 4). The Simpson and Shannon indexes were not significantly different among the groups (Fig. S2 and Supplementary Table 5). According to the principal coordinate analysis (PCoA) of weighted UniFrac and unweighted UniFrac based on the β-diversity analysis, it was revealed that gut microbiota in the ND group, T1DM group, T1DM-Rutin100 group and T1DM-Rutin200 group were clearly distinguished, suggesting that there were different microbiota structures among those groups (Fig. 4B, Fig. S3, Supplementary Table 6). In T2DM mice, a total of 2,017,480 original sequences were obtained, and an average of 106,183 ± 6479 sequences were acquired for each sample. A total of 1,562,265 effective sequences were generated with an average of 82,224 ± 499 sequences for each sample after quality screening. The abundance grade curves and species accumulation curves were drawn as shown in Fig. S4. The α-diversity analysis showed that the Chao1 index was significantly decreased in the T2DM group, indicating lower gut microbiota diversity in this group, which was reversed by rutin (Fig. 4C and Supplementary Table 7). No significant difference in the Simpson and Shannon indexes were observed among the different groups (Fig. S5 and Supplementary Table 8). The β-diversity calculated by unweighted UniFrac distances showed that the gut microbiota was clearly distinguished in the ND, T2DM, T2DM-Rutin100, and T2DM-Rutin200 groups, suggesting different microbiota among those four groups (Fig. 4D, Fig. S6, and Supplementary Table 9). The results showed that rutin changed the overall community structure of the gut microbiota in both T1DM and T2DM mice.

Rutin altered the gut microbiota diversity in T1DM and T2DM mice. (A) The Chao 1 index in the α-diversity analysis in T1DM mice. (B) β-diversity analysis in the ND, T1DM, T1DM-Rutin100, and T1DM-Rutin200 groups. (C) The Chao 1 index in α-diversity analysis in T2DM mice. (D) The β-diversity analysis in the ND, T2DM, T2DM-Rutin150, and T2DM-Rutin200 groups. One-way ANOVA was used for statistical comparisons of different groups. Values are means ± SD (n = 5): *p < 0.05, **p < 0.01, ***p < 0.001.
Rutin regulated the composition of the gut microbiota in diabetic mice
To study whether rutin regulates the composition of the gut microbiota in diabetic mice. We performed 16S rRNA sequencing for each mouse fecal sample. Firmicutes and Bacteroidetes at phylum level accounted for approximately 82.5% in the T1DM group (Fig. 5A and Supplementary table 10). Our data showed that the Bacteroidetes/Firmicutes (B/F) ratio in T1DM group was significantly lower than that in the ND group, but the B/F ratios in both the T1DM-Rutin100 group and T1DM-Rutin200 were higher than in the T1DM group. Compared to ND group, the relative abundance of Proteobacteria was significantly increased in the T1DM group, which was significantly reversed by rutin (Fig. 5B). LEfSe analysis was conducted to identify differentially abundant bacterial taxa among the 4 groups (LDA score > 2) (Fig. 5C and Supplementary Table 11). T1DM showed selective enrichment in 11 communities including f_Enterobacteriaceae, g_Helicobacter, g_Escherichia, and g_Mycoplasma, while T1DM-Rutin100 showed selective enrichment in 4 communities, including f_Bacteroidaceae, g_Bacteroides, o_Rickettsiales and g_Anaerotruncus. Moreover, two microbial communities were enriched in the TIDM-Rutin200 group, including o_Bacteroidales and f_Rikenellaceae. At the order level, the relative abundances of Lactobacillales, Coriobacteriales, and Campylobacterales were increased in the T1DM group, but decreased after rutin treatment. Furthermore, the relative abundances of Bacteroidales and Verrucomicrobiales were increased by rutin (Fig. S7 and Supplementary Table 12). The heatmap clustering analysis showed that compared to the ND group, the relative abundances of Dorea, Coprococcus, Enterobacter, Flexispira, Helicobacter, Proteus, Streptococcus, Coprobacillus, Escherichia and Lactococcus were increased in the T1DM group, which was reversed by rutin. Furthermore, rutin increased the relative abundances of Odoribacter, Rikenella, Staphylococcus, Akkermansia, Alisipes, Lactobacillus, Prevotella and Roseburia. These bacteria were negatively associated with diabetes (Fig. 5D). Additionally, Spearman’s correlation analysis showed that the gut microbiota was correlated with metabolic parameters related to diabetes (Fig. 5E and Supplementary Table 13). Serum glucose was positively correlated with the relative abundance of Flexispira and Helicobacter. Serum TC, TG, LDL-C, TNF-α, and IL-6 levels were positively correlated with the relative abundances of Enterobacteriaceae, Lactobacillus, and Lactococcus. The abundance of Sutterella was negatively associated with insulin.

Effects of rutin on the gut microbiota composition in T1DM mice. (A) The relative abundances of gut microbiota constituents at the phylum level. (B) Bacteroidetes/Firmicutes in feces of mice and relative abundance of Proteobacteria at the phylum level. (C) LEfSe analysis of gut microbiota in the ND, T1DM, T1DM-Rutin100, and T1DM-Rutin200 groups. LDA scores > 2 and P < 0. 05 are the taxonomic groups with significant differences between groups. (D) Heatmap of the relative abundance of gut microbiota at the genus level. Each column in the plot represents a sample, and each row represents the community structure. The color represents the relative abundance of the species. The heatmap was created using Sangerbox 3.0 (http://vip.sangerbox.com/). (E): Spearman’s correlation analysis of gut microbiota and metabolic parameters. Values are means ± SD (n = 5): *p < 0.05, **p < 0.01.
In the experiments with T2DM mice, Firmicutes and Bacteroidetes at the phylum level accounted for approximately 83% in the T2DM group (Fig. 6A and Supplementary table 14). The B/F ratio in T2DM group was lower than that in the ND group, however, rutin significantly reversed this change. The relative abundance of Proteobacteria in the T2DM group was significantly higher than ND group, and rutin treatment significantly decreased the relative abundance of Proteobacteria (Fig. 6B). Furthermore, the LEfSe analysis showed that T2DM had selective enrichment in 14 communities, including f_Desulfovibrionaceae, o_Clostridiales, c_Erysipelotrichi, g_Bilophila and p_Firmicutes, while the T2DM-Rutin100 group had selective enrichment in 14 communities including f_Lachnospiraceae, g_Blautia, g_Anoxybacillus and g_Coprobacillus. However, 19 communities were selectively enriched in the T2DM-Rutin200 group, such as g_Bacteroides, g_Oscillospira, p_Verrucomicrobia, g_Akkermansia and g_Butyricimonas (Fig. 6C and Supplementary Table 15). Rutin decreased the upregulated relative abundances of Erysipelotrichales, Lactobacillales and Bacillales in T2DM mice at the order level but increased those of Bacteroidales, Burkholderiales and Coriobacteriales (Fig. S8). The relative abundances of Helicobacter, Allobaculum, Parabacteroides, Mucispirillum, Enterococcus, Lactococcus, Proteus, Staphylococcus and Enterobacter were upregulated in T2DM mice at the genus level, which were reversed by rutin treatment. Additionally, rutin increased the relative abundances of Klebsiella, Bacteroides, Lactobacillus, Akkermansia, Pseudoramibacter, Eubacterium, Roseburia, Butyricimonas and Alisipes (Fig. 6D and Supplementary Table 16). The serum glucose level was positively correlated with the relative abundance of Mogibacteriaceae. The relative abundances of Clostridiales, Mogibacteriaceae, and Lachnospiraceae were positively correlated with LDL-C, and IL-6 levels, while those of Mogibacteriaceae and Lachnospiraceae were negatively associated with HDL-C (Fig. 6E and Supplementary Table 17). These findings indicated that rutin could regulate the composition of the gut microbiota in both T1DM and T2DM mice.

Effects of rutin on the gut microbiota composition in T2DM mice. (A) The relative abundances of gut microbiota constituents at the phylum level. (B) Bacteroidetes/Firmicutes in feces of mice and relative abundance of Proteobacteria at the phylum level. (C) LEfSe analysis of gut microbiota in the ND, T2DM, T2DM-Rutin100, and T2DM-Rutin200 groups, LDA scores > 2 and P < 0. 05 are the taxonomic groups with significant differences between groups. (D) Heatmap of the relative abundance of gut microbiota at the genus level. Each column in the plot represents a sample, and each row represents the community structure. The color represents the relative abundance of the species. The heatmap was created using Sangerbox 3.0 (http://vip.sangerbox.com/). (E): Spearman’s correlation analysis of gut microbiota and metabolic parameters. Values are means ± SD (n = 5): *p < 0.05, **p < 0.01.
Discussion
Accumulating evidence suggests that the gut microbiota plays an important role in regulating obesity, diabetes, colon cancer and nonalcoholic fatty liver disease8. Rutin is a type of flavonoid glycoside enriched in tea and buckwheat that has multiple biological activities, such as antioxidant, anti-inflammatory, antidiabetic and free radical scavenging activities. A previous study suggested that the combination of rutin and vitamin C can effectively improve oxidative stress and lower serum glucose in patients with T2DM22. Rutin can not only prevent STZ-induced oxidative damage, but also protect islet β cells to increase insulin secretion and therefore lower serum glucose36. However, whether rutin alleviates colon lesions and regulates the gut microbiota in diabetic mice is entirely unknown. Our study found that rutin could significantly improve the levels of serum glucose, TC, TG, and LDL-C in diabetic mice and increase HDL-C levels. Chronic inflammation caused by high serum glucose and excessive visceral fat accumulation could increase the risk of many different cancers, such as colon cancer and liver cancer37. We found that the villi of colon tissues were arranged disorderly and easy to break, along with many vacuolar changes and damaged crypts in diabetic mice, this phenomenon is might strongly associated with the loss of tight junction proteins, which are the expression levels of tight junction proteins in patients with diabetes are abnormal, which play an important role in maintaining the structure of colon38, however, whether rutin affects the expression levels of tight junction proteins in colon tissue of diabetic mice, we will further explore this hypothesis in the future. Disorders of the gut microbiota were reported to be closely related to the occurrence and development of colonic diseases39,40. We determined the effects of rutin on gut microbiota dysbiosis and colon lesions in diabetic mice and found that rutin significantly alleviated colon lesions in T1DM and T2DM mice (Fig. S9).
It is known that the gut microbiota condition affects the pathophysiological states of metabolic diseases. The abnormal gut microbiota in diabetic mice might be related to the occurrence of obesity, diabetes, inflammation and pathological microenvironments41,42. Reduced bacterial function diversity and low community stability were characterized in diabetic mice43. We hypothesized that the antidiabetic effects of rutin and its ability to repair colon lesion damage in diabetic mice might be closely related to the regulation of the gut microbiota. In our study, the abundance grade curves and species accumulation curves indicated that most of the gut microbial diversity was captured with the current sequencing depth in each sample. However, the rank abundance curve estimate for diversity might not be fully representative of reality. In addition, Chao1 and Shannon indexes were higher in the rutin treatment groups than in the T1DM, or T2DM group. Diversity analysis showed differences in gut microbiota distribution among the groups. However, there was still a difference between the intestinal flora after rutin treatment and that of nondiabetic mice. It is possible that the dosage of rutin is not enough to achieve the therapeutic effect, which might have an auxiliary therapeutic effect. The above results showed that rutin significantly changed the structure of the intestinal flora in diabetic mice, indicating a novel pathway through which rutin alleviates diabetes-related symptoms. It has been reported that obesity and metabolic syndromes are associated with changes in gut microbiota composition, including a decreased Bacteroidetes/Firmicutes (B/F) ratio and relative abundance of Proteobacteria44. It was also found that the abundance of Firmicutes and Bacteroidetes in diabetic mice was related to hyperglycemia and inflammatory state45. In accordance with previous studies, our results showed that rutin effectively increased Bacteroidetes and decreased Firmicutes, along with an increased ratio of Bacteroidetes to Firmicutes. The increase in the abundance of Proteobacteria could promote the endotoxin content, thus leading to the occurrence and development of chronic inflammatory diseases46. We also demonstrated that rutin reversed the relative abundance of Proteobacteria in diabetic mice. Furthermore, previous studies suggested that Erysipelotrichales were correlated with the development of obesity, systemic inflammation and metabolic diseases47,48. In our study, the Erysipelotrichaeae abundance in T2DM mice was much higher than that in the control ND group, which was reversed by rutin. Cani et al. reported a positive relationship between Escherichia and the occurrence and development of diabetes and obesity49, whereas rutin reduced the presence of Helicobacter, Enterobacter, and Escherichia in diabetic mice. The increase in the F/B ratio was related to obesity induced by a high-fat diet in mice50. Rutin could increase the B/F ratio and reduce the content of Proteobacteria, which was closely related to inflammation.
It was reported that Akkermansia was a mucin-degrading bacterium and improved insulin resistance51. Our data showed that rutin increased Akkermansia abundance. The relative abundances of some bacteria, such as Bilophila and Mucispirillum, showed a significant positive correlation with the development of diabetes47,52. Therefore, the protective effect of rutin on diabetic symptoms might be due to the decrease in Mucispirillum, Helicobacter, Enterobacter, Proteus, and Streptococcus abundances. In addition, rutin increases the relative abundances of bacteria, such as Odoribacter, Rikenella, Akkermansia, Alistipes and Roseburia, which impairs the development of diabetes. Spearman correlation analysis showed that key communities such as Enterobacteriaceae, Lactobacillus and Sutterella were related to the main metabolic parameters of diabetic mice. It is worth noting that blood lipids and inflammatory factors in diabetes mice are positively correlated with Lactobacillus, it is possible that the increase of Lactobacillus abundance can protect the damage of inflammation to the body, and may play an important role in immune defenses. Sutterella has anti-inflammatory properties and is negatively correlated with the content of IL-653,54. Therefore, the reduction in inflammation in diabetic mice treated with rutin might be due to the increase in Sutterella abundance, which was positively related to the decrease in serum IL-6, suggesting that the effect of rutin on the flora might be related to metabolic disorders. Several studies have demonstrated that short-chain fatty acids (SCFAs) are the main metabolites of gut microbiota that play a critical role in regulating metabolic syndrome and maintaining energy homeostasis and host insulin sensitivity55,56. Short chain fatty acids such as acetic acid, propionic acid and butyric acid play an important role in improving chronic inflammatory diseases and promoting colon cell health57. Alisipes, Roseburia, Rikenella and Odoribactor are bacteria that produce butyric acid, propionic acid and butyric acid in the intestine58,59. SCFAs produced by these microorganisms can provide energy for the host, improve the acidic environment in the colon, inhibit the production of inflammatory factors, and finally repair mucositis. Our study found that rutin significantly increased the abundances of Alisipes, Odoribacter, Roseburia, and Rikenella, which might lead to increased production of SCFAs that act beneficially on colon villi in our experimental diabetic mice. The recovery of colon damage by rutin might therefore be due to the enrichment of SCFA-producing bacteria. Targeted recovery of these SCFA products will provide a new therapeutic approach for diabetes patients.
Conclusions
In conclusion, rutin exhibited an antidiabetic effect and improved colon lesions in diabetic mice, possibly by regulating the gut microbiota community structure and composition, which might be a potential new strategy for treating diabetic patients, protecting intestinal health and/or preventing colon cancer carcinogenesis. Further study is needed to reveal the detailed mechanisms through which rutin regulates the gut microbiota in diabetic mice.
Data availability
The datasets generated and/or analysed during the current study are available in the Zenodo repository, https://zenodo.org/record/7404629, doi: https://doi.org/10.5281/zenodo.7404629.
References
Sharifuddin, Y., Chin, Y. X., Lim, P. E. & Phang, S. M. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 13, 5447–5491 (2015).
Xu, Yu. Prevalence and control of diabetes in Chinese adults. JAMA 310, 948–959 (2013).
Gonzalez, N. et al. update on the relationship between diabetes and colorectal cancer: Epidemiology, potential molecular mechanisms and therapeutic implications. Oncotarget 8(2017), 18456–18485 (2017).
Ibrahim, K. S., Craft, J. A., Biswas, L., Spencer, J. & Shu, X. Etifoxine reverses weight gain and alters the colonic bacterial community in a mouse model of obesity. Biochem. Pharmacol. 180, 114151 (2020).
Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 11, 639–647 (2013).
Yan, H. et al. Intake of total saponins and polysaccharides from Polygonatum kingianum affects the gut microbiota in diabetic rats. Phytomedicine 26, 45–54 (2017).
Qin, J. et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 490, 55–60 (2012).
Ruth, E. et al. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. U. S. A. 102, 11070–11075 (2005).
De Goffau, M. C. et al. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia 57, 1569–1577 (2014).
Phimister, E. G., Lynch, S. V. & Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 375, 2369–2379 (2016).
Martel, J. et al. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat. Rev. Endocrinol. 13, 149–160 (2017).
Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 203, 231–245 (2016).
Lee, D. G. et al. Effect of rutin from tartary buckwheat sprout on serum glucose-lowering in animal model of type 2 diabetes. Acta Pharm. 66, 297–330 (2016).
Jin, H.-R., Jin, Yu. & Choi, S.-J. Hydrothermal treatment enhances antioxidant activity and intestinal absorption of rutin in tartary buckwheat flour extracts. Foods 9, 1–14 (2019).
Aditya, G. & Ajay, S. The pharmacological potential of rutin. Saudi Pharm. J. 25, 149–164 (2017).
Nikfarjam, B. A. et al. Treatment with rutin—A therapeutic strategy for neutrophil-mediated inflammatory and autoimmune diseases: Anti-inflammatory effects of rutin on neutrophils. J. Pharmacopunct. 20, 52–56 (2017).
Silva, A. B. D. et al. The flavonoid rutin and its aglycone quercetin modulate the microglia inflammatory profile improving antiglioma activity. Brain Behav. Immun. 85, 170–185 (2020).
Habtemariam, S. & Lentini, G. The therapeutic potential of rutin for diabetes: An update. Mini Rev. Med. Chem. 15, 524–528 (2015).
Hasanein, P., Emamjomeh, A., Chenarani, N. & Bohlooli, M. Beneficial effects of rutin in diabetes-induced deficits in acquisition learning, retention memory and pain perception in rats. Nutr. Neurosci. 23, 563–574 (2020).
Amjadi, S. et al. Nanophytosomes for enhancement of rutin efficacy in oral administration for diabetes treatment in streptozotocin-induced diabetic rats. Int. J. Pharm. 610, 121208 (2021).
Attia, R. A., Abdel Fattah, S. & Nasralla, M. M. Concomitant administration of sitagliptin and rutin improves the adverse hepatic alterations in streptozotocin-induced diabetes mellitus in albino rats: An overview of the role of alpha smooth muscle actin. Folia Morphol. (Warsz) 80, 870–880 (2021).
Ragheb, S. R., El Wakeel, L. M., Nasr, M. S. & Sabri, N. A. Impact of Rutin and Vitamin C combination on oxidative stress and glycemic control in patients with type 2 diabetes. Clin. Nutr. ESPEN 35, 128–135 (2020).
Nafees, S. et al. Synergistic interaction of rutin and silibinin on human colon cancer cell line. Arch. Med. Res. 49, 226–234 (2018).
Guon, T. E. & Chung, H. S. Hyperoside and rutin of Nelumbo nucifera induce mitochondrial apoptosis through a caspase-dependent mechanism in HT-29 human colon cancer cells. Oncol. Lett. 11, 2463–2470 (2016).
Asfour, M. H. & Mohsen, A. M. Formulation and evaluation of pH-sensitive rutin nanospheres against colon carcinoma using HCT-116 cell line. J. Adv. Res. 9, 17–26 (2017).
Liu, G. M., Liang, L., Yu, G. Y. & Li, Q. H. Pumpkin polysaccharide modifies the gut microbiota during alleviation of type 2 diabetes in rats. Int. J. Biol. Macromol. 115, 711–717 (2018).
Huo, M. et al. Rutin promotes pancreatic cancer cell apoptosis by upregulating miRNA-877-3p expression. Molecules 27, 2293–2308 (2022).
Cheng, W. et al. Tartary buckwheat flavonoids improve colon lesions and modulate gut microbiota composition in diabetic mice. Evid. Based Complement Alternat. Med. 16, 4524444 (2022).
Lertpatipanpong, P. et al. The anti-diabetic effects of NAG-1/GDF15 on HFD/STZ-induced mice. Sci. Rep. 11, 15027 (2021).
Matboli, M. et al. Caffeic acid attenuates diabetic kidney disease via modulation of autophagy in a high-fat diet/streptozotocin- induced diabetic rat. Sci. Rep. 7, 2263 (2017).
Jiang, N. et al. HIF-1alpha ameliorates tubular injury in diabetic nephropathy via HO-1-mediated control of mitochondrial dynamics. Cell Prolif. 53, e12909 (2020).
Wu, L. et al. Analysis of the protective mechanism of liraglutide on retinopathy based on diabetic mouse model. Saudi J. Biol. Sci. 26, 2096–2101 (2019).
Choi, J. Y. et al. The n-butanol fraction and rutin from tartary buckwheat improve cognition and memory in an in vivo model of amyloid-β-induced Alzheimer’s disease. J. Med. Food 18, 631–41 (2015).
Fan, H. & Gao, Z. The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways. Phytomedicine. 58, 152864 (2019).
R Core Team, R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. https://www.R-project.org. 2018.
Mascaraque, C. et al. Rutin has intestinal anti-inflammatory effects in the CD4þ CD62Lþ T cell transfer model of colitis. Pharmacol. Res. 90, 48–57 (2014).
Cade, W. T. Diabetes-related microvascular and macrovascular diseases in the physical therapy setting. Phys. Ther. 88, 1322–1335 (2008).
Jiang, Q. W. et al. Diabetes inhibits corneal epithelial cell migration and tight junction formation in mice and human via increasing ROS and impairing Akt signaling. Acta Pharmacol. Sin. 40, 1205–1211 (2019).
Gao, R. et al. Gut microbiota dysbiosis signature is associated with the colorectal carcinogenesis sequence and improves the diagnosis of colorectal lesions. J. Gastroenterol. Hepatol. 4, 1–12 (2020).
Yang, Y. et al. Fusobacterium nucleatum increases proliferation of colorectal cancer cells and tumor development in mice by activating toll-like receptor 4 signaling to nuclear factor−κB, and up-regulating expression of MicroRNA-21. Gastroenterology 152, 851–866 (2017).
Verdam, F. J. et al. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity (Silver Spring) 21, 607–615 (2013).
Song, H. et al. Red pitaya betacyanins protects from diet-induced obesity, liver steatosis and insulin resistance in association with modulation of gut microbiota in mice. J. Gastroenterol. Hepatol. 31, 1462–1469 (2016).
Zhou, K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. J. Funct. Foods 33, 194–201 (2017).
Cani, P. D. et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56, 1761–1772 (2007).
Martinez, I. et al. Diet-induced alterations of host cholesterol metabolism are likely to affect the gut microbiota composition in hamsters. Appl. Environ. Microbiol. 79, 516–524 (2013).
Wang, J. et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 9, 1–15 (2015).
Everard, A. et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 60, 2775–2786 (2011).
Dao, M. C. et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 65, 426–436 (2016).
de la Cuesta-Zuluaga, J. et al. Metformin is associated with higher relative abundance of mucin-degrading akkermansia muciniphila and several short-chain fatty acid-producing microbiota in the gut. Diabetes Care 40, 54–62 (2017).
Tilg, H. & Moschen, A. R. Microbiota and diabetes: An evolving relationship. Gut 63, 1513–1521 (2014).
Xu, H. et al. Alterations of gut microbiome in the patients with severe fever with thrombocytopenia syndrome. Front. Microbiol. 9, 2315 (2018).
Turnbaugh, P. J. et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027–1031 (2006).
Koh, A., De Vadder, F., Kovatcheva-Datchary, P. & Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 165, 1332–1345 (2016).
Hartstra, A. V., Bouter, K. E., Backhed, F. & Nieuwdorp, M. Insights into the role of the microbiome in obesity and type 2 diabetes. Diabetes Care 38, 159–165 (2015).
Fu, X., Liu, Z., Zhu, C., Mou, H. & Kong, Q. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit. Rev. Food Sci. Nutr. 59, 130–152 (2019).
Liu, C. S. et al. Gegen Qinlian decoction treats diarrhea in piglets by modulating gut microbiota and short-chain fatty acids. Front. Microbiol. 10, 1–11 (2019).
Shi, H., Chang, Y., Gao, Y., Wang, X. & Tang, Q. J. Dietary fucoidan of Acaudina molpadioides alter gut microbiota and mitigate intestinal mucosal injury induced by cyclophosphamide. Food Funct. 8, 3383–3393 (2017).
Dai, S. et al. Effects of lactic acid bacteria-fermented soymilk on Isoflavone metabolites and short-chain fatty acids excretion and their modulating effects on gut microbiota. J. Food Sci. 84, 1854–1863 (2019).
Ruqin Lin, Yu. et al. Lactobacillus rhamnosus GG supplementation modulates the gut microbiota to promote butyrate production, protecting against deoxynivalenol exposure in nude mice. Biochem. Pharmacol. 175, 113868 (2020).
Acknowledgements
We acknowledge the College of Life and Environmental Science, Wenzhou University, Wenzhou, China, for providing the laboratory facilities.
Funding
This research was funded by the National Key R & D Program of China (Grant No. 2017YFE0117600) and the Key Research and Development Project of Zhejiang Province (Grant No. 2022C04003).
Author information
Authors and Affiliations
Contributions
Conceptualization, C.F.C. and W.W.C.; methodology, C.F.C. and W.W.C.; investigation, C.F.C., W.W.C. and T.T.S.; data curation, W.W.C.; writing—original draft preparation, W.W.C.; M.L.Z., Y.L.L., and Z.Y.L. revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cai, C., Cheng, W., Shi, T. et al. Rutin alleviates colon lesions and regulates gut microbiota in diabetic mice. Sci Rep 13, 4897 (2023). https://doi.org/10.1038/s41598-023-31647-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-31647-z
This article is cited by
-
A Dual Therapeutic Approach to Diabetes Mellitus via Bioactive Phytochemicals Found in a Poly Herbal Extract by Restoration of Favorable Gut Flora and Related Short-Chain Fatty Acids
Applied Biochemistry and Biotechnology (2024)
-
High-performance thin-layer chromatography method development and validation for quantification of rutin in different parts of Capparis zeylanica Linn. plant
JPC – Journal of Planar Chromatography – Modern TLC (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
