
Abstract
Occurrence of multiple abiotic stresses in a single crop season has become more frequent than before. Most of the traditional donors possessing tolerance to abiotic stresses are tall, low-yielding with poor grain quality. To facilitate efficient use of complex polygenic traits in rice molecular breeding research, we undertook development of introgression lines in background of high-yielding, semi-dwarf varieties with good grain quality. The study reports the development and evaluations of over 25,000 introgression lines in eleven elite rice genetic backgrounds for improvement of yield under multiple abiotic-stresses such as drought, flood, high/low temperature. The developed introgression lines within each genetic background are near isogenic/recombinant inbred lines to their recipient recurrent parent with 50 to 98% background recovery and additionally carry QTLs/genes for abiotic stresses. The multiple-stress tolerant pyramided breeding lines combining high yield under normal situation and good yield under moderate to severe reproductive-stage drought, semi-dwarf plant type with good grain quality traits have been developed. The introgression lines in dwarf backgrounds open new opportunity to improve other varieties without any linkage drag as well as facilitate cloning of QTLs, identification and functional characterization of candidate genes, mechanisms associated with targeted QTLs and the genetic networks underlying complex polygenic traits.
Similar content being viewed by others

Introduction
The worldwide rice production is now facing the problem of a rapidly changing climate manifested as increased frequency and intensity of occurrence of multiple abiotic stresses. Climatic factors, such as drought, flood and extreme temperatures (low temperature and heat) are the main detrimental abiotic environmental stresses restricting plant growth and crop productivity1. The environmental stresses affect ~ 30% of 700 million poor people living in rainfed rice growing regions of Asia. Mega rice varieties are highly susceptible to these abiotic stresses, thereby suffering high yield losses. There is an urgent need to develop climate-change ready rice exploiting recent advances in genomics. The important goal of molecular quantitative genetics is to identify the genomic loci and specific alleles underlying the genetic variation in quantitative traits of interest. The recent advances in marker-assisted breeding have streamlined the food and feeder crops breeding programs2,3. In the past two decades, many quantitative trait loci (QTLs) associated with agronomic, physiological, grain quality, biotic and abiotic stress tolerance have been detected and some of them were cloned by using molecular markers4,5,6,7,8,9.
Before the advent of the genomics era in the early 2000s, many genes and QTLs governing abiotic stress tolerance were identified but they were not utilized successfully in breeding programs through marker assisted selection. Molecular marker-assisted introgression/QTL pyramiding has allowed fast and accurate selection and development of elite breeding lines with the target traits of interest. Despite the major advancements in modern era of genomics, moving from genomic region to the causal alleles remain a major challenge in genetics research. Pyramiding of favourable QTLs/genes present in traditional varieties provides a precise way to improve the varieties that excel in large number of characteristics10,11,12,13,14,15,16,17.
To improve grain yield, quality and biotic/abiotic resistance; development of introgression lines exploiting genomics-assisted breeding is mandatory. Developed introgression lines could be ideal and very useful material for QTL validation, fine mapping of QTL, gene discovery and in investigating the genetic and physiological mechanism in crop plants. Marker-assisted QTL introgression/pyramiding may be a suitable approach to get the desired phenotypic variance in improving grain yield under stress conditions in rice18. Near Isogenic lines provides an opportunity to validate the effect of single or few major effect QTLs in addition to an increase in the recovery of recurrent parent genome19. Utilizing the existing molecular markers for introgression/pyramiding of QTLs through marker-assisted selection is most desirable step in improving existing mega varieties and to develop new abiotic stress tolerance rice breeding lines/varieties. The introgression lines harboring desired QTLs/genes for multiple abiotic/biotic stresses in elite background with better performance may serve as valuable material for direct release to the farmer’s field. They may also serve as useful resource to study the epistatic interactions among introgressed QTLs and with the genetic background17,20,21,22.
Several major and consistent effect rice grain yield QTLs for reproductive stage drought stress; qDTY1.1 (on chromosome 1), qDTY2.1, qDTY2.2, qDTY2.3 (on chromosome 2), qDTY3.1, qDTY3.2 (on chromosome 3), qDTY4.1 (on chromosome 4), qDTY6.1, qDTY6.2 (on chromosome 6), qDTY9.1 (on chromosome 9), qDTY10.1 (on chromosome 10) and qDTY12.1 (on chromosome 12) have been identified to be present in traditional germplasm accession and their effect validated in different rice genetic backgrounds such as Swarna, IR64, MTU1010, TDK1-Sub1, MRQ74, MR219, Savitri and Vandana using different molecular marker technology15,18,23,24,25,26,27,28.
Submergence tolerance is controlled by a strong effect QTL Sub1 from FR13A29, which can provide tolerance to rice up to 14 days of complete submergence. Flood tolerant version of many mega rice varieties such as Ciherang, Swarna, IR64 and TDK1 were developed using marker-assisted backcross breeding30,31,32.
The QTLs for low temperature tolerance at seedling, vegetative and booting stages33,34,35,36,37,38 along with the QTLs for tolerance to low temperature at reproductive stage39,40,41,42,43,44,45 have been identified in different populations. A total of 5 low temperature tolerance QTLs in a study with 122 RILs in F13 generation from a cross between Danteshwari and Dagaddeshi have been identified on chromosomes 1, 3, 6, 9 and 1246. The QTLs for heat tolerance in doubled haploid population47, recombinant inbred lines (RILs), back cross inbred lines (BILs) populations48,49,50,51,52,53,54,55,56,57 and chromosomal substitution lines58 have been reported.
Till date, many QTLs have been reported in different studies but very few have been deployed in marker-assisted breeding programs. Other than the stability of the QTLs effect across different genetic backgrounds and environments, one of the most important reason for this has been linkage drag of the identified QTLs with traits of less economic importance such as low grain yield, tall plant height, and poor grain quality59. The existing linkage drag need to be broken and QTLs need to be introgressed in backgrounds those possess other traits of economic importance. The aim of the study was to introgress newly identified QTLs to develop marker-assisted derived introgression lines in the background of popular cultivated varieties IR64, Swarna, MR219, TDK1, Samba Mahsuri, Kalinga 3, Vandana, Anjali and MTU1010 for tolerance to multiple abiotic stresses such as drought, low temperature and high temperature and make available appropriate genetic material to undertake more efficient, less cumbersome marker assisted introgression breeding programs as well as basic studies to clone genes, identify interactions and physiological mechanisms.
Results
Development of introgression lines
The drought tolerant introgression lines in background of a total of 11 popular lowland and upland adapted rice varieties free from the linkage drag with uneconomic traits were developed. The introgression schemes used to develop the dual drought-submergence tolerant introgression lines in background of lowland adapted rice varieties such as Swarna-Sub1, IR64-Sub1 and TDK1-Sub1 are summarised in Figs. S1, S2 and S3, respectively. The introgression schemes used to develop the drought tolerant introgression lines in background of lowland adapted rice varieties such as Savitri, Samba Mahsuri, MR219, MTU1010 and Swarna are summarised in Figs. S4, S5, S6, S7 and S8, respectively. The introgression schemes used to develop the introgression lines in background of upland adapted rice varieties such as Kalinga III, Vandana and Anjali are summarised in Figs. S9, S10, and S11, respectively. In addition, the multiple stress-tolerant; low temperature, heat and drought tolerant introgression lines in background of MTU1010 were developed.
The study generated a total of 25,000 introgression lines and evaluated 13,343 introgression lines under non-stress, 10,886 under reproductive stage drought stress, 400 under high and low temperature stress.
Phenotypic characterization of introgression lines in different backgrounds
The phenotypic screening of the developed introgression lines were carried out across seasons, generations and environments. The significant phenotypic differences were observed among the recipient parents and introgression lines in all backgrounds under different conditions such as, reproductive stage drought stress, high and low temperature stress. The results of mean grain yield of parents and introgression lines and least significant difference (LSD0.05) of trial mean of introgression lines under non-stress (control), reproductive stage drought stress, high and low temperature stress are summarized in Table S1. The mean grain yield performance of introgression lines in comparison with the respective recurrent parent under non-stress and reproductive stage drought stress across seasons at IRRI, Philippines and SAH (Hyderabad, India) is presented in Figs. S1, S2, S3, S4, S5, S6, S7, S8, S9, S10, and S11.
Phenotypic variability in multiple stress tolerant lowland adapted rice varieties
The phenotypic evaluations of a total of 9003 breeding lines in 58 experiments including 38 at IRRI, Philippines and 20 at IRRI, SAH were carried out between 2010–2017 (Table 1) in background of popular lowland adapted high-yielding varieties to validate the initial hypothesis of breakage of linkage drag and maintaining the effect of QTLs in the high yielding backgrounds. The mean grain yield of drought-submergence tolerant introgression lines in Swarna-Sub1 background varied from 3309 to 7878 kg ha-1 under non-stress and 634 to 2691 kg ha-1 under the reproductive stage drought stress (Table S1, Fig. S1). The variability for grain yield in IR64-Sub1 introgression lines ranged from 3024 to 7181 kg ha-1 under non-stress and 277 to 2998 kg ha-1 under the reproductive stage drought stress (Table S1, Fig. S2) whereas the mean grain yield in TDK1-Sub1 background ranged from 2715 to 6091 kg ha-1 and 198 to 1916 kg ha-1 under control non-stress and the drought stress, respectively (Table S1, Fig. S3). The variability for mean grain yield in low temperature-high temperature and drought tolerant introgression lines in MTU1010 background ranged from 5026 to 5176 kg ha-1 under non-stress, 1927 to 1985 kg ha-1 under the reproductive stage drought stress and 3585 to 3611 kg ha-1 under high temperature stress at reproductive stage (Table S1).
Phenotypic variability in drought tolerant lowland adapted rice varieties
The phenotypic and genotypic evaluations of total 9632 breeding lines in 75 experiments including 42 at IRRI, Philippines and 33 at IRRI, SAH were carried out between 2010–2017 (Table 1) to develop introgression lines in background of popular lowland adapted high-yielding varieties. The mean grain yield of Samba Mahsuri introgression lines varied from 2395 to 8133 kg ha-1 and 227 to 2731 kg ha-1 under non-stress and the reproductive stage drought stress respectively (Table S1, Fig. S4) and in Savitri background from 3428 to 6125 kg ha-1 and 293 to 800 kg ha-1 under non-stress and reproductive stage drought stress respectively (Table S1, Fig. S5). The mean grain yield of introgression lines in MR219 background ranged from 4585 to 8294 kg ha-1 under non-stress and 486 to 2683 kg ha-1 under reproductive stage drought stress (Table S1, Fig. S6). The variability for mean grain yield in MTU1010 background ranged from 4661 to 9698 kg ha-1 under non-stress and 244 to 2970 kg ha-1 under reproductive stage drought stress (Table S1, Fig. S7). The grain yield variability in Moroberekan/3*Swarna introgression lines ranged from 4359 to 8918 kg ha-1 and 341 to 1648 kg ha-1 under control non-stress and reproductive stage drought stress, respectively (Table S1, Fig. S8).
Phenotypic variability in drought tolerant upland adapted rice varieties
The phenotypic evaluations of a total of 5684 breeding lines in 48 experiments including 33 at IRRI, Philippines and 15 at IRRI, SAH were carried out between 2010–2017 (Table 1) to develop introgression lines in background of upland adapted high-yielding rice varieties. The mean yield of Kalinga III introgression lines ranged from 2058 to 4790 kg ha-1 under non-stress and from 221 to 3253 kg ha-1 under reproductive stage drought stress (Table S1, Fig. S9). The mean grain yield of Vandana introgression varied from 2118 to 6072 kg ha-1 and 286 to 2408 kg ha-1 under non-stress and reproductive stage drought stress, respectively (Table S1, Fig. S10). The introgressed lines in Anjali background showed grain yield variability ranged from 2925 to 5121 kg ha-1 and 83 to 2141 kg ha-1 under control non-stress and reproductive stage drought stress, respectively (Table S1, Fig. S11).
Genotypic characterization of introgression lines in different backgrounds
The genotypic selection (foreground and background selection) across generation advancements in all backgrounds were carried out to identify the high yielding stress tolerant promising lines with qDTY and qDTY combinations with high recurrent parent genome recovery (RPG) and free from linkage drag. The foreground/background selections were carried out and only the lines with desired phenotype and genotype (qDTY/qDTY combinations) were advanced to the next generation. The detailed information on the number of introgressed lines that were characterized genotypically in all the studied background in each season is presented in Table S1. The recurrent parent genome recovery (RPG) in the selected promising introgression lines carrying qDTYs in Swarna-Sub1 background ranged from 93 to 98%; 89 to 93% in IR64-Sub1 background; 89 to 90% in TDK-Sub1 background; 50 to 65% in Samba Mahsuri background; 88 to 89% in Savitri background and 66 to 73% in MR219 background (Table 1).
Performance of lowland adapted promising introgression lines under different stresses
The introgression lines harboring qDTY and/or different qDTY combinations showed grain yield advantage under the reproductive stage drought stress, high temperature stress and low temperature stress with no/zero yield penalty under control/non-stress in all lowland adapted backgrounds, clearly signifying the breakage of linkage with tall plant height, low yield potential for QTLs possessing such linkage drag. The drought-submergence tolerant Swarna-Sub1 introgression lines possessing qDTYs showed grain yield advantage ranging from 1004 kg ha-1 (qDTY1.1 + qDTY3.1 + Sub1) to 1786 kg ha-1 (qDTY3.1 + Sub1) under severe stress, 705 (qDTY1.1 + qDTY3.1 + Sub1) to 1482 kg ha-1 (qDTY3.1 + Sub1) under moderate stress, 902 (qDTY1.1 + qDTY2.1 + qDTY3.1 + Sub1) to 2658 kg ha-1 (qDTY3.1 + Sub1) under non-stress and 253 kg ha-1 (qDTY1.1 + qDTY2.1 + qDTY3.1 + Sub1) to 532 kg ha-1 (qDTY3.1 + Sub1) under submergence conditions over Swarna-Sub1 (Table 1, Fig. S1). Most of the Swarna-Sub1 introgression lines showed recurrent parent recovery more than 90%. A reduction of 71 to 76% and 68 to 72% in grain yield was observed in Swarna-Sub1 introgression lines under severe stress and moderate stress, respectively compared to non-stress. The PHT (plant height) was reduced by 19 to 38 cm and 14 to 23 cm under severe and moderate stresses, respectively and DTF (days to 50% flowering) increased by 3 to 4 days. In IR64-Sub1 introgression lines, the grain yield reduction under severe and moderate stresses ranged from 57 to 88% and 34 to 66% respectively compared to non-stress (Table 1). Under severe stress, DTF difference was up to 9 days and PHT difference was up to 32 cm. The grain yield improvement over the recurrent parent (IR64) ranged from 14 (qDTY4.1 + qDTY12.1) to 1568 kg ha-1 (qDTY12.1) under severe stress, 1145 (qDTY2.2 + qDTY12.1) to 2660 kg ha-1 (qDTY12.1) under moderate stress and 141 (qDTY12.1) to 1143 kg ha-1 (qDTY1.2 + qDTY12.1) under non-stress (Table 1, Fig. S2). The recurrent parent genome recovery ranged from 89 to 93% (Table 1, Fig. S2).
The grain yield advantage of introgression lines in TDK1-Sub1 background varied from 302 kg ha-1 (qDTY6.1 + qDTY6.2) to 465 kg ha-1 (qDTY3.1 + qDTY6.1 + qDTY6.2) under severe stress, 938 (qDTY6.1 + qDTY6.2) to 2656 kg ha-1 (qDTY3.1 + qDTY6.1 + Sub1) under moderate stress and 178 kg ha-1 (qDTY6.1 + qDTY6.2) to 317 kg ha-1 (qDTY3.1 + qDTY6.1 + qDTY6.2) under non-stress condition compared to the recurrent parent (Table 1, Fig. S3). The background recovery was around 90%.
The grain yield improvement of introgression lines harboring qDTY2.2 + qDTY4.1 in Samba Mahsuri background ranged from 250 to 340 kg ha-1, 166 to 1746 kg ha-1 and 115 to 1145 kg ha-1 under severe, moderate and non-stress conditions, respectively over Samba Mahsuri (Table 1, Fig. S4). Under severe stress, the introgression line possessing qDTY2.2 + qDTY3.1 + qDTY12.1 in MR219 background showed higher grain yield improvement over MR219, whereas under moderate stress and non-stress the line possessing qDTY3.1 + qDTY12.1 showed higher grain yield improvement (Table 1, Fig. S6). The large reduction in grain yield under severe stress compared to non-stress was observed in the introgression lines in TDK1-Sub1 (Fig. S3), Samba Mahsuri (Fig. S4), Savitri (Fig. S5), MTU1010 (Fig. S7) and Swarna (Fig. S8) backgrounds. Under moderate stress grain yield improvement of 252 kg ha-1 (qDTY3.1 + qDTY6.1 + qDTY6.2) over MTU1010 was observed in the drought tolerant introgression lines in MTU1010 background. The grain yield advantage under severe drought stress, moderate drought stress and control non-stress conditions was 495, 991 and 1621 kg ha-1, respectively in Swarna*3/Moroberekan introgression lines (qDTY3.2) compared to the recurrent parent (Table 1, (Fig. S7).
The stringent phenotypic selection was employed to select multiple stress tolerance lines possessing tolerance to low temperature, high-temperature and drought stress. The introgression lines IR 128762-187-1-B-B-B-B, IR 128762-83-B-B-B-B-B and IR 128,762-155-2-B-B-B-B showed better performance over other introgression lines and parents (MTU1010 and IR 91648-B-117-B-1-1) under multiple stress conditions (Table 2). The variation in low temperature tolerance at germination and seedling stage was observed. IR 128762-187-1-B-B-B-B was found to be strongly low temperature tolerant at seedling stage and tolerant at germination stage, IR 128762-83-B-B-B-B-B was moderately low temperature tolerant at seedling stage and tolerant at germination stage and IR 128762-155-2-B-B-B-B was strongly low temperature tolerant at seedling stage and tolerant at germination stage (Table 2). The yield potential of selected lines under non-stress condition ranged from 5329 to 6617 kg ha-1, under RS from 2111 to 2571 kg ha-1 and under high-temperature stress from 3694 to 4394 kg ha-1 (Table 2). The percentage reduction in grain yield (61%) under reproductive stage drought stress was more as compared to the yield reduction under reproductive stage high-temperature stress (34%) (Table 2).
Performance of upland adapted promising introgression lines under drought stress
The introgression lines harboring qDTY and/or different qDTYs combinations with on par improvement in grain yield under reproductive stage drought stress with no yield penalty under non-stress were selected in all upland adapted backgrounds. The percent grain yield reduction under SS ranged from 79 to 84% and under MS from 71 to 79% compared to NS in Kalinga III background (Table 3, Fig. S9). The drought tolerant introgression lines possessing qDTY12.1 in Vandana background showed grain yield improvement of 578 kg ha-1, 1394 kg ha-1 and 208 kg ha-1 over Vandana under SS, MS and NS respectively (Table 3, Fig. S10). The PHT was reduced by 8 cm under SS. The grain yield improvement of introgression lines possessing qDTY12.1 + qDTY3.1 in Anjali background ranged from 516 to 613 kg ha-1 and from 156 to 540 kg ha-1 over the recurrent parent under SS and MS (Table 3, Fig. S11). The data on the DTF and PHT of the selected promising lines under different level of stresses is presented in Table 3.
Genomic and environmental interactions
Despite being the large effect QTLs reported for the grain yield under drought stress at reproductive stage, the near isogenic lines with one, two, three or multiple QTLs showed grain yield variability in each of the genetic background. The contrasting performance of near isogenic lines possessing similar qDTY and also qDTY combinations may be due to the epistatic interactions of the introgressed QTLs among themselves and with the background marker loci under variable stresses level. The digenic interactions between marker loci and with the background were detected in low yielding nearly isogenic lines in IR64-Sub1, TDK1-Sub1, Savitri, MR219 and Samba Mahsuri backgrounds17,22. An epistatic interaction of qDTY12.1 with qDTY3.2 and qDTY2.3 in Vandana/Way Rarem background which was significantly improving yield of qDTY12.1 introgressed lines60. An epistatic interaction of qDTY4.1 on chromosome 4 and qDTY9.1 locus chromosome 9 with qDTY7.1 chromosome 7 in Samba Mahsuri background17, qDTY2.2 and qDTY3.1 with qDTY12.1 in MR219 background showed significant yield improvement over the lines having single QTLs14. The qDTY1.1 showed positive interactions with qDTY2.1, qDTY2.2, and qDTY3.1 across different genetic backgrounds used in the present study. Further, the positive interaction of qDTY2.2 with qDTY4.1, qDTY3.1 and qDTY12.1, qDTY3.1 with qDTY1.1, qDTY2.2, qDTY12.1, qDTY6.1, and qDTY6.2 also showed yield improvement over the lines with other qDTYs combinations in at least the genetic background used in the present study. Further, the release of different introgression lines with same or different QTL combinations indicated the role of QTL x environment interactions.
Marker-assisted breeding product development
A total of 38 improved introgression lines in Swarna-Sub1 background, 6 in IR64-Sub1 background, 27 in Samba Mahsuri background, 6 in TDK1-Sub1 background, 20 in MR219 background, 7 in Savitri background, 51 in MTU1010 background, 7 in Swarna background, 3 in Sahbhagi dhan background were selected for lowland conditions. Further, a total 6 improved introgression lines in Vandana background, 10 in Anjali background and 25 in Kalinga III background were selected for upland conditions.
The developed near isogenic lines in the background of mega varieties not only have multiple QTLs but possess high grain and cooking quality traits, to the extent that some of the near isogenic lines have been released as varieties in different countries. The dual drought-submergence tolerance introgression lines in Swarna-Sub1 background, IR 96322-34-223-B-1–1-1–1 possessing qDTY1.1 + qDTY2.1 + qDTY3.1 + Sub1 was released as variety under name CR dhan 801 in India in 2017; IR 96321-1447-651-B-1–1-2 possessing qDTY1.1 + qDTY3.1 + Sub1 and IR 94391-131-358-19-B-1-1-1 possessing qDTY3.1 + Sub1 were released as varieties in Nepal in 2017 under names Bahuguni dhan-1′ and ‘Bahuguni dhan-2′ respectively. The drought tolerant introgression line in IR64 background; IR 87707-445-B-B-B has been released in India for rainfed lowland regions as DRR Dhan 42; IR 87707-446-B-B-B in Nepal as Sukha dhan 4 in 2014 and in Myanmar as Yaenelo 4 in 2015. In addition, IR 87705-44-4-B-B, IR 87707-182-B-B-B and IR 87705-83-12-B-B have been released under names Yaenelo 5, Yaenelo 6 and Yaenelo 7, respectively in Myanmar in 2016. The release of marker-assisted breeding product in different countries prove the reasoning that QTLs for abiotic stress can be combined with high grain yield and good grain and cooking quality and also that the undesirable linkages reported earlier have been successfully broken to remove the linkage drag.
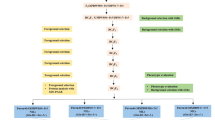
The summarized information on donors, QTLs used in marker-assisted introgression program, recipients, markers used for foreground and recombinant selection of introgressed region, size of introgressed segments and details on the number of total lines screened across different seasons/years and conditions and number of selected promising introgression lines in each background are presented in Fig. 1.

The summarized information on donors, QTLs used in marker-assisted introgression program, recipients, markers used for foreground and recombinant selection of introgressed region, size of introgressed segments and details on the number of total lines screened across different seasons/years and conditions and number of selected promising introgression lines in each background.
Discussion
Development of introgression lines have been extensively employed in the identification and validation of the useful effects of genomic regions from the donor parents. Majority of the QTLs that have been detected so far in many studies20,61,62,63,64,65,66,67,68,69,70,71,72 and very few of them have been exploited and validated further in breeding programs. Some of the candidate genes that have already been cloned73 could be exploited in future breeding programs.
The genetic backgrounds such as Swarna, Swarna-Sub1, IR64, Samba Mahsuri, Vandana, Anjali, Kalinga III, MTU1010, Savitri, MR219 and TDK1-Sub1 that were used in this study to develop introgression lines are widely cultivated and accepted by the farmer in South and South-East Asian countries such as India, Nepal, Malaysia and Lao PDR. All these have production bottleneck due to their susceptibility towards drought, heat and low temperature stresses. Rice breeding at IRRI has made a significant progress in finding the key genomic regions associated with drought as well as submergence tolerance. Majority of the large-effect grain yield drought QTLs and Sub1 for submergence have shown consistency in performances and explained a large part of phenotypic variances18,23,24,25,29,30.
The loci contributing grain yield improvement under drought stress reported to be collocated with the loci governing early days to flowering, plant height and reduced yield under irrigated conditions. The large segregating populations have been developed in Swarna-Sub1 and IR64 backgrounds to break these linkages and semi-dwarf medium duration lines have been successfully developed74. Apart from this, multilocational evaluations of the promising breeding lines offer opportunities to understand the QTL x environment interactions.
Combining multiple stress tolerances in single background of high yielding breeding line/variety are the breeder’s goals that have been targeted for a long time in the past. This is primarily because, in the years with well-characterised climate, the multiple stress-tolerant varieties should provide yield comparable to the popular high-yielding varieties in that area but shall also make the introgression of multiple lines easy, effective and without any undesirable effect. Rice drought molecular breeding at IRRI has witnessed some of the successful example of marker assisted introgression of drought QTLs in the elite backgrounds14,15,17,18.
The developed multiple stress tolerant introgression lines offer opportunities to the rice community to dissect the genetic variations existing among the introgression lines and to identify potentially useful pre-breeding lines to be serve as donors. In addition, very limited efforts have been performed to identify the QTL × background interactions influencing the overall expression of the target trait. There is a strong need to find such kind of unknown background noises for the effective deployment of the QTLs in marker-assisted breeding program. The developed introgression lines in different genetic backgrounds in the present study would be an important genetic material to predict such genetic interactions which complicates the genotype–phenotype relationship of complex quantitative traits75,76 such as drought, low temperature and high temperature tolerance. The developed introgression lines in different genetic backgrounds with same QTLs combination showed contrasting yield under different level of drought stress. The developed introgression lines in background of TDK1 Sub1, Samba Mahsuri, Savitri, MR219, MTU1010, Vandana, Anjali and Kalinga III can further be used for the genetic dissection of epistatic interactions contributing to the grain yield variability in rice under drought.
The complexity of genetic control of complex traits lead to wide variations in the performance of introgression lines. It has been observed that the same QTL showed variable performance across different backgrounds and environments. For a successful marker-assisted introgression program, it is very important to choose QTLs with large genetic effect and stability across wide range of environmental conditions and drought intensities25,77. Selection of the lines with higher yield based on phenotypic selection provided an opportunity to develop lines with positive interactions.
The marker-assisted breeding programme involving introgression of at least 2–3 major QTLs in different genetic backgrounds has permitted the development of many high-yielding multiple stress-tolerant lines. The introgression of 2–3 major QTLs in the background of widely cultivated popular high-yielding rice varieties lead an economic grain yield improvement of 1.0–1.2 t ha–1 under severe drought stress in farmers’ fields. The yield superiority of multiple stress tolerant lines across backgrounds and over generations clearly indicated that the tolerance of drought, submergence, low temperature and high temperature can be successfully combined even though these traits are controlled by different physiological mechanisms. The introgression lines possessed better grain quality than the traditional donors and so the development of multiple stress tolerant variety now with high quality has never been as easy as now. The development and release of marker-assisted breeding product for the rainfed lowland areas in IR64 and Swarna-Sub178 backgrounds are the successful examples that should instigate the rice breeders to deploy the identified QTLs in the marker-assisted breeding programs targeting grain yield improvement under multiple abiotic stresses.
Conclusions
The near isogenic lines with one, two, three and more QTLs have been developed in -eleven different elite backgrounds by the integration of phenotypic-genotypic selection approach. These near isogenic lines are free from linkage drag and hence we are able to combine increased yield under drought with high yield potential, dwarf plant type and good grain quality. Development of high-yielding and multiple stress tolerant introgression lines in wide range of backgrounds and for two diverse ecosystems validate the effectiveness of QTLs and marker-assisted introgression program. Development of such multiple stress tolerant rice varieties will provide opportunities to use these introgressed lines across different rice ecosystems to reduce yield under abiotic stresses and have an impact on increase in rice production and overall increase in the farm output in stress prone areas of South and South East Asia. The near isogenic lines with combination of QTLs brings a great advantage to breeding by providing opportunities to breeders to introgress QTLs in less time by just attempting one or two crosses as against three to four years that were spend earlier just to bring multiple QTLs together. The study clearly indicated that it is necessary to identify and break such linkage drags for effective use of QTLs in the trait development or breeding programs. The study also validated that once linkage drags are removed, it is possible to combine tolerance to multiple traits controlled by diverse physiological mechanisms—low temperature and high temperature; drought and low temperature, high yield and drought tolerance and low temperature. The findings from the current study can revolutionize the marker assisted breeding as very few of the large number of QTLs identified have been used in the past. The developed introgression lines in different genetic backgrounds are unique set of germplasm to the interaction studies to break the grain yield improvement barrier under multiple stresses. In addition, these lines may serve as an important material to study the functional characterization of candidate genes and genetic networks utilizing advances in functional genomics.
Materials and methods
The present study was conducted at International Rice Research Institute (IRRI), Los Baños, Laguna (Philippines) and IRRI South Asia breeding hub (SAH), Hyderabad, India during years from 2012 to 2017.
Breeding for drought tolerance at IRRI, Philippines
Plant material
The first step of breeding program is to identify the appropriate donor exploiting the genetic variations in the existing germplasm. To avoid undesirable linkages associated with the traditional donors, improved breeding lines with genomic regions associated with different abiotic stresses were used in the present study79. The improved breeding lines used as donors were IR 91659-54-35 (qDTY1.1), IR 77298-14-1-2-10 (qDTY2.2), IR 81896-B-B-195 (qDTY3.1), IR 77298-5-6-18 (qDTY3.2), IR 74371-46-1-1 and IR 84984-83-15-18-B (qDTY12.1), IR 86918-B-315 (qDTY1.1 + qDTY1.2), IR 81896-B-B-195 (qDTY2.1 + qDTY3.1), IR 87728-75-B-B and IR 77298-14-1-2-17 (qDTY2.2 + qDTY4.1), Vandana (qDTY2.3 + qDTY3.2), and IR55419-04 (qDTY3.1 + qDTY6.1 + qDTY6.2). The popular lowland adapted high-yielding varieties such as Swarna, Swarna-Sub1, IR64-Sub1, Samba Mahsuri, TDK1, MR219, Savitri and MTU1010, and the upland adapted rice varieties Kalinga 3, Vandana and Anjali were used as recipient parent to develop marker-assisted derived breeding lines for future climate-change ready varietal development.
Development of introgression lines
The Swarna-Sub1 introgression lines were developed using donors IR 91659-54-35 possessing qDTY1.1; IR 81896-B-B-195 possessing qDTY2.1 and qDTY3.1. The introgression lines in IR64-Sub1 background were developed from crossing involving donors IR 77298-14-1-2-17 possessing two qDTYs (qDTY2.2 and qDTY4.1)18, IR 86918-B-315 possessing two qDTYs (qDTY1.1 and qDTY1.2), Vandana possessing two qDTYs (qDTY2.3 and qDTY3.2) and Way Rarem possessing one qDTY (qDTY12.1). The introgression lines in TDK1-Sub1 background were developed using BC1F3:5 derived pyramided lines from the cross IR55419-04/2*TDK115. Savitri introgression lines were developed involving the pyramided lines derived from IR 77298-5-6-18/2*Savitri//IR 74371-1-1/2*Savitri///2*Savitri carrying qDTY3.2 and qDTY12.1 QTLs in Savitri background15,26,27. The introgression lines in Samba Mahsuri background were developed involving IR 87728-75-B-B, a donor of qDTY2.2 and qDTY4.117. MR219 introgression lines were generated using three donors IR 77298-14-1-2-10 (qDTY2.2), IR 81896-B-B-195 (qDTY3.1) and IR 84984-83-15-18-B (qDTY12.1)14. The introgression lines in MTU1010 background were developed using IR 77298-14-1-2-10 (qDTY2.2) and IR 55419-04 (qDTY3.1, qDTY6.1 and qDTY6.2). The Moroberekan/3*Swarna introgression lines were developed using Moroberekan as donor (qDTY3.2). The introgression lines in Vandana background was developed using IR 79971-B-102-B-B (qDTY12.1) as donor parent, in Anjali background were developed involving donors for qDTY3.1 (IR 81896-B-B-195-2), and qDTY12.1 (IR 84984-83-15-18-B-B-93) and in Kalinga III background using different donors of qDTY12.1 (IR 8498483-15-442, IR 84984-83-15-688-B and IR 84984-83-15-481-B).
Phenotyping screening
The screening of marker-assisted derived breeding lines was conducted under lowland and/or upland reproductive stage drought stress (RS) and irrigated control/non-stress (NS) conditions at IRRI (International Rice Research Institute). The experiments were planted in two to three replications in single or two or four-rows plots with row length of 5 m and 2.0 to 3.0 m in lowland and upland condition, respectively in an alpha lattice design. The lowland experiments involved transplanting that involved the transplantation of 21-days-old seedlings in the field with a single seedling per hill. The long duration introgression lines were planted 15–20 days earlier than the medium duration varieties. The field management activities of the lowland experiments were carried out in similar way as described earlier by Venuprasad et al.24. The detailed information on number of entries tested, generation, experimental design, treatments and number of replications is presented in Table S1.
Upland experiments were established under dry direct seeded conditions by maintaining 20 cm × 20 cm between row to row and plant to plant distance. Field management practises under upland was done as described by Bernier et al.23. In brief, under upland conditions, combination of Oxadiazon (pre-emergence herbicide) at 6 days after seeding (DAS) @ 0.5 kg ai ha-1, Bispyribac sodium (early post emergence) at 11 and 22 DAS @ 0.03 kg ai ha-1 in addition to spot weeding at 35 and 55 DAS were used to control the weeds. IPM (Integrated pest management) practices was followed involving ditrac @ 0.05 g kg-1 (0.005%), brodifacoum as rat bait to control rats, and pre-seeding application of the 0.075 kg ai ha-1 of Fipronil along bunds and the plot edges at 7 DAS.
Lowland drought screening. At 30 days after transplanting (DAT), complete drainage of water was carried out and the drought stress at reproductive stage was initiated by holding irrigation. The initiation of reproductive stage drought stress for the long duration lines were adjusted based on their reproductive cycle. When ~ 75% of the population showed symptoms of severe leaf rolling and the water table depth remained > 100 cm for approximately > 2 weeks; a life-saving irrigation was applied through the flash flooding. After 24 h, complete drainage of the water from the field was carried out again to impose the second cycle of drought stress. The 1.1 m PVC (polyvinyl chloride) pipe was installed in the soil to measure the water table depth at regular intervals in the experimental fields. The depletion of the water table level was measured using a meter scale day-to-day after the onset of the drought stress.
Upland drought screening. The upland experiments were irrigated using sprinkler-irrigation up to 45 DAS, with application of irrigation twice a week during seedling establishment stage and in early vegetative developmental stage. Thereafter, stress at reproductive stage was initiated by holding irrigation. The plots were re-irrigated when the tensiometer at 30 cm soil depth showed reading below – 50 kPa and most of the lines showed wilting and leaf drying. This kind of cyclic stress is effective in drought screening of populations consisting of variable genotypes having broad range of growth duration80,ensuring all breeding lines receive an adequate level of stress during the reproductive stage of development. All the cultural practices were same in upland non-stress and reproductive stage stress experiments, except that in non-stress experiments the irrigation was continued two times a week up to 10 days before the harvest of the crop.
Biotic stress screening. The introgression lines at F2 and F7/F8 stages were also evaluated for the disease resistances such as blast which is caused by Magnaporthe oryzae and bacterial blight which is caused by Xanthomonas oryzae pv. Oryzae. The blast screening was done using the mixed inoculum for the races presents in the Philippines at 10 DAS in the blast nursery. The plants possessing genes viz. pik-s, pi2, pi5(t) and pi9 showed resistance against the inoculum. The inoculation and screening for the bacterial blight race 1, PXO61 and race 2, PXO86 was done at maximum tillering stage in field following Kauffman81. The resistant or susceptibility of introgression lines against the blast and bacterial blight diseases was observed on IRRI, SES scale82.
Phenotypic data collection
The agronomic data on days to 50% flowering (DTF), plant height (PHT), and grain yield (GY) were recorded for all the experiments. DTF was recorded when ~ 50% the plants in a plot showed panicle exertion. At maturity, three random plants per plot were measured from the soil surface to the tip of the highest panicle on main tiller to record the PHT (cm). The plant samples at maturity were harvested, dried to 14% of the moisture before weighing, and weights to record the GY (kg ha–1). The developed introgression lines were screened under both reproductive-stage drought (RS) and irrigated control non-stress (NS) conditions along with the recipient parent, high-yielding check varieties and drought-tolerant donors. The imposed reproductive stage drought stress was classified into severe and moderate drought stress based on the relative reduction in the grain yield compared to non-stress79. The experiments with yield reduction of < 70% were characterize as severe stress (SS) and with yield reduction of 31–70% were categorized as moderate stress (MS). The selected introgression lines were also evaluated under national/state trials in different countries.
IRRI SAH
Breeding for drought, heat and low temperature tolerance
To develop multiple stress tolerance lines possessing tolerance to low temperature, high-temperature and drought stresses, a recombinant inbred lines (RILs) population consisting of 400 RILs involving IR 91648-B-117-B-1-1 (improved breeding lines from Swarna*3/Moroberekan)83 as donor parent and MTU1010 as recipient parent was developed. Pedigree based breeding was employed to develop multiple tolerance lines involving stringent phenotypic selection for the targeted traits. A series of experiments following standard agronomic practices were carried out between 2013 to 2017 at International Rice Research Institute-South Asia Hub (IRRI-SAH), Patancheru which is at 78° 16′ longitude and at 17° 32′ latitude. The experiments were planted in two replications in twelve-rows plots with 3 m row length maintaining 20 (plant to plant) × 15 (row to row) cm spacing under transplanted conditions in randomized complete block design (RCBD).
Phenotypic screening and data collection
The phenotypic screening of breeding lines for the drought stress at reproductive stage was carried under natural conditions as above-mentioned procedure in both dry and wet seasons. The low temperature and high-temperature screening were carried based on the SES (Standard evaluation system for rice)84. Germination vigor and the seedling survival rate were two main criteria that were used for the evaluation of low temperature tolerance in rice at germination stage85. The high-temperature screening at reproductive stage and low temperature tolerance screening at germination stage was carried out under field condition in dry season while the low temperature tolerance screening at seedling stage was conducted under field in the dry season as well as under controlled condition in the wet seasons. To compare the performance of breeding lines under treatment and non-stress (control) conditions, non-stress experiments were carried out for each drought, heat and low temperature tolerance experiments. Three plants per introgression line were chosen for the measurement of all traits. The phenotypic data on DTF, PHT, number of productive tillers (counted manually), flag leaf length (FLL,cm, panicle length (PL,cm, number of filled grains per panicle (FG/P, spikelet fertility (SF; %, 1000-grain weight (1000 GW; g), grain yield (GY; kg ha-1) and grain type was recorded.
Genotyping of the developed introgression lines in different backgrounds
In the marker-assisted introgression program, rice microsatellite (SSR; simple sequence repeats) markers were extensively used. Earlier reported SSR markers associated with the various grain yield qDTYs15,17,18,23,24,25,26,27,28 were used to detect the targeted region in the developed introgression lines in various backgrounds (Table S2).
The fresh-young leaves from the 21 days old seedlings were collected from each introgression line and the respective parents. The genomic DNA was extracted using the modified CTAB protocol86. Polymerase chain reaction (PCR) amplification was performed with each PCR reaction mixture of 15 μl containing DNA (10 ng), PCR buffer (1 ×), dNTPs (100 μM), oligonucleotide primers (100 μM) and Taq polymerase (1 unit). The PCR products were then resolved on high-resolution PAGE (polyacrylamide gel electrophoresis) (CBS scientific, MGV-202-33) ranged from 6–8% (v/v) depending on the SSR markers product size. The separated fragments of DNA were stained with the SYBER Safe™ before visualizing under UV trans-illuminator (AlphaImager™ System). The strong associations of the reported markers with the introgressed regions and the co-dominance nature of SSRs allowed the easy detection of heterozygotes. As many of the identified qDTYs regions that were used in the introgression program were not fine mapped, all SSRs present within the identified genomic regions/QTLs were used in the genotyping of the introgression lines to avoid a loss of the candidate genes associated with abiotic stresses tolerance (drought, low temperature and high temperature) in the region due to the crossover. To reduce the forward genotyping cost and labor, sequential genotyping approach was used. Firstly, the peak maker for each QTL was used to genotype the F2 segregants, then the breeding lines harbouring the respective donor allele at peak marker were further genotyped with the flanking markers, followed by genotyping with all the markers underlying the QTL regions.
Background recovery
To study the background recovery of the selected introgression lines, background genotyping was carried out. A total of ~ 600 SSR markers equally distributed on all the 12 chromosomes (https://www.gramene.org/) were used to study the polymorphism between the parents. The selected polymorphic SSR markers; 156, 111, 101, 89, 80, and 102 SSR markers were used to estimate the background recovery percentage in Swarna, IR64, TDK1-Sub1, Savitri, Samba Mahsuri, MR219 backgrounds, respectively were chosen. Background study has not been conducted yet in Anjali and Kalinga backgrounds. The background recovery percentage (BC%) of the selected introgression lines in different backgrounds were calculated as Vishwakarma et al.87.
where B is the SSR marker loci homozygous for the genetic background; H is the number of markers loci in heterozygous; N is the total number of polymorphic markers that were used for the background genotyping.
Statistical analysis
The agronomic data collected from all the experiments were analysed statistically. The experiment means and SED (standard error of difference) were calculated using the statistical software PBTools v1.4 developed at IRRI. The LSD (Least significant difference) at P = 5% and 1% significance were used to make the comparison of the test entries means and to estimate the significant differences of the traits of interest between the parents and the introgression lines.
The mixed linear model was used to calculate the analysis of variance:
where µ is the overall mean, Gi is the effect of ith genotype, Rj is the effect of jth replicate, BK (Rj) is block effect of the jth replicate and eijk is the error. Genotypes were kept as fixed and the replication and block effects were kept random for estimating entry means.
References
Krasensky, J. & Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 63(4), 1593–1608 (2012).
Bourke, P. M., Voorrips, R. E., Visser, R. G. & Maliepaard, C. Tools for genetic studies in experimental populations of polyploids. Front. Plant Sci. 9, 513–529 (2018).
Garrido-Cardenas, J. A., Mesa-Valle, C. & Manzano-Agugliaro, F. Trends in plant research using molecular markers. Planta 247(3), 543–557 (2018).
Jena, K. K. & Mackill, D. J. Molecular markers and their use in marker-assisted selection in rice. Crop Sci. 48(4), 1266–1276 (2008).
Lejeune-Hénaut, I. et al. The flowering locus Hr colocalizes with a major QTL affecting winter frost tolerance in Pisum Sativum L. Theor. App. Genet. 116(8), 1105–1116 (2008).
Liu, C. et al. Genetic architecture of the maize kernel row number revealed by combining QTL mapping using a high-density genetic map and bulked segregant RNA sequencing. BMC Genom. 17(1), 915 (2016).
Li, B. et al. High-resolution mapping of QTL for fatty acid composition in soybean using specific-locus amplified fragment sequencing. Theor. App. Genet. 130(7), 1467–1479 (2017).
Li, R. et al. Identification of a locus conferring dominant resistance to maize rough dwarf disease in maize. Sci. Rep. 8(1), 3248 (2018).
Kumar, A. et al. Marker-assisted selection strategy to pyramid two or more QTLs for quantitative trait-grain yield under drought. Rice 11(35), 16 (2018).
Mackill, D. J. & Bouman, J. M. Inheritance of blast resistance in near-isogenic lines of rice. Phytopathology 82(7), 746–749 (1992).
Telebanco-Yanoria, M. et al. A set of near-isogenic lines of Indica-type rice variety CO 39 as differential varieties for blast resistance. Mol. Breed. 27(3), 357–373 (2011).
Khanna, A. et al. Marker assisted pyramiding of major blast resistance genes Pi9 and Pita in the genetic background of an elite Basmati rice variety, Pusa Basmati 1. Indian J. Genet. 75(4), 417–425 (2015).
Xiao, W. M. et al. Pyramiding of Pi46 and Pita to improve blast resistance and to evaluate the resistance effect of the two R genes. J. Int. Agric. 15(10), 2290–2298 (2016).
Shamsudin, N. A. A. et al. Marker assisted pyramiding of drought yield QTLs into a popular Malaysian rice cultivar, MR219. BMC Genet. 17(1), 30 (2016).
Dixit, S. et al. Combining drought and submergence tolerance in rice: marker-assisted breeding and QTL combination effects. Mol. Breed. 37(12), 143 (2017).
Balachiranjeevi, C. H. et al. Marker-assisted pyramiding of two major, broad-spectrum bacterial blight resistance genes, Xa21 and Xa33 into an elite maintainer line of rice, DRR17B. PLoS ONE 13(10), e0201271 (2018).
Sandhu, N. et al. Positive interactions of major-effect QTLs with genetic background that enhances rice yield under drought. Sci. Rep. 8(1), 1626 (2018).
Swamy, B. M. et al. Genetic, physiological, and gene expression analyses reveal that multiple QTL enhance yield of rice mega-variety IR64 under drought. PLoS ONE 8(5), e62795 (2013).
Wijerathna, Y. M. Marker assisted selection: biotechnology tool for rice molecular breeding. Adv. Crop Sci. Technol. 3, 1–4 (2015).
Mei, H. W. et al. Gene actions of QTLs affecting several agronomic traits resolved in a recombinant inbred rice population and two backcross populations. Theor. App. Genet. 110(4), 649–659 (2005).
Zheng, T. Q. et al. Genetic effects of background-independent loci for grain weight and shape identified using advanced reciprocal introgression lines from Lemont× Teqing in rice. Crop Sci. 51(6), 2525–2534 (2011).
Yadav, S. et al. Epistatic interactions of major effect drought QTLs with genetic background loci determine grain yield of rice under drought stress. Sci. Rep. 9(1), 2616 (2019).
Bernier, J., Kumar, A., Ramaiah, V., Spaner, D. & Atlin, G. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci. 47(2), 507–516 (2007).
Venuprasad, R. et al. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor. App. Genet. 120(1), 177–190 (2009).
Vikram, P. et al. qDTY1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 12(1), 89 (2011).
Mishra, K. K. et al. qDTY 12.1: a locus with a consistent effect on grain yield under drought in rice. BMC Genet. 14(1), 12 (2013).
Yadaw, R. B. et al. A QTL for high grain yield under lowland drought in the background of popular rice variety Sabitri from Nepal. Field Crops Res. 144, 281–287 (2013).
Sandhu, N. et al. Identification and mapping of stable QTL with main and epistasis effect on rice grain yield under upland drought stress. BMC Genet. 15(1), 63 (2014).
Xu, K. & Mackill, D. J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 2(3), 219–224 (1996).
Xu, K. et al. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 442(7103), 705 (2006).
Neeraja, C. N. et al. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars. Theor. App. Genet. 115(6), 767–776 (2007).
Septiningsih, E. M. et al. Accelerating the development of new submergence tolerant rice varieties: the case of Ciherang-Sub1 and PSB Rc18-Sub1. Euphytica 202(2), 259–268 (2015).
Fujino, K. et al. Molecular identification of a major quantitative trait locus, qLTG3-1, controlling low-temperature germinability in rice. Proc. Nat. Acad. Sci. 105(34), 12623–12628 (2008).
Saito, K. et al. Identification of two closely linked quantitative trait loci for cold tolerance on chromosome 4 of rice and their association with anther length. Theor. App. Genet. 103(6–7), 862–868 (2001).
Andaya, V. & Mackill, D. QTLs conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica × indica cross. Theor. App. Genet. 106(6), 1084–1090 (2003).
Andaya, V. C. & Mackill, D. J. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 54(392), 2579–2585 (2003).
Andaya, V. C. & Tai, T. H. Fine mapping of the qCTS12 locus, a major QTL for seedling cold tolerance in rice. Theor. App. Genet. 113(3), 467–475 (2006).
Zhang, Z. H., Qu, X. S., Wan, S., Chen, L. H. & Zhu, Y. G. Comparison of QTL controlling seedling vigour under different temperature conditions using recombinant inbred lines in rice (Oryza sativa). Ann. Bot. 95(3), 423–429 (2004).
Sato, Y. et al. Heat shock-mediated APX gene expression and protection against chilling injury in rice seedlings. J. Exp. Bot. 52(354), 145–151 (2001).
Takeuchi, Y. et al. Mapping quantitative trait loci controlling cool-temperature tolerance at booting stage in temperate japonica rice. Breed. Sci. 51, 191–197 (2001).
Liu, F. et al. Identification and mapping of quantitative trait loci controlling coldtolerance of Chinese common wild rice (O. rufipogon Griff.) at booting to flowering stages. Chin. Sci. Bull. 48, 2068–2071 (2003).
Dai, L. et al. Identification of quantitative trait loci controlling cold tolerance at the reproductive stage in Yunnan landrace of rice Kunmingxiaobaigu. Breed. Sci. 54(3), 253–258 (2004).
Xu, L. M. et al. Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice near-isogenic line. Plant Sci. 174(3), 340–347 (2008).
Suh, J. P. et al. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerant genotypes of rice (Oryza Sativa L.). Theor. App. Genet. 120(5), 985–995 (2010).
Ye, C. et al. A QTL controlling low temperature induced spikelet sterility at booting stage in rice. Euphytica 176(3), 291–301 (2010).
Verma, S. K., Xalxo, M. S., Saxena, R. R. & Verulkar, S. B. Identification of QTLs for cold tolerance at seedling stage in rice (Oryza sativa L.). Indian J. Genet. Plant Breed. 74(1), 86–89 (2014).
Cao, L. et al. Mapping QTLs for heat tolerance and correlation between heat tolerance and photosynthetic rate in rice. Chin. J. Rice Sci. 17(3), 223–227 (2003).
Chang-lan, Z. et al. Mapping QTL for heat-tolerance at grain filling stage in rice. Rice Sci. 12(1), 33–38 (2005).
Zhang, T. et al. QTL mapping for heat tolerance of the tassel period of rice. Mol. Plant Breed. 6, 867–873 (2008).
Zhang, G. L. et al. Bulked segregant analysis to detect QTL related to heat tolerance in rice (Oryza sativa L.) using SSR markers. Agric. Sci. China 8(4), 482–487 (2009).
Jagadish, S. V. K. et al. Genetic analysis of heat tolerance at anthesis in rice. Crop Sci. 50(5), 1633–1641 (2010).
Xiao, Y. H. et al. Quantitative trait loci associated with pollen fertility under high temperature stress at flowering stage in rice (Oryza sativa). Rice Sci. 18(3), 204–209 (2011).
Ye, C. et al. Mapping QTL for heat tolerance at flowering stage in rice using SNP markers. Plant Breed. 131(1), 33–41 (2012).
Ye, C. et al. Identifying and confirming quantitative trait loci associated with heat tolerance at flowering stage in different rice populations. BMC Genet. 16(1), 41 (2015).
Cheng, L. R. et al. Genetic analysis of cold tolerance at seedling stage and heat tolerance at anthesis in rice (Oryza sativa L.). J. Int. Agric. 11(3), 359–367 (2012).
Poli, Y. et al. Characterization of a Nagina22 rice mutant for heat tolerance and mapping of yield traits. Rice 6(1), 36 (2013).
Hirabayashi, H. et al. qEMF3, a novel QTL for the early-morning flowering trait from wild rice, Oryza officinalis, to mitigate heat stress damage at flowering in rice. O. Sativa. J. Exp. Bot. 66(5), 1227–1236 (2014).
Zhao, L. et al. Mapping quantitative trait loci for heat tolerance at anthesis in rice using chromosomal segment substitution lines. Breed. Sci. 66(3), 358–366 (2016).
Vikram, P. et al. Linkages and interactions analysis of major effect drought grain yield QTLs in rice. PLoS ONE 11(3), e0151532 (2016).
Dixit, S. et al. Increased drought tolerance and wider adaptability of qDTY 12.1 conferred by its interaction with qDTY 2.3 and qDTY 3.2. Mol. Breed. 30(4), 1767–1779 (2012).
Zhuang, J. Y. et al. Analysis of QTL × environment interaction for yield components and plant height in rice. Theor. App. Genet. 95(5–6), 799–808 (1997).
Xiao, J. et al. Identification of trait-improving quantitative trait loci alleles from a wild rice relative Oryza rufipogon. Genetics 150(2), 899–909 (1998).
Yagi, T. et al. QTL mapping of spikelet number in rice (Oryza sativa L.). Breed. Sci. 51(1), 53–56 (2001).
Moncada, P. et al. Quantitative trait loci for yield and yield components in an Oryza sativa× Oryza rufipogon BC2F2 population evaluated in an upland environment. Theor. App. Genet. 102(1), 41–52 (2001).
Brondani, C., Rangel, P., Brondani, R. & Ferreira, M. QTL mapping and introgression of yield-related traits from Oryza glumaepatula to cultivated rice (Oryza sativa) using microsatellite markers. Theor. App. Genet. 104(6–7), 1192–1203 (2002).
Septiningsih, E. M. et al. Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor. App. Genet. 107(8), 1419–1432 (2003).
Hittalmani, S. et al. Identification of QTL for growth-and grain yield-related traits in rice across nine locations of Asia. Theor. App. Genet. 107(4), 679–690 (2003).
Kobayashi, S. et al. Identification and characterization of quantitative trait loci affecting spikelet number per panicle in rice (Oryza sativa L.). Field Crops Res. 89(2–3), 253–262 (2004).
Ashikari, M. et al. Cytokinin oxidase regulates rice grain production. Science 309(5735), 741–745 (2005).
Xing, Y. Z., Tang, W. J., Xue, W. Y., Xu, C. G. & Zhang, Q. Fine mapping of a major quantitative trait loci, qSSP7, controlling the number of spikelets per panicle as a single Mendelian factor in rice. Theor. App. Genet. 116(6), 789–796 (2008).
Ando, T. et al. Genetic dissection and pyramiding of quantitative traits for panicle architecture by using chromosomal segment substitution lines in rice. Theor. App. Genet. 116(6), 881–890 (2008).
Liu, T., Mao, D., Zhang, S., Xu, C. & Xing, Y. Fine mapping SPP1, a QTL controlling the number of spikelets per panicle, to a BAC clone in rice (Oryza Sativa). Theor. App. Genet. 118(8), 1509–1517 (2009).
Miura, K., Ashikari, M. & Matsuoka, M. The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci. 16, 319–326 (2011).
Vikram, P. et al. Drought susceptibility of modern rice varieties: an effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 5, 14799 (2015).
Cooper, M., Podlich, D. W. & Smith, O. S. Gene-to-phenotype models and complex trait genetics. Aust. J. Agr. Sci. 56, 895–918 (2005).
Cooper, M. & Podlich, D. W. The E(NK) model: Extending the NK model to incorporate gene-by-environment interactions and epistasis for diploid genomes. Complexity 7, 31–47 (2002).
Bernier, J. et al. Characterization of the effect of a QTL for drought resistance in rice, qtl 12.1, over a range of environments in the Philippines and eastern India. Euphytica 166(2), 207–217 (2009).
Sandhu, N. et al. Marker assisted breeding to develop multiple stress tolerant varieties for flood and drought prone areas. Rice 12(1), 8 (2019).
Kumar, A. et al. Breeding high-yielding drought-tolerant rice: genetic variations and conventional and molecular approaches. J. Exp. Bot. 65, 6265–6278 (2014).
Lafitte, H. R., Price, A. H. & Courtois, B. Yield response to water deficit in an upland rice mapping population: associations among traits and genetic markers. Theor. App. Genet. 109(6), 1237–1246 (2004).
Kauffman, H. E. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 57, 537–541 (1973).
International Rice Research Institute (IRRI). Standard Evaluation System for Rice (4th edn) International Rice Research Institute, Manila, the Philippines (1996).
Dixit, S. et al. QTLs for tolerance of drought and breeding for tolerance of abiotic and biotic stress: an integrated approach. PLoS ONE 9(10), e109574 (2014).
International Rice Research Institute (IRRI). Standard evaluation system for rice. 2014.
Han, L. Z. et al. Genetic and QTL analysis for low-temperature vigor of germination in rice. Acta. Genet. Sin. 33, 998–1006 (2006).
Murray, M. G. & Thompson, W. F. Rapid isolation of high molecular weight DNA. Nucleic Acids Res. 8, 4221–4235 (1980).
Vishwakarma, M. K. et al. Introgression of the high grain protein gene Gpc-B1 in an elite wheat variety of Indo-Gangetic Plains through marker assisted backcross breeding. Curr. Plant Bio. 1, 60–67 (2014).
Acknowledgements
We thank Paul Maturan and Ma. Teresa Sta. Cruz for the management of lowland and upland field experiments, Evangeline Angeles and Jocelyn Guevarra for assistance with seed preparations, and Ruth Erica Carpio for assistance in agronomic data management. This study was supported by the BMGF (Bill & Melinda Gates Foundation), the GTZ (Deutsche Gesellschaft für Technische Zusammenarbeit) and the ICAR (Indian Council of Agricultural Research). The authors thank BMGF, GTZ and ICAR for providing financial support to the study.
Author information
Authors and Affiliations
Contributions
Conceptualization, A.K; methodology, A.K., N.S.; experiment designing, set-up, data collection, genotyping data analysis, N.S., C.V., R.P., S.Y., R.R.M.; writing and original draft preparation: N.S.; review and editing: A.K., V.K.S. All authors have carefully read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kumar, A., Sandhu, N., Venkateshwarlu, C. et al. Development of introgression lines in high yielding, semi-dwarf genetic backgrounds to enable improvement of modern rice varieties for tolerance to multiple abiotic stresses free from undesirable linkage drag. Sci Rep 10, 13073 (2020). https://doi.org/10.1038/s41598-020-70132-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-70132-9
This article is cited by
-
Superior haplotypes of key drought-responsive genes reveal opportunities for the development of climate-resilient rice varieties
Communications Biology (2024)
-
Traces of Introgression from cAus into Tropical Japonica Observed in African Upland Rice Varieties
Rice (2023)
-
Genetic Trends Estimation in IRRIs Rice Drought Breeding Program and Identification of High Yielding Drought-Tolerant Lines
Rice (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
