
Abstract
Food intake follows a predictable daily pattern and synchronizes metabolic rhythms. Neurons expressing agouti-related protein (AgRP) read out physiological energetic state and elicit feeding, but the regulation of these neurons across daily timescales is poorly understood. Using a combination of neuron dynamics measurements and timed optogenetic activation in mice, we show that daily AgRP-neuron activity was not fully consistent with existing models of homeostatic regulation. Instead of operating as a ‘deprivation counter’, AgRP-neuron activity primarily followed the circadian rest–activity cycle through a process that required an intact suprachiasmatic nucleus and synchronization by light. Imposing novel feeding patterns through time-restricted food access or periodic AgRP-neuron stimulation was sufficient to resynchronize the daily AgRP-neuron activity rhythm and drive anticipatory-like behavior through a process that required DMHPDYN neurons. These results indicate that AgRP neurons integrate time-of-day information of past feeding experience with current metabolic needs to predict circadian feeding time.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
Data availability
Raw data are available from the corresponding author upon reasonable request. Source data are provided with this paper.
References
Mistlberger, R. E. Food as circadian time cue for appetitive behavior. F1000Res. 9, F1000 Faculty Rev-61 (2020).
Gooley, J. J., Schomer, A. & Saper, C. B. The dorsomedial hypothalamic nucleus is critical for the expression of food-entrainable circadian rhythms. Nat. Neurosci. 9, 398–407 (2006).
Crosby, P. et al. Insulin/IGF-1 drives PERIOD synthesis to entrain circadian rhythms with feeding time. Cell 177, 896–909.e20 (2019).
Landgraf, D. et al. Oxyntomodulin regulates resetting of the liver circadian clock by food. eLife 4, e06253 (2015).
Balsalobre, A. et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 289, 2344–2347 (2000).
LeSauter, J., Hoque, N., Weintraub, M., Pfaff, D. W. & Silver, R. Stomach ghrelin-secreting cells as food-entrainable circadian clocks. Proc. Natl Acad. Sci. USA 106, 13582–13587 (2009).
Gallardo, C. M. et al. Dopamine receptor 1 neurons in the dorsal striatum regulate food anticipatory circadian activity rhythms in mice. eLife 3, e03781 (2014).
Deem, J. D., Faber, C. L. & Morton, G. J. AgRP neurons: regulators of feeding, energy expenditure, and behavior. FEBS J. 289, 2362–2381 (2021).
Chen, Y., Lin, Y.-C., Kuo, T.-W. & Knight, Z. A. Sensory detection of food rapidly modulates arcuate feeding circuits. Cell 160, 829–841 (2015).
Podyma, B. et al. The p75 neurotrophin receptor in AgRP neurons is necessary for homeostatic feeding and food anticipation. eLife 9, e52623 (2020).
Tan, K., Knight, Z. A. & Friedman, J. M. Ablation of AgRP neurons impairs adaption to restricted feeding. Mol. Metab. 3, 694–704 (2014).
Bugarith, K., Dinh, T. T., Li, A.-J., Speth, R. C. & Ritter, S. Basomedial hypothalamic injections of neuropeptide Y conjugated to saporin selectively disrupt hypothalamic controls of food intake. Endocrinology 146, 1179–1191 (2005).
Reichenbach, A. et al. AgRP neurons require carnitine acetyltransferase to regulate metabolic flexibility and peripheral nutrient partitioning. Cell Rep. 22, 1745–1759 (2018).
Cedernaes, J. et al. Transcriptional basis for rhythmic control of hunger and metabolism within the AgRP neuron. Cell Metab. 29, 1078–1091 (2019).
Henry, F. E., Sugino, K., Tozer, A., Branco, T. & Sternson, S. M. Cell type-specific transcriptomics of hypothalamic energy-sensing neuron responses to weight-loss. eLife 4, e09800 (2015).
Mandelblat-Cerf, Y. et al. Arcuate hypothalamic AgRP and putative POMC neurons show opposite changes in spiking across multiple timescales. eLife 4, e07122 (2015).
Betley, J. N. et al. Neurons for hunger and thirst transmit a negative-valence teaching signal. Nature 521, 180–185 (2015).
Su, Z., Alhadeff, A. L. & Betley, J. N. Nutritive, post-ingestive signals are the primary regulators of AgRP neuron activity. Cell Rep. 21, 2724–2736 (2017).
Atasoy, D., Aponte, Y., Su, H. H. & Sternson, S. M. A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. J. Neurosci. 28, 7025–7030 (2008).
Savontaus, E., Conwell, I. M. & Wardlaw, S. L. Effects of adrenalectomy on AGRP, POMC, NPY and CART gene expression in the basal hypothalamus of fed and fasted rats. Brain Res. 958, 130–138 (2002).
Lu, X.-Y. et al. Diurnal rhythm of agouti-related protein and its relation to corticosterone and food intake. Endocrinology 143, 3905–3915 (2002).
Gyengesi, E. et al. Corticosterone regulates synaptic input organization of POMC and NPY/AgRP neurons in adult mice. Endocrinology 151, 5395–5402 (2010).
Perry, R. J. et al. Leptin’s hunger-suppressing effects are mediated by the hypothalamic-pituitary-adrenocortical axis in rodents. Proc. Natl Acad. Sci. USA 116, 13670–13679 (2019).
Geerling, J. C., Engeland, W. C., Kawata, M. & Loewy, A. D. Aldosterone target neurons in the nucleus tractus solitarius drive sodium appetite. J. Neurosci. 26, 411–417 (2006).
Broberger, C., Johansen, J., Johansson, C., Schalling, M. & Hokfelt, T. The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice. Proc. Natl Acad. Sci. USA 95, 15043–15048 (1998).
Krashes, M. J. et al. An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 507, 238–242 (2014).
Garfield, A. S. et al. Dynamic GABAergic afferent modulation of AgRP neurons. Nat. Neurosci. 19, 1628–1635 (2016).
Kim, E. R. et al. Paraventricular hypothalamus mediates diurnal rhythm of metabolism. Nat. Commun. 11, 3794 (2020).
Stephan, F. K. & Becker, G. Entrainment of anticipatory activity to various durations of food access. Physiol. Behav. 46, 731–741 (1989).
Honma, K. I., Honma, S. & Hiroshige, T. Feeding-associated corticosterone peak in rats under various feeding cycles. Am. J. Physiol. 246, R721–R726 (1984).
Fuller, P. M., Lu, J. & Saper, C. B. Differential rescue of light- and food-entrainable circadian rhythms. Science 320, 1074–1077 (2008).
Angeles-Castellanos, M., Aguilar-Roblero, R. & Escobar, C. c-Fos expression in hypothalamic nuclei of food-entrained rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 286, R158–R165 (2004).
Mieda, M., Williams, S. C., Richardson, J. A., Tanaka, K. & Yanagisawa, M. The dorsomedial hypothalamic nucleus as a putative food-entrainable circadian pacemaker. Proc. Natl Acad. Sci. USA 103, 12150–12155 (2006).
Atasoy, D., Betley, J. N., Su, H. H. & Sternson, S. M. Deconstruction of a neural circuit for hunger. Nature 488, 172–177 (2012).
Aponte, Y., Atasoy, D. & Sternson, S. M. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat. Neurosci. 14, 351–355 (2011).
Krashes, M. J. et al. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Invest. 121, 1424–1428 (2011).
Goldstein, N. et al. Hypothalamic neurons that regulate feeding can influence sleep/wake states based on homeostatic need. Curr. Biol. 28, 3736–3747.e3 (2018).
Buijs, F. N. et al. Suprachiasmatic nucleus interaction with the arcuate nucleus; essential for organizing physiological rhythms. eNeuro 4, ENEURO.0028-17.2017 (2017).
Mendez-Hernandez, R., Escobar, C. & Buijs, R. M. Suprachiasmatic nucleus-arcuate nucleus axis: interaction between time and metabolism essential for health. Obesity 28, S10–S17 (2020).
Ohta, H., Yamazaki, S. & McMahon, D. G. Constant light desynchronizes mammalian clock neurons. Nat. Neurosci. 8, 267–269 (2005).
Chou, T. C. et al. Critical role of dorsomedial hypothalamic nucleus in a wide range of behavioral circadian rhythms. J. Neurosci. 23, 10691–10702 (2003).
Kalsbeek, A. et al. GABA receptors in the region of the dorsomedial hypothalamus of rats are implicated in the control of melatonin and corticosterone release. Neuroendocrinology 63, 69–78 (1996).
Tso, C. F. et al. Astrocytes regulate daily rhythms in the suprachiasmatic nucleus and behavior. Curr. Biol. 27, 1055–1061 (2017).
Damiola, F. et al. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 14, 2950–2961 (2000).
Stephan, F. K., Swann, J. M. & Sisk, C. L. Anticipation of 24-hr feeding schedules in rats with lesions of the suprachiasmatic nucleus. Behav. Neural Biol. 25, 346–363 (1979).
Patton, D. F. & Mistlberger, R. E. Circadian adaptations to meal timing: neuroendocrine mechanisms. Front. Neurosci. 7, 185 (2013).
Faber, C. L. et al. Leptin receptor neurons in the dorsomedial hypothalamus regulate diurnal patterns of feeding, locomotion, and metabolism. eLife 10, e63671 (2021).
Tang, Q. et al. Leptin receptor neurons in the dorsomedial hypothalamus input to the circadian feeding network. Sci. Adv. 9, eadh9570 (2023).
Luby, M. D. et al. Food anticipatory activity behavior of mice across a wide range of circadian and non-circadian intervals. PLoS ONE 7, e37992 (2012).
Reichenbach, A. et al. Carnitine acetyltransferase (Crat) in hunger-sensing AgRP neurons permits adaptation to calorie restriction. FASEB J. 32, fj201800634R (2018).
Reed, F., Lockie, S. H., Reichenbach, A., Foldi, C. J. & Andrews, Z. B. Appetite to learn: an allostatic role for AgRP neurons in the maintenance of energy balance. Curr. Opin. Endocr. Metab. Res. 24, 100337 (2022).
Sterling, P. Allostasis: a model of predictive regulation. Physiol. Behav. 106, 5–15 (2012).
Ramsay, D. S. & Woods, S. C. Clarifying the roles of homeostasis and allostasis in physiological regulation. Psychol. Rev. 121, 225–247 (2014).
Zhang, Q. et al. Food-induced dopamine signaling in AgRP neurons promotes feeding. Cell Rep. 41, 111718 (2022).
Grove, J. C. R. et al. Dopamine subsystems that track internal states. Nature 608, 374–380 (2022).
Reichenbach, A. et al. Metabolic sensing in AgRP neurons integrates homeostatic state with dopamine signalling in the striatum. eLife 11, e72668 (2022).
He, Z. et al. Cellular and synaptic reorganization of arcuate NPY/AgRP and POMC neurons after exercise. Mol. Metab. 18, 107–119 (2018).
Hatori, M. et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 15, 848–860 (2012).
Chaix, A., Zarrinpar, A., Miu, P. & Panda, S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 20, 991–1005 (2014).
Acosta-Rodriguez, V. A., Rijo-Ferreira, F., Green, C. B. & Takahashi, J. S. Importance of circadian timing for aging and longevity. Nat. Commun. 12, 2862 (2021).
Matikainen-Ankney, B. A. et al. An open-source device for measuring food intake and operant behavior in rodent home-cages. eLife 10, e66173 (2021).
Aklan, I. et al. NTS catecholamine neurons mediate hypoglycemic hunger via medial hypothalamic feeding pathways. Cell Metab. 31, 313–326.e5 (2020).
Jones, J. R., Simon, T., Lones, L. & Herzog, E. D. SCN VIP neurons are essential for normal light-mediated resetting of the circadian system. J. Neurosci. 38, 7986–7995 (2018).
Jones, J. R., Chaturvedi, S., Granados-Fuentes, D. & Herzog, E. D. Circadian neurons in the paraventricular nucleus entrain and sustain daily rhythms in glucocorticoids. Nat. Commun. 12, 5763 (2021).
Berens, P. CircStat: a MATLAB toolbox for circular statistics. J. Stat. Softw. 31, 1–21 (2009).
Hughes, M. E., Hogenesch, J. B. & Kornacker, K. JTK_CYCLE: an efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythms 25, 372–380 (2010).
Acknowledgements
This work is supported by National Institutes of Health grant R01DK126740 to D.A. We thank E. Herzog, S. Sternson and K. Rahmouni for feedback on the paper and J. Resch for providing the 11βHSD2 virus.
Author information
Authors and Affiliations
Contributions
N.S.-A. performed fiber photometry recording experiments and genotyping, prepared MATLAB and Arduino codes for data acquisition, analyzed the data and prepared the figures. I.A. and N.S.-A. performed surgeries. N.S-.A. and I.A. performed optoentrainment experiments. Y.Y., C.L. and I.A. performed electrophysiological recordings. H.K. and J.R. performed imaging. H.K., J.R., I.A. and N.S.-A. contributed to post hoc analysis. D.D. managed mouse breeding. M.I.A. and B.Y. contributed to behavioral setup instrumentation. D.A. and N.S.-A. conceived experiments and prepared the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks Zane Andrews and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
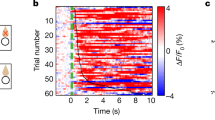
Extended Data Fig. 1 AgRP-neuron activity shifts to anti-phase in the absence of food.
a. Representative concatenated raw 465 and 405 traces recorded 5 minutes every 30 minutes from AgRP-neurons under AL conditions. Dark shade: dark phase, no shade: light phase. b. Average AgRP activity in dark and light phases (n = 14 mice, two-tailed paired t-test, ***p < 0.0001). c. Left: Averaged daily AgRP GFP activity (n = 5 mice). Right: Rayleigh-plot of AgRP:GFP rhythm in AL fed mice, mean peak: ZT17.7, not significant. d. Daily locomotor activity (normalized to total daily activity of each animal) in LD (left panel) and its heatmap showing each animal’s data (right panel). n = 14 mice. e. Two-tailed Pearson correlation between AgRP-neuron activity and locomotor activity across same circadian time points under AL conditions. f. Representative hourly AgRP GCaMP7s traces (z-score) recorded from mice fasted at ZT11, by acquiring for 10 min at every 30 mins in LD (12 h light:12 h dark). Yellow bar: Light phase, dark gray bar: dark phase. g. Averaged AgRP GCaMP7s rhythms recorded from mice fasted at ZT22 (n = 8 mice). Dashed line is from AL. h. Change in AgRP GCaMP activity in dark and light phases in mice fasted at ZT11 or at ZT22 (two-tailed paired t-test, n = 10 animals, **p = 0.00304, *p = 0.0148) i. Scheme describing the protocol in j. j. Left: AgRP GCaMP activity recorded from 24 h fasted mice that were injected satiety-hormone cocktail during day (ZT3), or night (ZT15). Right: Quantification of activity drop by satiety hormones. k. Two-tailed Pearson correlation between AgRP-neuron activity from mice fasted at ZT22 and previous days’ average food intake across the same circadian timepoints. l. Locomotor activity recorded from mice that were fasted at ZT11 (left panel) and heatmap showing each animal’s data (right panel). n = 10 mice. m. Two-tailed Pearson correlation between AgRP-neuron activity (z-score) and locomotor activity across same circadian time points under fasted conditions (fasting started at ZT11). n,o. Same with l,m, but mice were fasted at ZT22. Data are presented as mean ± SEM. For exact P-values and statistics, see Extended Data Fig. 1 Source Data.
Extended Data Fig. 2 Effect of light schedule on AgRP-neuron rhythms.
a. Average locomotor activity (left) and food intake (right) rhythms in the first days of constant dark (DD, left, 6 mice). b. Average locomotor activity, food intake, AgRP GCaMP rhythms (5/6 mice were still rhythmic) under AL conditions, Rayleigh-plots (ad lib and fasted phases differ significantly, p = 0.0329, circular one-way ANOVA), and AgRP activity under fasted conditions, respectively, in later days of DD light setting. c. Average locomotor activity (left) and food intake (middle) rhythms in the first days of constant light (LL, 8 mice) and AgRP GCaMP activity on 10th day of LL setting (right). 0/7 animals were rhythmic. d-f. Two-tailed Pearson correlation between average food intake and locomotor activity across same circadian time points under 12 h Light :12 h Dark (d), DD day 9 (e), and LL day 10 (f). g,h. Average AgRP-neuron activity on baseline day (BL, 7 mice) overlaid with the first jetlag day (JD-1, 6 mice) (g) and quantification of post light phase shift mid-dark phase and mid-light phase average AgRP activities (h) in AL fed mice. i,j. Same as (g,h) except no food was provided (8 BL, 6 JD-1 mice). Data are presented as mean ± SEM. For exact P-values and statistics, see Extended Data Fig. 2 Source Data.
Extended Data Fig. 3 AgRP-neuron specific disruption of cort signaling alters feeding time and meal pattern.
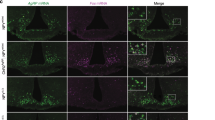
a. Schematic of in vivo fiber photometry recording of AgRP calcium activity in AgRP specific 11βHSD2 expressing (HSD) mice, and representative images showing GCaMP and 11βHSD2 expression in AgRP cells. b,c. Average (b) and individual (heatmap, c) AgRP GCaMP7s activity rhythms recorded from AgRP:HSD mice compared to WT mice in free feeding. d. Rayleigh-plots of AgRP rhythms in HSD animals under AL (mean peak times at ZT4.9, Rayleigh p = 0.0113, Rayleigh test for non-uniformity of circular data) and fasted (mean peak time at ZT23.33, not significant, Rayleigh test for non-uniformity of circular data) conditions. Phases do not differ significantly (one-way circular ANOVA, n = 5 mice each). e. Heatmap showing individual AL locomotor activity in AgRP:HSD mice. f,g. Average (f) and individual (heatmaps, g) daily food intake in AgRP:HSD mice compared to WT mice. Average of 6 days has been shown, n = 5 mice each (two-tailed unpaired t-test, ZT11 *p < 0.05). h,i. Averaged (h) and individual (i) meal size frequencies in wild type and HSD mice (two-tailed unpaired t-test, 6 pellets *p < 0.05). Data are presented as mean ± SEM. For exact P-values, and statistics, see Extended Data Fig. 3 Source Data.
Extended Data Fig. 4 Circadian variations of synaptic input contribute to AgRP-neuron activity rhythms.
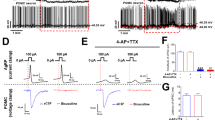
a. Schematic of ex-vivo recording from GFP labeled ARCNPY neurons in Npy-gfp mice and culling times. b,c. Representative loose-seal traces (b, scale: 1 s) and average ARCNPY neuron activity (c) overlaid with average in vivo AgRP:GCaMP7s trace. n = 39-82 neurons for control, and n = 8-19 neurons for data with synaptic blockers. d,e. Representative whole-cell recordings from ARCNPY neurons showing synaptic events (d) and average (e) spontaneous IPSC and EPSC frequency across day overlaid with average in vivo AgRP:GCaMP7s activity. n = 8-17 neurons for sEPSC, and n = 6-22 neurons for sIPSC, two-tailed, unpaired t-test, *p < 0.05, **p < 0.01. Scale: 2 s, 20 pA (sEPSC), 50 pA (sIPSC) f. Schematic of in vivo fiber photometry recording of glutamate dynamics on AgRP-neurons and representative image showing AgRP cells expressing iGluSnFR, and the placement of the fiber in the ARC (Scale 100 μm). g-i. Representative AgRP- iGluSnFR raw trace (g) and averaged signal (h) recorded during access to food in fasted mice and quantification of changes in iGluSnFR signal in the first minutes of food access (two-tailed paired t-test, p = 0.008, n = 3 mice, i). j-l. Summary of changes in glutamate levels on AgRP-neurons over 24 h in AL (average of 3 animals, 3 days, j) and fasted (k) animals, and overlay of two rhythms in averaged in 4 h bins (l, two-tailed paired t-test, *p < 0.05, **p < 0.01). m. Comparison of pre-dark onset (ZT8-ZT12) and dark onset (ZT12-ZT16) levels of glutamate on AgRP-neurons in AL and fasted animals (two-tailed paired t-test, **p = 0.0024, *p = 0.013, n = 3 mice). n-u. Same as f-m, except iGABASnFR was examined (**p = 0.0025, q; **p = 0.0076, *p < 0.05, t; *p = 0.029, u, n = 4 ad lib, n = 5 fasted). Data are presented as mean ± SEM. For exact P-values, number of subjects/neurons, and statistics, see Extended Data Fig. 4 Source Data.
Extended Data Fig. 5 Impairing input from PVNTRH does not affect rhythms for feeding and AgRP activity.
a. Schematic and representative image of in vivo fiber photometry recording of TRH neuronal Ca2+ activity in PVN. ft: Fiber tract. Scale: 300 μm. b,c. Average PVNTRH Ca2+ activity overlaid with AgRP-neuron activity (from Fig. 1) in AL (b) and fasted (c) states. n = 6 AL, n = 7 fasted animals. d. Rayleigh-plot of PVNTRH activity in AL fed (mean peak time ZT14.7, not significant) and fasted (mean peak time at ZT21, not significant) mice. Phases do not differ significantly. e. Two-tailed Pearson correlation between average fasted PVNTRH and average AgRP activities. f. Schematic of in vivo fiber photometry recording from ARCNPY neurons in mice with PVNTRH ablation. Representative images of ARC:GCaMP7s and PVN showing AAV-tdTomato marking the site of caspase injection (upper scale: 300 μm lower scale: 250 μm). g. Average ARCNPY GCaMP activity in animals with PVNTRH ablation (PVNTRH-X) or with intact PVNTRH neurons (Ctrl). Average of 3 days and n = 4 Ctrl and n = 7 PVNTRH-X mice. h. Rayleigh-plot of ARCNPY GCaMP activity in PVNTRH-X mice (mean peak at ZT7) versus control (mean peak time at ZT7.08). Phases do not differ significantly between two groups. i. Comparison of dark and light phase ARCNPY activity in mice with or without PVNTRH ablation, two-tailed, unpaired t-test, n = 4 ctrl, 7 PVNTRH-X mice. j,k. Feeding pattern of PVNTRH-X mice (j, n = 8 Ctrl and 10 PVNTRH-X mice) and total consumed food during daytime and nighttime (k) compared to control animals, two-tailed, unpaired t-test, n = 8 Ctrl and 10 PVNTRH-X mice. Data are presented as mean ± SEM. For exact P-values and statistics, see Extended Data Fig. 5 Source Data.
Extended Data Fig. 6 Rhythmic activity of DMHPDYN is altered by food access.
a. Schematic of in vivo fiber photometry recording from ARCNPY neurons in mice whose DMHPDYN neurons have been transduced with AAV-FLEX-hM3D-mCherry. b. Effect of DMHPDYN neuron activation on ARCNPY calcium response (n = 5 mice). c. Average ARCNPY activity in squared period (b) after saline (SAL) or DCZ injection (two-tailed paired t-test, *p = 0.028, n = 5 mice). d. In vivo DMHPDYN neuron activities in AL and fasted mice (6 mice). e. Difference in average DMHPDYN neuron activity in dark (ZT14-20) and light (ZT2-8) phases in AL and fasted mice (n = 6 mice, two-tailed paired t-test, **p = 0.005). f. Schematic and representative image of caspase ablation of DMHPDYN neurons (DMHPDYN-X). g. Circadian amplitude of ARCNPY/AGRP activity in intact (n = 5) and DMHPDYN-X mice (n = 3, two-tailed unpaired t-test, *p = 0.013). h. Schematic and representative image of in vivo fiber photometry recording from ARCNPY neurons in mice with chronic DMHPDYN activation. Scale: 200 μm i. Average ARCNPY activity in mice that received DCZ in their drinking water versus control mice (Ctrl: no DCZ, average of 2 days, n = 5 mice each). j. Rayleigh-plot of ARCNPY activity (DCZ mean peak: ZT5.4, Rayleigh p = 0.0123, Ctrl mean peak: ZT6.4, Rayleigh p = 0.0417, phases do not differ significantly). k. Average ARCNPY activity for dark and light phases, two-tailed, paired t-test, *p = 0.012, n = 5 mice. l. Circadian amplitude of ARCNPY activity in control and DCZ groups (n = 5 mice each). m,n. Daily feeding pattern (m, DCZ vs Ctrl *p < 0.05, **p = 0.0036, 5-days average, n = 6 mice each) and total food intake (n, two-tailed, paired t-test, * p = 0.017, 5-days average, n = 6 mice each). o-r. Same as i,k,m,n, but with mice without viral DREADD injection (n = 4 mice each). s. ARCNPY activity in DCZ group fasted at ZT11 (n = 5 mice). t. Rayleigh-plots showing peak time in fasted control and DCZ mice (Control mean peak: ZT17.81, DCZ: ZT4.04, phases differ significantly, p = 0.0154, circular one-way ANOVA). u. Average dark and light time ARCNPY activity in fasted control and DCZ groups (n = 5 mice, two-tailed, unpaired t-test, *p < 0.03, **p = 0.007. Data are presented as mean ± SEM. For exact P-values, and statistics, see Extended Data Fig. 6 Source Data.
Extended Data Fig. 7 Time restricted food access synchronizes AgRP-neuron activity rhythms.
a,b. Average daily actogram (a) and average locomotor activity graph on TRF days 23 and no food access day 24 (b) in animals that were given restricted access to food only during daytime (ZT0-12). Orange* denotes fasted days. c. Food intake on day 23 in animals going through daytime TRF. d,e. Nighttime surge in AgRP GCaMP activity dynamics in each mouse shown as heatmap (d) and on average (e) on TRF days 1, 4, 7 and 10 (n = 19, 9, 10, 9 animals, respectively) where animals had access to food between ZT3-7. f. Average locomotor activity in mice under TRF regimen (day 10) compared to AL animals (n = 10 TRF,15 AL mice. Two-tailed unpaired t-test, * p < 0.05, b p < 0.01, a p < 0.001). g. Same as f, but TRF day 11 without food access is compared to fasted animals on otherwise AL schedule (n = 10 mice each). h. Average locomotor activity around food access time at ZT3 in animals on TRF day 10 and day 11 without food access (n = 10 mice, two-tailed paired t-test, ZT3.5 *p = 0.043, ZT4 *p = 0.038). i,j. Two-tailed Pearson correlation of AgRP GCaMP activity (z-score) and locomotor activity in animals on TRF day 10 (i) and day 11 without food access (j). Data are presented as mean ± SEM. for exact P values, see Extended Data 7 Source Data.
Extended Data Fig. 8 Time restricted food access alters AgRP-neuron activity rhythms.
a. Average locomotor activity in mice under TRF regimen (day-17), where animals had access to food between ZT18-22, compared to their AL days (n = 7 animals, two-tailed paired t-test, *p < 0.05). b. Same as a, but TRF day-18 without food access is compared to fasted days while on AL schedule (*p < 0.05, bp < 0.01, ap < 0.001). c,d. Overlaid average locomotor activities during 24-h (c) and around food access time at ZT18 (d) in animals on TRF with and without food access (n = 7 mice, two-tailed paired t-test, *p < 0.05, bp < 0.01, ap < 0.001). e,f. Two-tailed Pearson correlation of AgRP GCaMP activity and locomotor activity in animals on TRF day-17 (e) and day-18 without food access (f). g-i. AgRP (g,h) and ARCNPY (i) neuron activities recorded from 11βHSD2 expressing (g), SCN-ablated (h) and WT control (i) mice subjected to TRF between ZT3-7 for more than 10 days and was provided no food during the recording period. Upper panels: average of n = 5 (g), n = 8 (h) and n = 4 (i) mice, middle panels: individual data from each animal, lower panels: comparison of AgRP/NPY activity in BL-1 (before light phase, ZT21-0), BL-2 (light onset, before food, ZT0-ZT3) and post-food time (Post or P1, ZT4-7), or P2 (ZT7-10, h) periods. One-way ANOVA followed by Tukey multiple comparison test. *p < 0.05, **p < 0.01 j. AgRP-neuron activity recorded from mice that had been subjected to TRF between ZT18-22 for 17 days. Upper panel: average of n = 6 animals, middle panel: individual data from each animal, lower panel: AgRP activity in BL (ZT15-17.5) and post-food time (ZT19-22) periods ***p < 0.0001. Green shades depict food availability. k-o. AgRP (k-m) and ARCNPY (n,o) neuron activities recorded from 11βHSD2 expressing (k), SCN-ablated (l), SCN-ablated, under DD conditions (m), DMHPDYN-ablated (n) and intact control (o) mice that had been subjected to TRF between ZT3-7 for more than 10 days. Upper panels: average of n = 5 (k,n), n = 8 (l,m) and n = 4 (o) mice; middle and lower panels: same as in g-i, ***p < 0.001). Data are presented as mean ± SEM. For exact P-values, see Extended Data 8 Source Data.
Extended Data Fig. 9 DMHPDYN neurons provide timed disinhibition to AgRP-neurons in TRF.
a. Left: Schematic of in vivo fiber photometry recording of PDYN neuronal Ca2+ activity in DMH. Right: Diagram describing TRF setting. b,c. Heatmap showing individual (b) and line graph showing averaged (c) DMHPDYN activity in mice under TRF between ZT3-7. n = 6 mice. d. Average DMHPDYN activity in B1 (ZT21-0), B2(ZT0-3) and Post (ZT4-7) periods in TRF and no food access days. Two-way ANOVA with Tukey’s multiple comparison analysis (No food, B1 vs Post, p = 0.012, B2 vs Post p = 0.016, n = 6 mice). e. Change in DMHPDYN activity (Δ = z-score on TRF day – z-score on no food access day from c) overlaid with ARCNPY/AgRP (from Extended Data Figure 8i). Data are presented as mean ± SEM. For exact P values and data, see Extended Data 9 Source Data.
Extended Data Fig. 10 Allostatic regulation of AgRP-neuron activity by past circadian feeding experience.
a. Proposed model for daily fluctuations in relative AgRP-neuron activity levels. Outside of feeding window, AgRP-neuron activity is relatively unaffected by feeding status (at least on the first day in food deprivation), whereas during feeding time the activity is highly sensitive to food availability. The ‘Adapted feeding window’ is the time of day during which feeding periodically occurred in previous days. In free feeding mice, this window is dark phase and synchronized by light; however, it can be dissociated from circadian phase and reset by restricted food access. b. Schematic depiction of long-term regulation of AgRP-neuron activity by previous day’s circadian feeding experience. Feeding time is encoded into AgRP-neuron activity such that activity rises if anticipated food access does not occur. Top: Under free feeding conditions, AgRP-neuron activity directly or indirectly synchronizes to light through a process that requires intact SCN. Bottom: In time restricted feeding, AgRP activity uncouples from light and synchronizes to feeding time in a process that required intact DMH input.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3 and legends.
Supplementary Code 1
MATLAB (2020b) script for analysis of circadian GCaMP data.
Supplementary Code 2
Arduino script for acquiring locomotor activity.
Supplementary Code 3
MATLAB (2020b) script for extracting FED3 data.
Supplementary Data 1
Source data for Supplementary Fig. 3.
Source data
Source Data Fig. 1
Source data for Fig. 1.
Source Data Fig. 2
Source data for Fig. 2.
Source Data Fig. 3
Source data for Fig. 3.
Source Data Fig. 4
Source data for Fig. 4.
Source Data Fig. 5
Source data for Fig. 5.
Source Data Fig. 6
Source data for Fig. 6.
Source Data Fig. 7
Source data for Fig. 7.
Source Data Extended Data Fig. 1
Source data for Extended Data Fig. 1.
Source Data Extended Data Fig. 2
Source data for Extended Data Fig. 2.
Source Data Extended Data Fig. 3
Source data for Extended Data Fig. 3.
Source Data Extended Data Fig. 4
Source data for Extended Data Fig. 4.
Source Data Extended Data Fig. 5
Source data for Extended Data Fig. 5.
Source Data Extended Data Fig. 6
Source data for Extended Data Fig. 6.
Source Data Extended Data Fig. 7
Source data for Extended Data Fig. 7.
Source Data Extended Data Fig. 8
Source data for Extended Data Fig. 8.
Source Data Extended Data Fig. 9
Source data for Extended Data Fig. 9.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sayar-Atasoy, N., Aklan, I., Yavuz, Y. et al. AgRP neurons encode circadian feeding time. Nat Neurosci 27, 102–115 (2024). https://doi.org/10.1038/s41593-023-01482-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-023-01482-6
This article is cited by
-
Daily feeding entrains hunger-sensing neurons
Nature Neuroscience (2024)
