
Abstract
Mutations affecting the gene encoding the ubiquitin ligase UBE3A cause Angelman syndrome. Although most studies focus on the synaptic function of UBE3A, we show that UBE3A is highly enriched in the nucleus of mouse and human neurons. We found that the two major isoforms of UBE3A exhibit highly distinct nuclear versus cytoplasmic subcellular localization. Both isoforms undergo nuclear import through direct binding to PSMD4 (also known as S5A or RPN10), but the amino terminus of the cytoplasmic isoform prevents nuclear retention. Mice lacking the nuclear UBE3A isoform recapitulate the behavioral and electrophysiological phenotypes of Ube3am–/p+ mice, whereas mice harboring a targeted deletion of the cytosolic isoform are unaffected. Finally, we identified Angelman syndrome-associated UBE3A missense mutations that interfere with either nuclear targeting or nuclear retention of UBE3A. Taken together, our findings elucidate the mechanisms underlying the subcellular localization of UBE3A, and indicate that the nuclear UBE3A isoform is the most critical for the pathophysiology of Angelman syndrome.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
References
LaSalle, J. M., Reiter, L. T. & Chamberlain, S. J. Epigenetic regulation of UBE3A and roles in human neurodevelopmental disorders. Epigenomics 7, 1213–1228 (2015).
Sell, G. L. & Margolis, S. S. From UBE3A to Angelman syndrome: a substrate perspective. Front. Neurosci. 9, 322 (2015).
Scheffner, M. & Kumar, S. Mammalian HECT ubiquitin-protein ligases: biological and pathophysiological aspects. Biochim. Biophys. Acta 1843, 61–74 (2014).
Judson, M. C. et al. GABAergic neuron-specific loss of Ube3a causes Angelman syndrome-like EEG abnormalities and enhances seizure susceptibility. Neuron 90, 56–69 (2016).
Li, W. et al. Angelman syndrome protein Ube3a regulates synaptic growth and endocytosis by inhibiting BMP signaling in Drosophila. PLoS Genet. 12, e1006062 (2016).
Greer, P. L. et al. The Angelman syndrome protein Ube3A regulates synapse development by ubiquitinating Arc. Cell 140, 704–716 (2010).
Kuhnle, S., Mothes, B., Matentzoglu, K. & Scheffner, M. Role of the ubiquitin ligase E6AP/UBE3A in controlling levels of the synaptic protein Arc. Proc. Natl Acad. Sci. USA 110, 8888–8893 (2013).
Pignatelli, M. et al. Changes in mGlu5 receptor-dependent synaptic plasticity and coupling to homer proteins in the hippocampus of Ube3A hemizygous mice modeling angelman syndrome. J. Neurosci. 34, 4558–4566 (2014).
Dindot, S. V., Antalffy, B. A., Bhattacharjee, M. B. & Beaudet, A. L. The Angelman syndrome ubiquitin ligase localizes to the synapse and nucleus, and maternal deficiency results in abnormal dendritic spine morphology. Hum. Mol. Genet. 17, 111–118 (2008).
Gustin, R. M. et al. Tissue-specific variation of Ube3a protein expression in rodents and in a mouse model of Angelman syndrome. Neurobiol. Dis. 39, 283–291 (2010).
Miao, S. et al. The Angelman syndrome protein Ube3a is required for polarized dendrite morphogenesis in pyramidal neurons. J. Neurosci. 33, 327–333 (2013).
Burette, A. C. et al. Subcellular organization of UBE3A in neurons. J. Comp. Neurol. 525, 233–251 (2017).
Burette, A. C. et al. Subcellular organization of UBE3A in human cerebral cortex. Mol. Autism 9, 54 (2018).
Yamamoto, Y., Huibregtse, J. M. & Howley, P. M. The human E6-AP gene (UBE3A) encodes three potential protein isoforms generated by differential splicing. Genomics 41, 263–266 (1997).
Valluy, J. et al. A coding-independent function of an alternative Ube3a transcript during neuronal development. Nat. Neurosci. 18, 666–673 (2015).
Wang, R. & Brattain, M. G. The maximal size of protein to diffuse through the nuclear pore is larger than 60 kDa. FEBS Lett. 581, 3164–3170 (2007).
Ronchi, V. P., Klein, J. M., Edwards, D. J. & Haas, A. L. The active form of E6-associated protein (E6AP)/UBE3A ubiquitin ligase is an oligomer. J. Biol. Chem. 289, 1033–1048 (2014).
Lemak, A., Yee, A., Bezsonova, I., Dhe-Paganon, S. & Arrowsmith, C. H. Zn-binding AZUL domain of human ubiquitin protein ligase Ube3A. J. Biomol. NMR 51, 185–190 (2011).
Seibel, N. M., Eljouni, J., Nalaskowski, M. M. & Hampe, W. Nuclear localization of enhanced green fluorescent protein homomultimers. Anal. Biochem. 368, 95–99 (2007).
Kudo, N. et al. Leptomycin B inactivates CRM1/exportin 1 by covalent modification at a cysteine residue in the central conserved region. Proc. Natl Acad. Sci. USA 96, 9112–9117 (1999).
Ronchi, V. P., Klein, J. M. & Haas, A. L. E6AP/UBE3A ubiquitin ligase harbors two E2~ubiquitin binding sites. J. Biol. Chem. 288, 10349–10360 (2013).
Verma, R., Oania, R., Graumann, J. & Deshaies, R. J. Multiubiquitin chain receptors define a layer of substrate selectivity in the ubiquitin–proteasome system. Cell 118, 99–110 (2004).
Sakata, E. et al. Localization of the proteasomal ubiquitin receptors Rpn10 and Rpn13 by electron cryomicroscopy. Proc. Natl Acad. Sci. USA 109, 1479–1484 (2012).
Young, P., Deveraux, Q., Beal, R. E., Pickart, C. M. & Rechsteiner, M. Characterization of two polyubiquitin binding sites in the 26S protease subunit 5a. J. Biol. Chem. 273, 5461–5467 (1998).
Soukupova, M., Sprenger, C., Gorgas, K., Kunau, W. H. & Dodt, G. Identification and characterization of the human peroxin PEX3. Eur. J. Cell Biol. 78, 357–374 (1999).
Yi, J. J. et al. An Autism-linked mutation disables phosphorylation control of UBE3A. Cell 162, 795–807 (2015).
Mueller, O. T. & Coovadia, A. Gene symbol: UBE3A. Disease: Angelman syndrome. Hum. Genet. 123, 538 (2008).
Matsuura, T. et al. De novo truncating mutations in E6-AP ubiquitin-protein ligase gene (UBE3A) in Angelman syndrome. Nat. Genet. 15, 74–77 (1997).
Silva-Santos, S. et al. Ube3a reinstatement identifies distinct developmental windows in a murine Angelman syndrome model. J. Clin. Invest. 125, 2069–2076 (2015).
Sonzogni, M. et al. A behavioral test battery for mouse models of Angelman syndrome: a powerful tool for testing drugs and novel Ube3a mutants. Mol. Autism 9, 47 (2018).
Rotaru, D. C., van Woerden, G. M., Wallaard, I. & Elgersma, Y. Adult Ube3a gene reinstatement restores the electrophysiological deficits of prefrontal cortex layer 5 neurons in a mouse model of Angelman syndrome. J. Neurosci. 38, 8011–8030 (2018).
Sailer, C. et al. Structural dynamics of the E6AP/UBE3A–E6–p53 enzyme–substrate complex. Nat. Commun. 9, 4441 (2018).
Lee, S. Y. et al. Ube3a, the E3 ubiquitin ligase causing Angelman syndrome and linked to autism, regulates protein homeostasis through the proteasomal shuttle Rpn10. Cell. Mol. Life Sci. 71, 2747–2758 (2013).
Kosugi, S., Hasebe, M., Tomita, M. & Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl Acad. Sci. USA 106, 10171–10176 (2009).
Kawahara, H. et al. Developmentally regulated, alternative splicing of the Rpn10 gene generates multiple forms of 26S proteasomes. EMBO J. 19, 4144–4153 (2000).
Besche, H. C., Haas, W., Gygi, S. P. & Goldberg, A. L. Isolation of mammalian 26S proteasomes and p97/VCP complexes using the ubiquitin-like domain from HHR23B reveals novel proteasome-associated proteins. Biochemistry 48, 2538–2549 (2009).
Wang, X. et al. Mass spectrometric characterization of the affinity-purified human 26S proteasome complex. Biochemistry 46, 3553–3565 (2007).
Scanlon, T. C. et al. Isolation of human proteasomes and putative proteasome-interacting proteins using a novel affinity chromatography method. Exp. Cell Res. 315, 176–189 (2009).
Tai, H. C., Besche, H., Goldberg, A. L. & Schuman, E. M. Characterization of the brain 26S proteasome and its interacting proteins. Front. Mol. Neurosci. 3, 12 (2010).
Martinez-Noel, G. et al. Identification and proteomic analysis of distinct UBE3A/E6AP protein complexes. Mol. Cell. Biol. 32, 3095–3106 (2012).
Pack, C.-G. et al. Quantitative live-cell imaging reveals spatio-temporal dynamics and cytoplasmic assembly of the 26S proteasome. Nat. Commun. 5, 3396 (2014).
Kühnle, S. et al. Angelman syndrome-associated point mutations in the Zn2+-binding N-terminal (AZUL) domain of UBE3A ubiquitin ligase inhibit binding to the proteasome. J. Biol. Chem. 293, 18387–18399 (2018).
Jantz, D., Amann, B. T., Gatto, G. J. & Berg, J. M. The design of functional DNA-binding proteins based on zinc finger domains. Chem. Rev. 104, 789–799 (2004).
Ramamoorthy, S. & Nawaz, Z. E6-associated protein (E6-AP) is a dual function coactivator of steroid hormone receptors. Nucl. Recept. Sign. 6, e006 (2008).
Reid, G. et al. Cyclic, proteasome-mediated turnover of unliganded and liganded ERalpha on responsive promoters is an integral feature of estrogen signaling. Mol. Cell 11, 695–707 (2003).
Khan, O. Y. et al. Multifunction steroid receptor coactivator, E6-associated protein, is involved in development of the prostate gland. Mol. Endocrinol. 20, 544–559 (2006).
Sadhwani, A. et al. Two Angelman families with unusually advanced neurodevelopment carry a start codon variant in the most highly expressed UBE3A isoform. Am. J. Med. Genet. A 176, 1641–1647 (2018).
Nawaz, Z. et al. The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nuclear hormone receptor superfamily. Mol. Cell. Biol. 19, 1182–1189 (1999).
Krishnan, V. et al. Autism gene Ube3a and seizures impair sociability by repressing VTA Cbln1. Nature 543, 507–512 (2017).
Hokayem, El,J. & Nawaz, Z. E6AP in the brain: one protein, dual function, multiple diseases. Mol. Neurobiol. 49, 827–839 (2013).
Loregger, A. et al. Haploid mammalian genetic screen identifies UBXD8 as a key determinant of HMGCR degradation and cholesterol biosynthesis. Arterioscler. Thromb. Vasc. Biol. 37, 2064–2074 (2017).
Warlich, E. et al. Lentiviral vector design and imaging approaches to visualize the early stages of cellular reprogramming. Mol. Ther. 19, 782–789 (2011).
Gunhanlar, N. et al. A simplified protocol for differentiation of electrophysiologically mature neuronal networks from human induced pluripotent stem cells. Mol. Psych. 23, 1336–1344 (2018).
Verschueren, E. et al. Evolution of the SH3 domain specificity landscape in yeasts. PLoS One 10, e0129229 (2015).
Huibregtse, J. M., Scheffner, M., Beaudenon, S. & Howley, P. M. A family of proteins structurally and functionally related to the E6-AP ubiquitin-protein ligase. Proc. Natl Acad. Sci. USA 92, 5249 (1995).
Nelson, J. K. et al. The deubiquitylase USP2 regulates the LDLR pathway by counteracting the E3-ubiquitin ligase IDOL. Circ. Res. 118, 410–419 (2016).
Amin, N. et al. A rare missense variant in RCL1 segregates with depression in extended families. Mol. Psych. 23, 1120–1126 (2018).
Ran, F. A. et al. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 8, 2281–2308 (2013).
Acknowledgements
This work was funded by the following grants: a joint NWO-ZonMw TOP grant (no. 91216045) to Y.E., B.D. and S.A.K. (to study the mechanism of nuclear UBE3A targeting); grants from the Angelman Syndrome Foundation to Y.E. (to generate and characterize isoform-specific Ube3a mice) and B.D. (to identify novel targets and activators of UBE3A); an Erasmus MC grant to Y.E. and S.A.K. (to study UBE3A localization in IPS cells); a grant from the Angelman Syndrome Alliance to B.D., Y.E. and G.M.v.W. (to characterize UBE3A-interacting proteins); and a fellowship from the Associazione Angelman and FROM to M.S. The authors are grateful to the following individuals and groups: M. Elgersma, C. de Koning and M. A. Jolfaei for colony management and genotyping; the Erasmus MC iPS Core Facility for reprogramming and characterization of human iPSC lines; L. Huang for providing the HEK293RPN11-HTBH cells; C. Hoogenraad for providing the mRFP-PEX3 constructs; Y. van der Weegen and E. Hanemaaijer for construction of the plasmids; N. Zelcer for providing material and technical support; and B. Philpot, M. Judson and M. Elgersma for critical reading of the manuscript and valuable advice.
Author information
Authors and Affiliations
Contributions
Y.E., B.D., R.A.T., E.J.M., G.M.v.W. and S.A.K. conceptualized the study. Y.E., B.D., R.A.T., M.S., S.N.V.B., G.M.v.W. and E.J.M. designed the methodology. R.A.T. cultured the mouse neurons and performed confocal imaging and western blotting. M.S. carried out the mouse behavior experiments, statistical analyses and contributed to the confocal imaging and western blot experiments. S.N.V.B. carried out the Y2H assays and live confocal imaging experiments. F.I.Z. contributed to confocal imaging experiments and statistical analyses. A.M.P. contributed to confocal imaging experiments, statistical analyses and western blotting. E.J.M. designed and generated the isoform-specific mutant mice and quantified isoform-specific RNA. M.v.d.B. performed the GST pull-down experiments. S.T.M., F.M.S.d.V. and S.A.K. contributed to the human iPSC studies. J.S., J.M.K. and S.A.K contributed to the human postmortem tissue studies. D.C.R. and L.M.C.K. performed and analyzed the electrophysiological recordings, and contributed to the writing of the results section on these experiments. H.H. and M.W. contributed to patient recruitment, genotyping, and phenotype and clinical data collection and analyses. Y.E. and B.D. wrote the original draft of the manuscript. Y.E., B.D., S.A.K. and R.A.T. reviewed and edited the manuscript. Y.E., B.D., S.A.K., G.M.v.W. and E.J.M. supervised the research. Y.E., B.D. and S.A.K acquired funding.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Journal peer review information: Nature Neuroscience thanks Jon Huibregtse and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 UBE3A isoform expression in mouse and human brain.
a. Mouse genomic structure of the Ube3a locus indicating the annotated splice variants relative to the longest isoform (Iso 2; NCBI Reference Sequence: NM_011668.3). ATG refers to the initiation codon in exon 3 (UBE3A iso2 protein) and to the initiation codon at the border of exons 4/5 (UBE3A iso3 protein). mUbe3a-Iso1 (NCBI Reference Sequence: NM_173010.3) is a non-coding transcript, but note that this RefSeq has been removed from the NCBI database because of insufficient support for this transcript. b. Quantification of hUBE3A-Iso3 and hUBE3A-Iso1 of three independent human post-mortem cortical (PFC) lysates. Inset shows a representative UBE3A immunoblot. The image is vertically stretched to separate the isoform bands for better quantiifcation. Data are shown as the mean +/- SEM. n=3 biologically independent samples. c. Quantification of mUbe3a-Iso1 RNA transcript in wild-type and isoform specific mutant mice. qPCR was performed on cDNA generated from total cortex RNA obtained from WT (n=3 mice), mUbe3a-Iso2KO(n=2 mice) and mUbe3a-Iso3KO (n=3 mice) mice, by using a reverse primer in exon 11b and exon 13, to determine the relative level of isoform 1 transcripts. The level of exon 10- exon 11a containing transcripts was set at 100%. The relative amount of transcripts containing exon 10-11a-11b was determined using the same forward primer in exon 10 and a reverse in Exon 11b. Note that the levels of mUbe3a-Iso1 are very low compared to total Ube3a mRNA. Data are shown as the mean +/- SEM. d. Full Western blot used for Supplementary Fig. 1b.
Supplementary Figure 3 Identification of PSMD4 as binding partner of UBE3A by yeast two-hybrid screening.
a. Screening for UBE3A interacting proteins. Mouse UBE3A-Iso3 lacking the C- terminal 6 amino acids was fused in frame to the LexA DNA-binding domain and a custom yeast two-hybrid screen (Hybrigenics) was performed against 69 million independent mouse adult brain cDNA clones, fused to the Gal4 activation domain. Twenty-four high confidence clones, which activated multiple reporter genes, encoding four different proteins were isolated: PSMD4 (also known as Rpn10 or S5a), UbcH7 (cognate E2 of UBE3A), and two novel UBE3A binding proteins, NSUN2 and RPH3A. b. Mapping of the binding sites on UBE3A with the four identified proteins. The full-length mUBE3A-Iso3 (FL) interacts strongly with PSMD4 and NSUN2 while the interaction with RPH3A and UbcH7 is relatively weak. Deletion of the UBE3A AZUL domain (Δ76, the domain required for nuclear localization of mUBE3A-Iso3) results in loss of interaction with PSMD4 while interaction with the other three binding partners is unaffected. Upon further deletion the interaction with RPH3A (Δ274) and NSUN2 (Δ466) is also lost, showing that the three binding partners, PSMD4, NSUN2 and RPH3A, interact with different regions in the N-terminal domain of UBE3A. All N-terminal deletion constructs harbor the HECT domain, which is the binding site for UbcH7 and therefore, the interaction with UbcH7 is unaffected. Strength of interaction was determined by spotting serial dilutions of yeast cells co-transformed with bait and prey plasmids on solid medium plates without histidine or without histidine and containing 10 mM 3-AT (3-amino triazol; inhibitor of the HIS3 gene product), or without histidine and containing 20 mM 3-AT or on medium lacking adenine. ‘–,’ indicates no growth on His- plates (no interaction); +, growth on His- plates (weak interaction); ++, growth on His- +10 mM 3AT plates (medium strength interaction); +++, growth on His- + 20 mM 3AT (strong interaction); ++++, growth on plates lacking adenine(Ade-) (very strong interaction). c. Graphical representation of the Selected Interaction Domain (SID) of PMSD4, based on the shared overlap of the 6 independently identified PMSD4 clones in the two-hybrid screen (see panel A). Indicated are the von Willebrand factor type A domain (VWA; green) and the two Ubiquitin Interacting Motifs (UIMs) of PMSD4. The SID encompasses approximately half of the second UIM and most of the C- terminal tail (amino acids 288-352).
Supplementary Figure 4 Full Western blots from Fig. 4b.
a-c. Full Western blots used for Fig. 4b showing the pull-down (PD) experiment verifying the interaction of the AZUL domain with PSMD4.
Supplementary Figure 5 Association of endogenous UBE3A with proteasomes and identification of the regions in PSMD4 required for interaction with UBE3A.
a. Association of endogenous UBE3A with proteasomes. Affinity purification of proteasomes from HEK293TRPN11-HTBH cells stably expressing biotinylated Rpn11 (subunit of the 19S regulatory particle) and wild-type HEK293T (control) cells using streptavidin beads. The fractions were analyzed by SDS-PAGE and immunoblotting with the indicated antibodies. 5% of the total lysates (TL) and unbound fractions (UF) were loaded as compared to the affinity-purified proteasome fractions (PF) in all immunoblots, except for the streptavidin immunoblot where equivalent volumes of each fraction were loaded. The UBE3A immunoblot shows that approximately 5% of total, endogenous, UBE3A is recovered in the proteasome fraction (PF) derived from HEK293TRPN11-HTBH cells, but is absent in the PF from control HEK293T cells. The HEK293TRPN11-HTBH derived PF is highly enriched for the α/β subunits of the 20S core particle and PSMD4, a subunit of the 19S regulatory particle, demonstrating that the 20S core particle is recovered together with the 19S regulatory particle. Affinity-purified proteasomes contain very little cross-contaminating proteins as shown by the absence of GAPDH, an abundant cytosolic protein, in the PF fraction. The streptavidin immunoblot shows the high recovery of biotinylated RPN11, which was used as the affinity tag to pull-down proteasomes. Data are representative blots from three independent experiments with similar results obtained. b. Yeast two-hybrid analysis of the UBE3A-PSMD4 interaction. The indicated PSMD4 deletion constructs in a prey vector were tested against mUBE3A-Iso3 in a bait vector. Neither the deletion of the VWA domain nor the UIM domains of PSMD4, either separately or in combination, abrogates the interaction with UBE3A, indicating that these structural domains in PSMD4 are not required for UBE3A interaction. Strength of interaction was determined as described in the legend to Supplementary Fig. 3b. c. Multiple sequence alignment of PSMD4 tails from multicellular species (BoxShade 3.21 software): Mus musculus (Mm); Homo sapiens (Hs); Danio rerio (Dr); Drosophila melanogaster (Dm). Residues that are identical in at least three proteins are shaded black, while similar residues are shaded grey. Indicated by a red asterisk is the strictly conserved valine residue (V344 in mouse) that was mutated in this study. Data are representative blots from three independent experiments with similar results obtained. d,e. Full Western blots used for Supplementary Fig. 5a showing the association of endogenous UBE3A with proteasomes.
Supplementary Figure 7 Interaction PSMD4 with AS-associated missense mutations in UBE3A.
a. Two-hybrid interaction analysis of PMSD4 with selected AS-associated missense mutations which are distributed broadly throughout UBE3A and covering all mutational hotspots (see Fig. 6). Mutations are indicated in the human (h) UBE3A-Iso1 and mouse (m) UBE3A-Iso3 numbering, but were all assessed in mUBE3a-Iso3. Only the hUBE3AG593R/mUBE3AG590R mutant shows a strongly reduced interaction with PSMD4. The interaction of wild-type (WT) mUBE3a-Iso3 is shown as control. Strength of interaction as indicated in the left column, was determined in a spot assay on selective growth media, as described in the legend to Supplementary Fig. 3b. A representative yeast two-hybrid spot assay is shown on the right. Representative data from 3 independent experiments with similar results obtained. b. Pull-down (PD) experiment to verify the interaction of the PSMD4-tail with the AS-associated p.G20V, p.C21Y and p.G593R mutations (human UBE3A isoform 1 numbering). GST pull-down experiments were performed on lysates of E. coli cells expressing HA-mUBE3A-Iso3FL, HA-mUBE3A-Iso3ΔAZUL, HA-mUBE3A-Iso3G20V, HA-mUBE3A-Iso3C21Y or HA-mUBE3A-Iso3G590R (G593R in human UBE3A-Iso1). Lysates were incubated with GST-PSMD4324-379 (GST-PSMD4 tail) bound to glutathione beads or GST only (negative control). Eluted proteins were analyzed by immunoblotting using the indicated antibodies (right panels). Inputs (5%) are shown in the left panel. Representative data from 2 independent experiments with similar results obtained. c,d. Full Western blot used for Supplementary Fig. 7b showing the pull-down (PD) experiment to verify the interaction of the PSMD4-tail with the AS-associated p.G20V, p.C21Y and p.G593R mutations (human UBE3A isoform 1 numbering).
Supplementary Figure 8 AS-associated missense mutations in the Zn-finger of the AZUL domain and in the HECT domain disturb nuclear localization of UBE3A via distinct mechanisms.
a. Localization of UBE3A isoforms and AS-linked UBE3A mutants in HEK293T cells. Cells transfected with the indicated UBE3A-GFP constructs were imaged by live cell confocal microscopy approximately 48 hours after transfection. Data are representative images from three independent experiments with similar results obtained. b. AS-associated Zn-finger mutations p.G20V and p.C21Y abrogate nuclear localization. Mouse E16.5 derived hippocampal AS neurons transfected at DIV5 with mUBE3A-Iso3G20V, mUBE3A-Iso3C21Y or mUBE3A-Iso3 (WT) were fixed at DIV7. a-b, scale bar: 50 μm. Data are representative images from three independent experiments with similar results obtained.
Supplementary Figure 9 Full Western blot used for Fig. 7b.
Full Western blot for Fig. 7b showing selective expression of mUBE3A isoforms in targeted mutants.
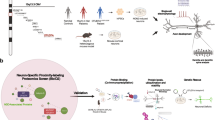
Supplementary Figure 10 Model for UBE3A localization.
Nuclear localization of UBE3A is controlled by two steps: (1) PSMD4 mediated import into the nucleus of both isoforms (mediated by the AZUL domain, as well as sequences in the HECT domain which may be required for proper folding) and (2) selective retention of the mUBE3A-Iso3 isoform in the nucleus by means of a functional Zn-finger. The N-terminal extension of mUBE3A-Iso2 interferes with nuclear retention. The AS-associated p.G20V, p.C21Y and p.G593R mutations (human UBE3A isoform 1 numbering; mouse UBE3A-Iso3 p.G590R) cause mislocalization of UBE3A by either affecting import in the nucleus (p.G593R) or by affecting nuclear retention (p.G20V, p.C21Y). See discussion for more details about this model. TF: transcription factor.
Supplementary information
Supplementary Information
Supplementary Figures 1–10 and Supplementary Tables 1 and 2.
Supplementary Table 3
Oligo used in this study.
Rights and permissions
About this article

Cite this article
Avagliano Trezza, R., Sonzogni, M., Bossuyt, S.N.V. et al. Loss of nuclear UBE3A causes electrophysiological and behavioral deficits in mice and is associated with Angelman syndrome. Nat Neurosci 22, 1235–1247 (2019). https://doi.org/10.1038/s41593-019-0425-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-019-0425-0
This article is cited by
-
A cross-species spatiotemporal proteomic analysis identifies UBE3A-dependent signaling pathways and targets
Molecular Psychiatry (2022)
-
Profiling Analysis of Circular RNA and mRNA in Human Temporal Lobe Epilepsy with Hippocampal Sclerosis ILAE Type 1
Cellular and Molecular Neurobiology (2022)
-
Evaluation of UBE3A antibodies in mice and human cerebral organoids
Scientific Reports (2021)
-
Deleting a UBE3A substrate rescues impaired hippocampal physiology and learning in Angelman syndrome mice
Scientific Reports (2021)
-
Mono-ubiquitination of Rabphilin 3A by UBE3A serves a non-degradative function
Scientific Reports (2021)
