
Abstract
Antiretroviral therapy is highly effective in suppressing human immunodeficiency virus (HIV)1. However, eradication of the virus in individuals with HIV has not been possible to date2. Given that HIV suppression requires life-long antiretroviral therapy, predominantly on a daily basis, there is a need to develop clinically effective alternatives that use long-acting antiviral agents to inhibit viral replication3. Here we report the results of a two-component clinical trial involving the passive transfer of two HIV-specific broadly neutralizing monoclonal antibodies, 3BNC117 and 10-1074. The first component was a randomized, double-blind, placebo-controlled trial that enrolled participants who initiated antiretroviral therapy during the acute/early phase of HIV infection. The second component was an open-label single-arm trial that enrolled individuals with viraemic control who were naive to antiretroviral therapy. Up to 8 infusions of 3BNC117 and 10-1074, administered over a period of 24 weeks, were well tolerated without any serious adverse events related to the infusions. Compared with the placebo, the combination broadly neutralizing monoclonal antibodies maintained complete suppression of plasma viraemia (for up to 43 weeks) after analytical treatment interruption, provided that no antibody-resistant HIV was detected at the baseline in the study participants. Similarly, potent HIV suppression was seen in the antiretroviral-therapy-naive study participants with viraemia carrying sensitive virus at the baseline. Our data demonstrate that combination therapy with broadly neutralizing monoclonal antibodies can provide long-term virological suppression without antiretroviral therapy in individuals with HIV, and our experience offers guidance for future clinical trials involving next-generation antibodies with long half-lives.
Similar content being viewed by others
Main
Modern antiretroviral therapy (ART) enables the near-complete suppression of plasma viraemia in most individuals with HIV1. However, difficulties in adherence to a lifetime of medication, long-term side effects and the possibility for developing antiretroviral drug-resistant virus have prompted intense research aimed at developing new therapies to achieve sustained virological remission without the need for daily ART4. In this regard, considerable efforts have focused on eliminating the persistent HIV reservoir5,6—one of the major impediments to viral eradication7,8,9—in individuals with HIV receiving clinically effective ART10. However, despite decades of research, it is becoming increasingly clear that complete eradication of the persistent HIV reservoir in an individual with infection is not feasible using currently available approaches and therapies11. In the absence of an effective vaccine against HIV and/or therapeutic agents that can eradicate the persistent viral reservoir, long-term suppression of plasma viraemia by infrequent administration of long-acting antiretroviral drugs12,13,14 or broadly neutralizing monoclonal antibodies (bNAbs)15,16,17,18 against HIV remains the most realistic approach for achieving ART-free virological suppression. In this regard, passive transfer of single bNAbs in individuals with HIV in the context of analytical treatment interruption (ATI) has shown limited success in part due to the presence of pre-existing and/or emergent antibody-resistant virus19,20,21,22. To mitigate this shortcoming, a combination approach involving the use of two bNAbs that bind to different regions of the HIV Env glycoprotein has led to promising results15,23. One such study used two bNAbs—one targeting the CD4-binding site on the HIV Env spike (3BNC117) and the other targeting the V3 loop and surrounding glycans (10-1074)—demonstrated that three infusions over a period of 6 weeks significantly delayed plasma viral rebound in individuals with infection after ATI23. Furthermore, previous studies have suggested that, besides their ability to neutralize HIV, certain bNAbs could potentially mediate the clearance of persistent viral reservoirs and/or enhance host immunity against the virus24,25,26,27. Accordingly, we conducted this study to further investigate the long-term effect of treatment with a combination of the bNAbs 3BNC117 and 10-1074 on the suppression of plasma viraemia and the dynamics of persistent viral reservoirs and their effect on immune parameters/responses in individuals with HIV.
Study design and participants
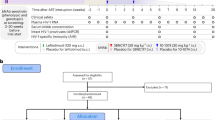
Between September 2018 and January 2021, we conducted a phase 1 clinical trial to assess the safety, tolerability and efficacy of the combination of the bNAbs 3BNC117 and 10-1074 in individuals with HIV. This trial comprised two components: (1) a randomized, double-blind, placebo-controlled study involving 14 participants in whom ART was initiated during the acute/early phase of infection and who subsequently underwent ATI shortly after receiving the first infusion of bNAbs or placebo (group 1); and (2) an open-label study involving 5 individuals with viraemic control who were naive for ART and had baseline plasma viraemia of between 200–5,000 copies per ml (group 2) (Table 1, Extended Data Fig. 1 and Supplementary Table 1). The study participants were not prescreened for the sensitivity of their HIV to 3BNC117 and 10-1074 before enrolment (the study inclusion criteria are described in the Methods). The randomized controlled portion of the study had a planned enrolment of 30 study participants. However, the study was prematurely halted in March 2020 owing to the increased safety concerns associated with ATI in the setting of the COVID-19 pandemic.
The study participants in the antibody arm of group 1 and group 2 received 4–8 (median, 8) 3BNC117 (30 mg kg−1) and 10-1074 (30 mg kg−1) infusions— two in the first month and once monthly thereafter (Fig. 1a). The infusions of bNAbs were well tolerated, with most adverse events being mild-to-moderate (grade 1 or 2) transient symptoms, including chills and/or fever (Supplementary Table 2). One study participant in group 1 (participant 01) experienced grade 1 fever and grade 2 rigors during the first 3BNC117 infusion and did not receive the first dose of 10-1074. Thus, this participant was effectively on antibody monotherapy for the first 2 weeks of the study. For subsequent infusions, participant 01 was premedicated with ibuprofen and completed the remaining study infusions of both antibodies with minimal (grade 1) or no reactions. No participant discontinued study medications due to antibody-related adverse events. There was no grade 3 or higher adverse event, including serious adverse events that were judged to be possibly, probably or definitely related to the study drugs. The baseline median CD4+ T cell counts for the antibody and the placebo arm of group 1 were 799 and 612 cells per µl, respectively (Table 1). The baseline median CD4+ T cell count of group 2 was 640 cells per µl (Table 1).

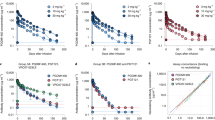
a, Schematic of the clinical trial design. The blue triangles indicate infusions of 3BNC117 and 10-1074 or placebo. F/U, follow-up; W, week. b, Plasma viraemia of the study participants. The participants in group 1 were randomized to receive either a combination of anti-HIV broadly neutralizing antibodies (bNAbs) 3BNC117 (30 mg kg−1) and 10-1074 (30 mg kg−1) (n = 7) or placebo (n = 7). ART was discontinued 3 days after receiving the first dose of bNAbs or placebo. Group 2 (open-label) consisted of 5 ART-naive individuals with viraemic control. The blue triangles indicate the time of administration of the antibodies or placebo. The grey shaded boxes indicate the duration of ATI. The grey dotted horizontal lines indicate the limit of detection of the assay (40 copies of HIV RNA per ml). The white triangles indicate undetectable plasma viraemia (<40 copies of HIV RNA per ml) and the red triangles indicate detectable plasma viraemia (≥40 copies of HIV RNA per ml). Plasma antibody concentrations were determined using the TZM-bl assay. The dark blue and green circles indicate the plasma concentration of 3BNC117 and 10-1074, respectively. The x axis label (time (weeks)) applies to all of the plots in b. c, Kaplan–Meyer analysis of suppression of plasma viraemia after ATI in the study participants in group 1. Comparison of the proportion of the study participants remaining off ART during ATI in the bNAb versus placebo arm (left). The vertical dotted line indicates the virological end point. Right, the duration of plasma viraemia at below 200 copies of HIV RNA per ml after ATI was compared between the participants in the bNAb arm and the placebo arm. P values were determined using exact log-rank tests. d, Comparison of plasma viraemia at the time of ART reinitiation between the bNAb (n = 6) and placebo (n = 6) arms in the group 1 study participants. The grey bars represent the geometric mean values. The P value was determined using a two-sided Mann–Whitney U-test.
Effect of bNAbs on virological parameters
Three days after receiving the first infusion of 3BNC117 and 10-1074 or placebo, the study participants in group 1 underwent ATI and plasma viraemia and CD4+ T cell counts were measured every 2 weeks (Fig. 1a). For group 1, the protocol-predefined virological end point was the difference between the bNAb and placebo arms in the number of study participants who experienced plasma viral rebound and met the criteria to restart ART before study week 28. As shown in Fig. 1b, c, 6 out of the 7 study participants in the placebo arm experienced plasma viral rebound and met the criteria to restart ART before study week 28 compared with none of the 7 participants in the treatment arm. The median duration off ART was 39.6 weeks (range, 9.9–49.6 weeks) and 9.4 weeks (range, 5.3–26 weeks) for the group 1 bNAb and placebo participants, respectively (P = 0.001; Fig. 1c (left)). Notably, participant 11, in the placebo arm, who did not meet criteria to restart ART, deviated from the protocol and surreptitiously resumed ART at study week 12 owing to concerns about rising plasma viraemia. Participant 14, in the bNAb arm, reinitiated ART before meeting the restart criteria due to concerns associated with the COVID-19 pandemic. Thus, the data from these participants were censored for end-point analysis.
In an additional post hoc analysis, 5 out of the 7 study participants in the bNAb arm of group 1 maintained suppression of plasma viraemia (<40 copies per ml), whereas all of the study participants in the placebo arm of group 1 experienced plasma viral rebound within the first 8 weeks of ATI. The median duration of plasma viraemia suppression at <200 copies per ml in group 1 was 33.4 weeks (range, 7.4–43.3 weeks) and 3.4 weeks (range, 1.9–7.9 weeks) in the bNAb and placebo arms, respectively (P = 0.002; Fig. 1c (right)). Notably, two group 1 bNAb study participants (01 and 14), whose plasma viraemia rebounded by more than 200 copies per ml within 8 weeks into ATI, carried bNAb-resistant, replication-competent HIV in their CD4+ T cells at the baseline (Fig. 2a). The median duration of suppression of plasma viraemia at below 200 copies per ml for the participants with sensitive virus was 35.4 weeks (range, 31.7–43.3 weeks). There was no difference in the level of plasma viraemia before reinitiation of ART between the bNAb and placebo arms in the group 1 study participants (P = 0.82; Fig. 1d), suggesting that bNAb-mediated virological suppression may not necessarily alter the kinetics of plasma viral rebound after clearance of the antibodies in vivo. In group 2, 2 out of the 5 study participants whose baseline infectious HIV was sensitive to both antibodies maintained complete suppression of plasma viraemia for an average of 41.7 weeks (Figs. 1b and 2a). Taken together, these results demonstrate that the combination therapy with 3BNC117 and 10-1074 is highly effective in suppressing HIV in the absence of ART for extended periods, provided that antibody-resistant virus is not present at the baseline.

a, The IC80 concentrations of 3BNC117 and 10-1074 against autologous, replication-competent viral isolates from the study participants. Near-clonal replication-competent HIV isolates were generated by co-culturing autologous CD4+ T cells with activated CD4+ T cells from healthy HIV-seronegative donors as described in the Methods. The IC80 concentrations of 3BNC117 and 10-1074 against infectious HIV isolates were determined using the TZM-bl target cell neutralization assay. The grey bars indicate the geometric mean values. The x axis label (time (weeks)) applies to all of the plots in a. b, The sensitivity of infectious HIV isolates derived from participant 02 to 3BNC117 and 10-1074. The IC80 concentrations of 3BNC117 and 10-1074 against replication-competent viral isolates derived during ATI and after the reinitiation of ART were determined to examine the decay characteristics of antibody-resistant virus over time. c, Pharmacokinetics of 3BNC117 and 10-1074 in study participants. The plasma levels of 3BNC117 and 10-1074 were determined on the basis of a validated luciferase-based neutralization assay in TZM-bl cells. The red and green lines represent the geometric mean values of 3BNC117 and 10-1074, respectively.
The sensitivity (80% inhibitory concentration (IC80)) of replication-competent HIV isolates to 3BNC117 and 10-1074 was longitudinally monitored in the study participants before and after the infusions of bNAbs. At the baseline, 38% of the study participants in group 1 had resistant (IC80 > 10µg ml−1) infectious HIV to either 3BNC117 (3 out of 13 participants, 23%), 10-1074 (3 out of 13 participants, 23%), or both bNAbs (1 out of 13 participants, 8%) (Fig. 2a). There was no evidence that antibody-resistant virus emerged over time for the participants (02, 05, 06, 08 and 13) in the bNAb arm of group 1 who maintained sustained virological suppression during ATI (Fig. 2a). Notably, participant 01, whose baseline virus was largely resistant to 3BNC117, started developing resistance to 10-1074 at week 16 after ATI. Moreover, the baseline virus of participant 14, whose plasma viraemia rebounded >40 copies per ml within 6 weeks after ATI, was highly resistant to both antibodies (Fig. 2a), although only one infectious isolate could be examined due to the extraordinarily low frequency of CD4+ T cells carrying replication-competent HIV. Three out of the five group 2 participants, whose baseline CD4+ T cells contained resistant infectious HIV to either bNAb, did not suppress their plasma viraemia and subsequently developed fully resistant virus to both 3BNC117 and 10-1074 (Fig. 2a). Notably, group 1 bNAb participant 02, who suppressed plasma viraemia for more than 43 weeks in the absence of ART, developed a marked resistance to 10-1074 (but not to 3BNC117) after plasma viral rebound, probably due to a longer half-life of 10-1074 compared with 3BNC117 that resulted in subtherapeutic concentrations in vivo23 (Fig. 2b). After reinitiation of ART according to the protocol in participant 02, the proportion of CD4+ T cells carrying 10-1074-resistant replication-competent HIV gradually diminished over time, suggesting a natural decay of the viral reservoir carrying 10-1074-resistant virus in the absence of immunologic pressure26. The precise half-life of each antibody could not be assessed in our study due to the unavailability of plasma immediately after each infusion. However, the overall pharmacokinetics observed in our study participants were similar to those of a previous study23 (Fig. 2c) . Collectively, our data suggest that bNAb therapy with 3BNC117 and 10-1074 does not lead to a precipitous accumulation of resistant HIV during long-term virological suppression, provided that antibody-resistant virus is not present at the baseline and serum antibody titres remain high.
The levels of CD4+ T cells carrying total HIV DNA, cell-associated HIV RNA, intact proviral DNA and replication-competent virus were longitudinally monitored in group 1 bNAb participants whose plasma viraemia remained suppressed >30 weeks after ATI (Fig. 3a). No statistical significance was reached when the levels of HIV reservoirs were compared between the two time points (weeks 0 and 24) (Fig. 3b). Furthermore, a comparison of the fold changes in the size of HIV reservoirs over time between the group 1 bNAb arm and a cohort of early-treated individuals who remained continuously on ART28 showed no significant difference (Fig. 3c). A marked increase in the frequency of CD4+ T cells carrying total HIV DNA was observed in the group 1 placebo participants after ATI (Extended Data Fig. 2a). Notably, a reduction (transient for participant S3) in the level of total HIV DNA in the CD4+ T cells was observed in group 2 participants whose plasma viraemia was suppressed after bNAb infusions (Extended Data Fig. 2b). These data suggest that the combination bNAbs did not have a significant effect on the persistent HIV reservoir in our study participants, although a long-term study involving a much larger cohort would be necessary to definitively address this question.

a, Frequencies of CD4+ T cells carrying total HIV DNA, cell-associated HIV RNA, intact proviral DNA and replication-competent HIV are shown in five study participants in the bNAb arm of group 1 in whom sustained virological suppression was achieved after ATI. b, Dynamics of HIV reservoirs in the CD4+ T cells of group 1 bNAb study participants (n = 5) before and after multiple infusions of 3BNC117 and 10-1074. The black lines indicate the geometric mean values. Statistical significance was tested using two-sided Wilcoxon matched-pairs signed-rank tests. c, Comparison of the fold changes in the size of HIV reservoirs over time between the bNAb arm (n = 5) and a control cohort of individuals with infection (n = 13) who initiated ART during the acute/early phase of infection and did not undergo therapeutic interventions or ATI. The black lines indicate the median values. P values were determined using two-sided Mann–Whitney U-tests.
The effect of bNAbs on immune parameters
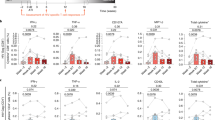
Throughout the bNAb infusion periods, the CD4+ T cell counts of all of the participants, including those in the placebo arm of group 1, remained stable (Extended Data Fig. 3a). Nonetheless, we considered whether immunologic abnormalities (such as inflammation and exhaustion)29 occurred in study participants who received infusions of combination bNAbs and maintained extended periods of virological suppression. To this end, we used high-dimensional flow cytometry to longitudinally examine immune parameters in peripheral blood mononuclear cells of our study participants. The frequencies of TIGIT+, PD-1+, CD38+HLA-DR+ and subsets of CD8+ T cells remained unchanged over time in all of the groups (Extended Data Fig. 3b). Optimized t-distributed stochastic neighbour embedding using high-dimensional mapping of the 21-parameter flow cytometry data was conducted to monitor immune cells of the study participants over time (Extended Data Fig. 4a, b). A total of 15 distinct T cell clusters was identified by FlowSOM30 analyses (Extended Data Fig. 4a–c). The levels of CD8+ T cell clusters remained stable over time in all of the groups (Extended Data Fig. 4d). Cytokines (IL-6, IL-8 and TNF), chemokines (MIP-1β and RANTES) and markers of immune activation (IL-2Rα, CD40, PD-L1, IP-10, C-reactive protein and d-dimer) measured in the plasma of the study participants between week 0 and week 24 all remained unchanged in all groups (Extended Data Fig. 5). Taken together, our data suggest that substantial immunologic abnormalities, as assessed by cellular phenotypes and plasma biomarkers, did not arise during prolonged ATI in study participants in whom the bNAb-mediated virological suppression was achieved.
Previous studies have demonstrated that passive transfer of bNAbs in SIV-infected animals24,31 and humans with HIV25,26, including individuals whose plasma viraemia was controlled by bNAbs in the absence of ART27, could potentially lead to enhanced antiviral immunity. In humans, HIV-Gag-specific CD4+ and CD8+ T cell responses peaked at week 6/7 and were no longer significant by week 18 (ref. 27). We longitudinally examined HIV-specific CD8+ T cells in our study participants by performing intracellular cytokine staining after stimulation with a pool of overlapping HIV Gag peptides and molecular analyses of the HIV-specific CD8+ T cell receptor (TCR) repertoire. The level of polyfunctional (IFNγ+TNF+MIP-1β+) HIV-Gag-specific CD8+ T cells remained unchanged in group 1 bNAb and group 2 participants (Extended Data Fig. 6a). A modest increase in the frequency of HIV-specific CD8+ T cells was observed over time in the group 1 placebo participants, probably due to the rising plasma viraemia after ATI (Extended Data Fig. 6a). To gain further insights into the effect of combination bNAbs on HIV-specific T cells at the molecular level, we conducted longitudinal deep-sequencing analysis of TCR genes using genomic DNA isolated from highly enriched CD8+ T cells. The average breadth of HIV-specific CD8+ TCR clonotypes remained stable in group 1 bNAb participants (Extended Data Fig. 6b). A modest increase, albeit statistically not significant (P = 0.062), in the depth of HIV-specific CD8+ TCR clonotypes was observed in these participants after passive transfer of the bNAbs. Notably, the breadth and depth of HIV-specific CD8+ TCR clonotypes in the group 1 placebo participants remained largely unchanged over time. There was a modest increase, albeit statistically not significant (P = 0.062), in the breadth of HIV-specific CD8+ TCR clonotypes in group 2 over time, possibly due to ongoing viral replication in 3 out of the 5 participants. To further investigate the changes in the TCR repertoires of the study participants, we compared the frequencies of the HIV-specific clonotypes at three different time points. For each group, the frequencies of the clonotypes present in the top 25 comparisons (Supplementary Table 3) that were ranked on the basis of statistical significance and the usage levels of the TRBV–TRBJ gene pairs in these clonotypes (Supplementary Table 4) were used to conduct principal component analysis. On the basis of the clonotype frequencies and gene usage levels, two reduced states for each TCR repertoire were computed. The variability of both repertoire states decreased at week 24 compared with at week 0 among the study participants in the bNAb arm of group 1, as illustrated by the smaller-sized ellipses over time (Extended Data Fig. 6b). A larger study involving multiple infusions of bNAbs and extended periods of follow-up will be necessary to confirm these findings.
Discussion
Despite decades of intensive research, the prospects of achieving an HIV cure as manifested by eradication of the virus remain elusive2. A more realistic alternative to a cure may be through immune-based therapies aimed at achieving maximal virological suppression without the need for life-long and predominantly daily ART and without the need to eliminate the persistent HIV reservoir in individuals with infection32,33. To date, one of the most promising and realistic approaches for accomplishing these outcomes is by infrequent passive infusions of combination HIV-specific bNAbs15,16,17,18. Numerous HIV-specific bNAbs have been isolated in recent years16,17,34,35 and more than a dozen of them, either as single or combination therapies, have been tested in humans for safety and therapeutic efficacy15. However, although certain bNAbs have been shown to be highly effective in neutralizing HIV in vitro by standardized assays involving large panels of HIV Env-pseudotyped viruses35, it is becoming increasingly clear that durable and ART-free virological suppression will require a combination of two or more antibodies together with screening for bNAb resistance to the pre-existing virus in individuals with infection19,22. The data obtained from our clinical trial, a randomized study involving participants in whom ART was initiated during the acute/early phase of infection with a long duration of follow-up, clearly demonstrated that complete and sustained virological suppression is achievable with intermittent administration of combination bNAbs. However, the combination bNAbs was ineffective in maintaining suppression of plasma viraemia in 2 out of 7 study participants in our small cohort in whom replication-competent HIV exhibited resistance to one or both antibodies at the baseline. This will potentially pose a substantial challenge for the treatment of individuals with HIV with bNAbs on a large scale. Nonetheless, our findings revealed several positive outcomes. Over the course of 24 weeks of virological suppression in the study participants who received the combination bNAbs, there were no significant immunologic or virological alterations, such as increases in immune activation and exhaustion or the size of HIV reservoirs. A recent study has demonstrated that combination bNAbs could further augment T-cell responses against HIV in individuals with infection while suppressing their plasma viraemia27. Although we did not observe any significant changes in the frequency of HIV-specific polyfunctional CD8+ T cells in our study, further studies with early and frequent time points27, as well as detailed assessments of virus-specific proliferative and cytotoxic effector functions36, may be needed to fully delineate the role of T cells in bNAb therapeutic agents. It is also plausible that the marked virological suppression mediated by the bNAbs in our study cohort (those who initiated ART during the acute/early phase of infection), combined with their relatively low HIV reservoir burden at baseline, may have prevented the formation of immune complexes18 and/or the residual levels of viral expression needed for the induction of robust antiviral T-cell responses33,37. It also remains to be fully elucidated whether additional benefits of bNAbs, aside from their ability to neutralize HIV and maintain viral suppression in the absence of ART, can ultimately alter the course of infection (that is, long-term suppression of plasma viraemia in the absence of both bNAbs and ART) after clearance of the antibodies in vivo. Although a larger study involving multiple infusions of bNAbs and extended periods of follow-up will be necessary to address this question, our data strongly suggest that the virus will ultimately rebound after bNAb clearance after 24 weeks of bNAb therapy. In this regard, future trials may need to incorporate a prespecified ART initiation shortly after the last infusion of bNAbs to prevent the emergence of antibody-resistant virus in study participants.
One of the major confounding factors of this study is the small sample size and it is therefore essential to conduct a much larger study involving study participants in whom ART was initiated during the acute/early phase of infection to confirm our findings. Other major caveats of our study include the lack of lymphoid tissue analyses and the limited number of infusions (up to eight). Nonetheless, our findings offer clear evidence that combination bNAb therapy in individuals with HIV is safe and well tolerated and, for those with antibody-sensitive virus, offers marked virological suppression without any significant or unforeseen immunologic and virological anomalies. As the next generation of bNAbs with increased breadth and prolonged half-lives (>60 days)15,17,35 become available, there is a reason to believe that the infrequent administration (that is, twice a year) of such antibodies, possibly along with a long-acting injectable antiretroviral drug12,13,14, could lead to ART-free HIV suppression for extended periods (years) in individuals with infection.
Methods
Study design, participants and procedures
The purpose of this clinical trial was to assess the safety, tolerability and virological efficacy of a combination of two bNAbs 3BNC117 and 10-1074 in individuals with HIV. The study consisted of two components: a randomized, double-blind, placebo-controlled trial (group 1) and an open-label, single-arm trial (group 2). The study inclusion criteria for group 1 included start of ART within 12 weeks of being diagnosed with acute or early HIV infection, a CD4+ T cell count of >450 cells per µl at screening, continuous ART treatment with suppression of plasma viraemia below the limit of detection for ≥1 year and with general good health. The participants who had previously participated in an ATI study were not excluded. Acute infection was defined as plasma viraemia of greater than 2,000 copies of HIV RNA per ml with a negative HIV-1 enzyme immunoassay (EIA; criterion 1); a positive result from an HIV-1 EIA with a negative or indeterminate HIV-1 western blot that subsequently evolves to a confirmed positive result (criterion 2); or a negative result from an HIV-1 EIA within the past 4 months and plasma viraemia greater than 400,000 copies per ml in the setting of a potential exposure to HIV-1 (criterion 3). Early infection was defined as a negative result from an HIV-1 EIA within 6 months before a positive result from an HIV-1 EIA and confirmatory HIV-1 western blot (criterion 4); a negative result from a rapid HIV-1 test within 1 month before a positive result from an HIV-1 EIA and western blot (criterion 5); or the presence of low level of HIV antibodies as determined by having a positive EIA and western blot with a non-reactive detuned EIA according to a multi-assay testing algorithm for recent infection (criterion 6). The study inclusion criteria for group 2 included no ART within 24 months, plasma viraemia of 200–5,000 copies per ml, and at least two documented instances of plasma viraemia ≥200 copies per ml on at least two occasions in the 12 months before screening. The predetermined primary end point of the study was the rate of occurrence of grade 3 or higher adverse events or serious adverse events that were probably or definitely related to the study antibodies. Adverse events were determined according to the Division of AIDS Table for Grading the Severity of Adult and Pediatric Adverse Events, Version 2.1. The virological end points were the number of study participants who experienced plasma viral rebound after ATI and who met criteria to restart ART before study week 28 (group 1) or the number of study participants who achieved sustained suppression of plasma viraemia by study week 28 (group 2).
The study participants in group 1 were randomized to receive multiple (up to 8) infusions of 3BNC117 (30 mg kg−1 body weight) and 10-1074 (30 mg kg−1 body weight) or placebo during the 24-week period. One study participant (participant 05) whose regimen contained a non-nucleoside reverse transcriptase inhibitor (NNRTI) was switched to an integrase inhibitor-based regimen 2 weeks before ATI due to the long half-life of NNRTIs. The study participants in group 1 discontinued ART 3 days after the first infusion of 3BNC117 and 10-1074 or placebo. The protocol investigators and study participants in group 1 were blinded to treatment assignments for the duration of study. All of the study participants in group 2 received up to 8 infusions of 3BNC117 (30 mg kg−1 body weight) and 10-1074 (30 mg kg−1 body weight) during the 24-week period. The study participants received sequential 3BNC117 and 10-1074 intravenously according to their body weight over a 60 min period per antibody. Plasma viraemia and CD4+ T cell count were monitored every 2 weeks. Plasma viraemia was determined using the Abbott Real-Time HIV-1 assay with a detection limit of 40 copies of HIV RNA per ml. The study participants in group 1 discontinued antibody infusions or placebo and reinitiated ART if they met one of more of the following criteria during the ATI phase: (1) a confirmed >30% decline in baseline CD4+ T cell count; (2) an absolute CD4+ T cell count of <350 cells per µl; or (3) sustained plasma viraemia of >1,000 copies per ml for greater than 4 weeks. The study participants in group 2 discontinued antibody infusions if their CD4+ T cells declined as specified above or they had an increase in baseline plasma viraemia of >0.5 log10.
Blood and leukapheresed products were collected in accordance with protocols approved by the Institutional Review Board of the National Institute of Allergy and Infectious Diseases, National Institutes of Health (ClinicalTrials.gov ID: NCT03571204). All of the participants provided written informed consent.
Measurements of IC80
Near-clonal replication-competent HIV isolates were derived from coculturing enriched CD4+ T cells of the study participants with CD8-depleted anti-CD3 stimulated peripheral blood mononuclear cells (PBMCs) from HIV-seronegative donors as described below. The concentration of each infectious viral isolate was initially determined by HIV p24 enzyme-linked immunosorbent assay (ELISA). Each isolate was titrated using a standard TZM-bl target cell assay. To determine the IC80 concentrations of both antibodies against reservoir-derived replication-competent HIV over time, each viral isolate was incubated with serially diluted (40, 10, 2.5, 0.625, 0.156, 0.039 µg ml−1 in duplicate) 3BNC117 and/or 10-1074 for 90 min followed by incubation with TZM-bl cells for 48 h. Cells were then lysed and substrate (Promega) was added to measure the luciferase activity (SparkControl, v.3.1, Tecan).
Pharmacokinetics of 3BNC117 and 10-1074
The levels of 3BNC117 and 10-1074 in the plasma of the study participants were longitudinally measured using a validated luciferase-based neutralization assay in TZM-bl cells as previously described38. In brief, plasma samples were tested in duplicate using a primary 1:20 dilution with a fivefold titration series and tested against the HIV Env pseudotyped viruses Q461.e2 (3BNC117-sensitive/10-1074-resistant) and Du422.1 (10-1074-sensitive/3BNC117-resistant). Pseudotyped virus with murine leukaemia virus (MuLV) Env was used as a negative control. 3BNC117 and 10-1074 clinical drug products were used as positive controls and tested using a primary concentration of 10 μg ml−1 with fivefold serial dilution series. Plasma concentrations (µg ml−1) of 3BNC117 and 10-1074 were determined as follows: plasma 50% inhibitory dilution (ID50) titre (dilution) × IC50 titre (µg ml−1) of 3BNC117 or 10-1074.
Quantification of HIV reservoirs
The frequency of CD4+ T cells carrying total HIV DNA was determined by droplet digital PCR as previously described39. In brief, genomic DNA was isolated from highly enriched CD4+ T cells using the Puregene DNA extraction kit (Qiagen) and digested with restriction enzyme MscI (New England BioLabs). Subsequently, the digested genomic DNA was analysed using droplet digital PCR (QuantaSoft, v.1.7.4.0917, Bio-Rad) according to the manufacturer’s instructions. The following PCR primers and probe were used for amplification of the 5′ LTR region of HIV DNA: forward primer 5′-GRAACCCACTGCTTAAGCCTCAA-3′ (nucleotides 506–528 in HXB2; GenBank: K03455.1), reverse primer 5′-TGTTCGGGCGCCACTGCTAGAGA-3′ (nucleotides 648–626) and probe 5′-6FAM-AGTAGTGTGTGCCCGTCTGTT-IABkFQ-3′ (nucleotides 552–572). The following PCR primers and probe were used for amplification of the housekeeping gene RPP30: forward primer 5′-GATTTGGACCTGCGAGCG-3′ (nucleotides 29–46; GenBank: NM_001104546.2), reverse primer 5′-GCGGCTGTCTCCACAAGT-3′ (nucleotides 90–73) and probe 5′-HEX-TTCTGACCTGAAGGCTCTGCGC-IABkFQ-3′ (nucleotides 49–70). Copy numbers of HIV DNA were normalized per 1 × 106 CD4+ T cells.
The level of CD4+ T-cell-associated HIV RNA was determined by PCR with reverse transcription (RT–PCR). Total RNA was isolated from highly enriched CD4+ T cells using the RNeasy Mini kit (Qiagen), followed by synthesis of complementary DNA (cDNA) using the qScript XLT cDNA Master Mix (Quanta Biosciences) according to the manufacturer’s instructions. cDNA was analysed using droplet digital PCR (Bio-Rad Laboratories) using the following primers: HIV-specific primers HIV-US-F (5′-TCTCTAGCAGTGGCGCCCGAACA-3′, nucleotides 626–648), HIV-US-R (5′-TCTCCTTCTAGCCTCCGCTAGTC-3′, nucleotides 786–764) and HIV-US-probe (5′-6FAM-CAAGCCGAGTCCTGCGTCGAGAG-IABkFQ-3′, nucleotides 705–683); and TATA-box-binding protein (TBP; housekeeping gene)-specific primers TBP-F (5′-CACGAACCACGGCACTGATT-3′, nucleotides 863–882; GenBank: NM_003194.5) and TBP-R (5′-TTTTCTTGCTGCCAGTCTGGAC-3′, nucleotides 951–930) and TBP-probe (5′-HEX-TGTGCACAGGAGCCAAGAGTGAAGA/3-IABkFQ-3′, nucleotides 902–926). Copy numbers of cell-associated HIV RNA were normalized per 1 × 106 copies of TBP.
The frequency of CD4+ T cells carrying intact HIV proviruses was determined using the Intact Proviral DNA Assay (IPDA) as previously described40.
The level of CD4+ T cells carrying replication-competent HIV was determined by quantitative co-culture assay using serially diluted and replicates of 5 × 106 CD4+ T cells as previously described39. Highly enriched CD4+ T cells were incubated with anti-CD3 antibodies and irradiated PBMCs from healthy HIV-negative donors. After incubation for 1 day, 1 × 106 CD8-depleted and anti-CD3-stimulated PBMCs from HIV-negative donors were added to each well, followed by periodic removal of cell suspensions and replenishment with fresh medium containing IL-2. Subsequently, HIV p24 ELISA was performed on the culture supernatants to identify wells containing replication-competent HIV. The infectious units per million cells from quantitative coculture assays were determined as previously described41.
Phenotypic characterization of immune cells using flow cytometry
Cryopreserved PBMCs were thawed, washed and stained with the viability reagent Zombie NIR (BioLegend, 423106) and fluorophore-conjugated antibodies in Brilliant Stain Buffer Plus (BD, 566385). Flow cytometry data were acquired on the Cytek Aurora cytometer using the SpectroFlo Software (Cytek Biosciences) and analysed using FlowJo v.10.7.1 and the OMIQ platform (https://www.omiq.ai/).
High-dimensional data analysis of flow cytometry data
Optimized t-distributed stochastic neighbour embedding (opt-SNE) and FlowSOM analyses were conducted using OMIQ software (https://www.omiq.ai/). Opt-SNE analysis was performed using equal sampling of 100,000 CD3+ T cells from each FCS file, with 1,000 iterations, a perplexity of 30 and a theta of 0.5. The following immune markers were used to generate opt-SNE maps: CD4, CD8, CD45RA, CCR7, CD27, CD28, CD38, HLA-DR, CD226, TIGIT, PD-1, 2B4, CD160, CTLA-4, CD96, OX40, CXCR5, ICOS and 4-1BB. The resulting opt-SNE maps were used for the FlowSOM algorithm. The self-organizing map was generated using hierarchical consensus clustering and 15 meta-clusters were identified. The heat map displaying column-scaled z-scores of mean fluorescence intensity for individual FlowSOM clusters was generated using OMIQ platform.
Intracellular cytokine staining assay
The frequency of virus-specific CD8+ T cells was assessed by incubating PBMCs with a pool of HIV-1 consensus B Gag overlapping peptides (NIH AIDS Reagent Program) with brefeldin A (Sigma-Aldrich) for 6 h at 37 °C. Unstimulated cells were used as a negative control for background subtraction. Cells were stained with Zombie NIR (BioLegend, 423106) and antibodies to immune markers (Supplementary Table 5). Cells were then fixed with 1× lysing solution (BD Biosciences) and permeabilized with 1× permeabilization solution 2 (BD Biosciences). After washing, cells were stained with the antibodies to the intracellular cytokines and chemokines (Supplementary Table 5 and Supplementary Fig. 1). Data were acquired on the Cytek Aurora cytometer using the SpectroFlo Software (Cytek Biosciences) and analysed using FlowJo v.10.7.1.
TCR repertoire analysis
Complementarity determining regions (CDR) 3 of TCR β-chains present in highly enriched CD8+ T cells of the study participants were sequenced in a high-throughput manner using the immunoSEQ assay42,43 after amplification of the extracted DNA in a bias-controlled multiplex PCR. The resulting CDR3 sequences were collapsed and filtered to quantify the absolute abundance and frequency of each unique TCR β region with immunoSEQ Analyzer (Adaptive Biotechnologies)44. TCR repertoire statistics, including gene usage (the fraction of clonotypes in which a given TRBV or TRBJ gene is present), were computed using Immunarch45. TCR sequencing data are available online (https://clients.adaptivebiotech.com/login; login: chun-review@adaptivebiotech.com, password: chun2021review). HIV-specific breadth and depth were computed using the HIV-specific CDR3 sequences previously reported in the literature from four databases, namely the immune epitope database (IEDB)46, VDJdb47, McPAS-TCR48 and the Pan immune repertoire database (PIRD)49. The reported breadth values represent the fraction of clonotypes in each repertoire that are HIV-specific, whereas depth calculations take the abundance of each clonotype into account, such that each HIV-specific clonotype affects the overall HIV-specific depth (per sample) with a magnitude proportional to its abundance.
Measurements of biomarkers in plasma
Levels of biomarkers in plasma were determined using the ELLA platform (Simple Plex Runner, v.3.7.2.0, ProteinSimple) according to the manufacturer’s instructions.
Statistical analysis
P values for the virological end points (group 1) were determined using exact log-rank tests. Sensitivity analyses were performed to handle the two study participants (11 and 14) who reinitiated ART before meeting the restart criteria. Two independent analyses, with and without censoring the data from the above study participants, generated the same P values.
P values for paired and unpaired comparisons were computed using Wilcoxon signed-rank tests and Mann–Whitney U-tests, respectively, using Prism v.9.1 (GraphPad). P values computed in the TCR repertoire analysis were determined using Wilcoxon signed rank tests (paired) and Mann–Whitney U-tests (unpaired) using the R package ggpubr50. The factoMineR51 package in R was used to perform PCA with the TCR repertoire data.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
TCR sequencing data are available online (https://clients.adaptivebiotech.com/login; login: chun-review@adaptivebiotech.com, password: chun2021review). The HIV-specific CDR3 sequences were downloaded from the following four databases: the immune epitope database (IEDB; http://www.iedb.org/), VDJdb (https://vdjdb.cdr3.net), McPAS-TCR (http://friedmanlab.weizmann.ac.il/McPAS-TCR/) and the Pan Immune Repertoire Database (PIRD; https://db.cngb.org/pird/).
Code availability
The R scripts that were used in the data analysis have been deposited at GitHub (https://github.com/cihangenome/combination-antibodies-HIV). The following R packages were used: factoextra (v.1.0.7), FactoMineR (v.2.4), reshape (v.0.8.8), reshape2 (v.1.4.4), writexl (v.1.4.0), gdata (v.2.18.0), psych (v.2.1.9), car (v.3.0-11), carData (v.3.0-4), corrr (v.0.4.3), lubridate (v.1.8.0), readxl (v.1.3.1), forcats (v.0.5.1), stringr (v.1.4.0), purrr (v.0.3.4), readr (v.2.0.2), tidyr (v.1.1.4), tibble (v.3.1.5), tidyverse (v.1.3.1), ggpubr (v.0.4.0), immunarch (v.0.6.6), patchwork (v.1.1.1), data.table (v.1.14.2), dtplyr (v.1.1.0), dplyr (v.1.0.7) and ggplot2 (v.3.3.5).
References
Deeks, S. G., Lewin, S. R. & Havlir, D. V. The end of AIDS: HIV infection as a chronic disease. Lancet 382, 1525–1533 (2013).
Chun, T. W., Moir, S. & Fauci, A. S. HIV reservoirs as obstacles and opportunities for an HIV cure. Nat. Immunol. 16, 584–589 (2015).
Chun, T. W., Eisinger, R. W. & Fauci, A. S. Durable control of HIV infection in the absence of antiretroviral therapy: opportunities and obstacles. JAMA 322, 27–28 (2019).
Ndung’u, T., McCune, J. M. & Deeks, S. G. Why and where an HIV cure is needed and how it might be achieved. Nature 576, 397–405 (2019).
Cohn, L. B., Chomont, N. & Deeks, S. G. The biology of the HIV-1 latent reservoir and implications for cure strategies. Cell Host Microbe 27, 519–530 (2020).
Sengupta, S. & Siliciano, R. F. Targeting the latent reservoir for HIV-1. Immunity 48, 872–895 (2018).
Chun, T. W. et al. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl Acad. Sci. USA 94, 13193–13197 (1997).
Finzi, D. et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278, 1295–1300 (1997).
Wong, J. K. et al. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 278, 1291–1295 (1997).
Margolis, D. M. et al. Curing HIV: seeking to target and clear persistent infection. Cell 181, 189–206 (2020).
Lewin, S. R. & Rasmussen, T. A. Kick and kill for HIV latency. Lancet 395, 844–846 (2020).
Swindells, S. et al. Long-acting cabotegravir and rilpivirine for maintenance of HIV-1 suppression. N. Engl. J. Med. 382, 1112–1123 (2020).
Orkin, C. et al. Long-acting cabotegravir and rilpivirine after oral induction for HIV-1 infection. N. Engl. J. Med. 382, 1124–1135 (2020).
Overton, E. T. et al. Long-acting cabotegravir and rilpivirine dosed every 2 months in adults with HIV-1 infection (ATLAS-2M), 48-week results: a randomised, multicentre, open-label, phase 3b, non-inferiority study. Lancet 396, 1994–2005 (2021).
Caskey, M., Klein, F. & Nussenzweig, M. C. Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic. Nat. Med. 25, 547–553 (2019).
Haynes, B. F., Burton, D. R. & Mascola, J. R. Multiple roles for HIV broadly neutralizing antibodies. Sci. Transl. Med. 11, eaaz2686 (2019).
Gama, L. & Koup, R. A. New-generation high-potency and designer antibodies: role in HIV-1 treatment. Annu. Rev. Med. 69, 409–419 (2018).
Nishimura, Y. & Martin, M. A. Of mice, macaques, and men: broadly neutralizing antibody immunotherapy for HIV-1. Cell Host Microbe 22, 207–216 (2017).
Bar, K. J. et al. Effect of HIV antibody VRC01 on viral rebound after treatment interruption. N. Engl. J. Med. 375, 2037–2050 (2016).
Caskey, M. et al. Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117. Nature 522, 487–491 (2015).
Caskey, M. et al. Antibody 10-1074 suppresses viremia in HIV-1-infected individuals. Nat. Med. 23, 185–191 (2017).
Scheid, J. F. et al. HIV-1 antibody 3BNC117 suppresses viral rebound in humans during treatment interruption. Nature 535, 556–560 (2016).
Mendoza, P. et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature 561, 479–484 (2018).
Nishimura, Y. et al. Early antibody therapy can induce long-lasting immunity to SHIV. Nature 543, 559–563 (2017).
Lu, C. L. et al. Enhanced clearance of HIV-1-infected cells by broadly neutralizing antibodies against HIV-1 in vivo. Science 352, 1001–1004 (2016).
Schoofs, T. et al. HIV-1 therapy with monoclonal antibody 3BNC117 elicits host immune responses against HIV-1. Science 352, 997–1001 (2016).
Niessl, J. et al. Combination anti-HIV-1 antibody therapy is associated with increased virus-specific T cell immunity. Nat. Med. 26, 222–227 (2020).
Sneller, M. C. et al. A randomized controlled safety/efficacy trial of therapeutic vaccination in HIV-infected individuals who initiated antiretroviral therapy early in infection. Sci. Transl. Med. 9, eaan8848 (2017).
Deeks, S. G. HIV infection, inflammation, immunosenescence, and aging. Annu. Rev. Med. 62, 141–155 (2011).
Van Gassen, S. et al. FlowSOM: using self-organizing maps for visualization and interpretation of cytometry data. Cytometry A 87, 636–645 (2015).
Nishimura, Y. et al. Immunotherapy during the acute SHIV infection of macaques confers long-term suppression of viremia. J. Exp. Med. 218, e20201214 (2021).
Barouch, D. H. & Deeks, S. G. Immunologic strategies for HIV-1 remission and eradication. Science 345, 169–174 (2014).
Collins, D. R., Gaiha, G. D. & Walker, B. D. CD8+ T cells in HIV control, cure and prevention. Nat. Rev. Immunol. 20, 471–482 (2020).
Kwong, P. D. & Mascola, J. R. HIV-1 vaccines based on antibody identification, B cell ontogeny, and epitope structure. Immunity 48, 855–871 (2018).
Sok, D. & Burton, D. R. Recent progress in broadly neutralizing antibodies to HIV. Nat. Immunol. 19, 1179–1188 (2018).
Collins, D. R. et al. Functional impairment of HIV-specific CD8+ T cells precedes aborted spontaneous control of viremia. Immunity 54, 2372–2384 (2021).
Migueles, S. A. & Connors, M. Success and failure of the cellular immune response against HIV-1. Nat. Immunol. 16, 563–570 (2015).
Sarzotti-Kelsoe, M. et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 409, 131–146 (2014).
Clarridge, K. E. et al. Effect of analytical treatment interruption and reinitiation of antiretroviral therapy on HIV reservoirs and immunologic parameters in infected individuals. PLoS Pathog. 14, e1006792 (2018).
Bruner, K. M. et al. A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nature 566, 120–125 (2019).
Myers, L. E., McQuay, L. J. & Hollinger, F. B. Dilution assay statistics. J. Clin. Microbiol. 32, 732–739 (1994).
Robins, H. S. et al. Comprehensive assessment of T-cell receptor β-chain diversity in αβ T cells. Blood 114, 4099–4107 (2009).
Carlson, C. S. et al. Using synthetic templates to design an unbiased multiplex PCR assay. Nat. Commun. 4, 2680 (2013).
Snyder, T. M. et al. Magnitude and dynamics of the T-cell response to SARS-CoV-2 infection at both individual and population levels. Preprint at MedRxiv https://doi.org/10.1101/2020.07.31.20165647 (2020).
ImmunoMind Team. immunarch: an R package for painless bioinformatics analysis of T-cell and B-cell immune repertoires (version 0.6.7) (Zenodo, 2019).
Vita, R. et al. The immune epitope database (IEDB): 2018 update. Nucleic Acids Res. 47, D339–D343 (2019).
Shugay, M. et al. VDJdb: a curated database of T-cell receptor sequences with known antigen specificity. Nucleic Acids Res. 46, D419–D427 (2018).
Tickotsky, N., Sagiv, T., Prilusky, J., Shifrut, E. & Friedman, N. McPAS-TCR: a manually curated catalogue of pathology-associated T cell receptor sequences. Bioinformatics 33, 2924–2929 (2017).
Zhang, W. et al. PIRD: pan immune repertoire database. Bioinformatics 36, 897–903 (2020).
Kassambara, A. ggpubr: ‘ggplot2’ based publication ready plots. R package version 0.1.7 (2018).
Lê, S., Josse, J. & Husson, F. FactoMineR: an R package for multivariate analysis. J. Stat. Softw. 25, 1–18 (2008).
Acknowledgements
We thank the study volunteers for their participation in this study; D. Asmuth, J. Mascola and S. Read for their guidance; and the NIAID HIV Outpatient Clinic staff for their assistance in the execution of this study. This work was supported by the Intramural Research Program of the National Institute of Allergy and Infectious Diseases, National Institutes of Health.
Author information
Authors and Affiliations
Contributions
M.C.S. and T.-W.C. designed the clinical trial and research. J.B., J.S.J., V.S., B.D.K., E.J.W., R.F.S. M.S.S., S.M. and T.-W.C. performed experiments. J.B. and C.O. performed bioinformatic analysis. M.C.S., K.G., J.T., G.M., E.B., C.K. and T.-W.C. contributed to recruitment of study participants. M.C. and M.C.N. provided study drugs. M.C.S., J.B., M.A.P., C.O., M.S.S, S.M. and T.-W.C. analysed data. M.C.S., A.S.F., S.M. and T.-W.C. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
M.C.N. is listed as an inventor for patents on 3BNC117 (PTC/US2012/038400) and 10-1074 (PTC/US2013/065696); 3BNC117 and 10-1074 are licensed to Gilead Sciences by Rockefeller University from which M.C.N. has received payments. M.C.N. is a member of the Scientific Advisory Boards of Celldex Therapeutics, Walking Fish Therapeutics and Frontier Biotechnologies. M.C.N. had no control over the direction and ultimately the reporting of the clinical portion of the research while holding their financial interests. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks Lu Zheng and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Consolidated Standards of Reporting Trials (CONSORT) flow diagram for the trial.
CONSORT diagram shows the study enrolment of 14 participants who underwent randomization to the bNAb or placebo groups.
Extended Data Fig. 2 Dynamics of HIV reservoirs.
a. Frequencies of CD4+ T cells carrying total HIV DNA in study participants in the placebo arm of Group 1. b. Frequencies of CD4+ T cells carrying total HIV DNA in study participants in the Group 2 in whom plasma viraemia was suppressed by the combination bNAbs.
Extended Data Fig. 3 Longitudinal measurements of CD4+ T cell counts and phenotypic analyses of CD8+ T cells.
a. Levels of CD4+ T cell counts of the bNAb (n = 7) and placebo (n = 7) arms of Group 1 and Group 2 (n = 5) study participants are shown. b. Frequencies of the activation/exhaustion markers TIGIT, PD-1, CD38 and HLA-DR (left) and T cell subsets (TN, naive; TCM, central memory; TTM, transitional memory; TEM, effector memory; TTD, terminally differentiated) on CD8+ T cells of the bNAb (n = 5) and placebo (n = 7) arms of Group 1 and Group 2 (n = 5) study participants are shown. The grey lines indicate median values. P values were determined using the two-sided Wilcoxon matched-pairs signed rank test and were adjusted for multiple testing. ns, not significant.
Extended Data Fig. 4 Phenotypic analysis of T cells.
Longitudinal high-dimensional flow cytometric analyses of PBMCs of study participants. a. Global opt-SNE plots of CD3+ T cells of combined data from each group of study participants. b. Opt-SNE visualization of expression of the indicated markers are shown. c. Opt-SNE map of T cell clusters identified by FlowSOM clustering. Each number indicates a distinct cluster. Heatmap shows the level of expression (MFI) within individual clusters. d. Comparison of frequencies of T cells expressing markers associated with indicated clusters in the bNAb (n = 5) and placebo (n = 7) arms of Group 1 and Group 2 (n = 5) study participants are shown. P values were determined using the two-sided Wilcoxon matched-pairs signed rank test and were adjusted for multiple testing. ns, not significant.
Extended Data Fig. 5 Levels of biomarkers in the plasma of the bNAb (n = 5) and placebo (n = 7) arms of Group 1 and Group 2 (n = 5) study participants over time.
The grey lines indicate median values. P values were determined using the two-sided Wilcoxon matched-pairs signed rank test and were adjusted for multiple testing. ns, not significant.
Extended Data Fig. 6 Analysis of HIV-specific CD8+ T cells.
Frequencies of HIV Gag-specific CD8+ T cells and dynamics of CD8+ T cell receptor (TCR) repertoire. a. Frequencies of polyfunctional (IFN-γ+TNF-α+MIP-1β+) HIV Gag-specific CD8+ T cells in the bNAb (n = 5) and placebo (n = 7) arms of Group 1 and Group 2 (n = 5) study participants are shown. The grey lines indicate median values. P values were determined using the two-sided Wilcoxon matched-pairs signed rank test. b. Changes in the HIV-specific breadth and depth of CD8+ T cells of study participants are shown (upper panels). Highly enriched CD8+ T cells were obtained using a bead-based purification method. The analysis includes 35 CD8+ T cell-derived genomic DNA samples from 12 study participants (15 samples from 5 participants in the bNAb arm of Group 1, 5 samples from 2 participants in the placebo arm of Group 1, and 15 samples from 5 participants in Group 2). Violin plots show the Gaussian kernel probability density of the breadth/depth values over time. The median values and interquartile ranges of the time point-specific distribution are shown as circles and vertical lines, respectively. Principal component analysis (PCA) of the changes in the TCR repertoire characteristics is shown (lower panels). Each ellipse shows the 95% confidence interval in the PCA space and the center of each ellipse is indicated by larger sized symbols that represent specific time points. Lower left panels depict PCA results with the frequencies of the HIV-specific clonotypes ranked among the top 25 with respect to their P values associated with the pairwise comparisons between the three time points. Lower right panels depict PCA results with the gene usage profiles derived from the TRBV-TRBJ gene pairs in the above clonotypes. Principal component (PC) 1 and PC2 represent a lower-dimensional representation of the input data consisting of the frequencies of the HIV-specific clonotypes (lower left panel) and the usage levels of the TRBV-TRBJ gene pairs (lower right panel) for each patient group. P values were determined using the two-sided Wilcoxon signed-rank test.
Supplementary information
Supplementary Information
Supplementary Tables 1–5 and Supplementary Fig.1, containing the gating strategy for flow cytometry analysis.
Rights and permissions
About this article

Cite this article
Sneller, M.C., Blazkova, J., Justement, J.S. et al. Combination anti-HIV antibodies provide sustained virological suppression. Nature 606, 375–381 (2022). https://doi.org/10.1038/s41586-022-04797-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-04797-9
This article is cited by
-
Immune targeting of HIV-1 reservoir cells: a path to elimination strategies and cure
Nature Reviews Microbiology (2024)
-
Predicted resistance to broadly neutralizing antibodies (bnAbs) and associated HIV-1 envelope characteristics among seroconverting adults in Botswana
Scientific Reports (2023)
-
Structures and immune recognition of Env trimers from two Asia prevalent HIV-1 CRFs
Nature Communications (2023)
-
Prevention, treatment and cure of HIV infection
Nature Reviews Microbiology (2023)
-
High monoclonal neutralization titers reduced breakthrough HIV-1 viral loads in the Antibody Mediated Prevention trials
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
