
Abstract
The role of microbiota in health and diseases is being highlighted by numerous studies since its discovery. Depending on the localized regions, microbiota can be classified into gut, oral, respiratory, and skin microbiota. The microbial communities are in symbiosis with the host, contributing to homeostasis and regulating immune function. However, microbiota dysbiosis can lead to dysregulation of bodily functions and diseases including cardiovascular diseases (CVDs), cancers, respiratory diseases, etc. In this review, we discuss the current knowledge of how microbiota links to host health or pathogenesis. We first summarize the research of microbiota in healthy conditions, including the gut-brain axis, colonization resistance and immune modulation. Then, we highlight the pathogenesis of microbiota dysbiosis in disease development and progression, primarily associated with dysregulation of community composition, modulation of host immune response, and induction of chronic inflammation. Finally, we introduce the clinical approaches that utilize microbiota for disease treatment, such as microbiota modulation and fecal microbial transplantation.
Similar content being viewed by others

Introduction
The origin of “microbiota” can be dated back to early 1900s. It was found that a vast number of microorganisms, including bacteria, yeasts, and viruses, coexist in various sites of the human body (gut, skin, lung, oral cavity).1 In addition, the human microbiota, also known as “the hidden organ,” contribute over 150 times more genetic information than that of the entire human genome.2 Although “microbiota” and “microbiome” are often interchangeable, there are certain differences between the two terms. Microbiota describes the living microorganisms found in a defined environment, such as oral and gut microbiota. Microbiome refers to the collection of genomes from all the microorganisms in the environment, which includes not only the community of the microorganisms, but also the microbial structural elements, metabolites, and the environmental conditions.3 In this regard, microbiome encompasses a broader spectrum than that of microbiota. In the current review, we mainly focus on the function of microbiota in human health and diseases.
The composition of microbiota varies from site to site (depicted in Fig. 1). Gut microbiota is considered the most significant one in maintaining our health.4 The gut bacteria serve several functions, such as fermentation of food, protection against pathogens, stimulating immune response, and vitamin production.5 Generally, the gut microbiota is composed of 6 phyla including Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia, among which Firmicutes and Bacteroidetes are the major types.6 The most studied fungi (gut mycobiota) are Candida, Saccharomyces, Malassezia, and Cladosporium.7 In addition to bacteria and fungi, the human gut microbiota also contain viruses, phages, and archaea, mainly M. smithii.8

Human microbiota composition in different locations. Predominant bacterial genera in the oral cavity, respiratory tract, skin, gut, and vagina are highlighted
While less well established compared with gut, microbiota is also localized in other regions including the oral cavity, lung, vagina, and skin. Oral microbiota is considered the second largest microbial community in human.9 The oral cavity can be further divided into multiple habitats of microbiota, including saliva, tongue, tooth surfaces, gums, buccal mucosa, palate, and subgingival/supragingival plaque, which may exhibit substantial and rapid changes in composition and activity, owing to the factors such as changes in pH, gene mutations, and interactions among the bacteria.10 The microbiota composition in all seven sites shares overall similarities but with small scale differences. In general, the major bacteria present in oral microbiota are Firmicutes, Proteobacteria, Bacteroidetes, Actinobacteria, and Fusobacteria.
Although healthy human lungs were long considered sterile, numerous studies have demonstrated that microbiota is also present in lung tissues.11 The core lung microbiota included Actinobacteria, Bacteroidetes, Firmicutes, and Proteobacteria. The composition of lung microbiota is primarily determined by three factors: 1) microbial immigration, 2) the elimination of microorganisms, and 3) the reproduction rates of microorganisms.12
In human skin, the distribution and variety of glands and hair follicles vary among each geographic region. The physical and chemical differences of skin regions create distinct composition of microbiota.13 Generally, the skin microbiota is composed of Actinobacteria, Bacteroidetes, Cyanobacteria, Firmicutes, and Proteobacteria.
In recent decades, tremendous amount of work has highlighted the relationship between microbiota and diseases such as cancers, diabetes, and neurological disorders. Moreover, manipulating microbiota in human body can be key for disease treatment. Here, we summarize and discuss the current state of knowledge of human microbiota in development of diseases, mediating health conditions, and the potential clinical application in disease treatments.
Microbiota in health
The “healthy” gut microbiota
Intestinal microbial balance is closely relevant to human diseases and health. Compared with other regions of the body, the human gastrointestinal (GI) tract contains an abundant microbial community which gathers ~100 trillion microorganisms.14 Extensive studies have been performed to reveal the important relationship between gut microbiota and basic human biological processes. For example, current advances have shown that human microbiota is closely involved in nutrient extraction, metabolism, and immunity.15 Microbiota may affect biological processes via several mechanisms. For energy and nutrient extraction from food, microbiota plays crucial roles due to the versatile metabolic genes which provide independent unique enzymes and biochemical pathways.16 Moreover, the biosynthesis of bioactive molecules such as vitamins, amino acids and lipids, are also highly dependent on the gut microbiota.17 Regarding the immune system, the human microbiota not only protects the host from external pathogens by producing antimicrobial substances but also serves as a significant component in the development of intestinal mucosa and immune system.
In healthy conditions, the gut microbiota exhibits stability, resilience, and symbiotic interaction with the host. There is a lot of research into the definition of a “healthy” gut microbiota and its link to host physiological functions. Gut microbiota is composed of bacteria, yeasts, and viruses. A healthy microbiota community often demonstrates high taxonomic diversity, high microbial gene richness and stable core microbiota.18 However, it should be noted that the relative distribution of microorganisms is unique between individuals and may undergo variations within the same individual. In human, gut microbiota may vary due to age and environmental factors (for example, medication usage). Additionally, gut microbiota varies in different anatomical parts of the GI tract. For example, Proteobacteria such as Enterobacteriaceae are found in the small intestine but not the colon. Instead, Bacteriodetes such as Bacteroidaceae, Prevotellaceae and Rikenellaceae are often found in the colon.19 Such variations are majorly due to the different environments. In the small intestine, the transit time is short and bile concentration is high, while in the colon, which has slower flow rates and milder pH, as well as larger microbial communities, especially anaerobic types, are commonly observed.20 Besides spatial distribution, gut microbiota also differs by age. Generally, the microbiota diversity increases in the time between childhood and adulthood and decreases at older age (over 70).21 Before the formation of a relatively stable gut microbiota composition, the diversity of children’s microbiota is dominated by Akkermansia muciniphila, Bacteroides, Veillonella, Clostridium coccoides spp., and Clostridium botulinum spp.22 At about age 3, children’s gut microbiota becomes comparable to that of adults, with three major microbial phyla including Firmicutes, Bacteroidetes and Actinobacteria becoming dominant.23 Subsequently at older age, dietary and immune system change potentially affect the composition of the human gut microbiota. Specifically, elder people tend to exhibit decreased Bifidobacterium and increased Clostridium and Proteobacteria.24 The decrease in the anaerobic bacteria Bifidobacterium is considered relevant to deteriorated inflammatory status due to its role in stimulating the immune system. Since the microbiota plays an important role in human well-being, also proactively involves in multiple biological processes and disease development, the research on human microbiota is going beyond compositional studies and investigation on members’ associations. Specifically, more attention has been paid on explaining the causality of microbiota functions, especially with the boom of new techniques of high-throughput sequencing, microbiota interactive modeling and simulation. Overall, further investigations are still necessary to unveil the roles of human microbiota, in order to support the development of microbiome-based diagnosis and personalized medicine (Table 1).
Rodent models for human microbiota research
The human microbiota has attracted more and more research in recent decades. However, the studies of local microbiota require invasive sampling methods, with practical or ethical reasons in concern. Animal models, particularly mouse and rat models, have also been used to study the pathogenic and therapeutic potential of microbiota with varies diseases.25 With a majority of microbiota research is focusing on gut microbiota, the use of germ-free (GF) mouse model has become popular due to its translatability. It should be noted that, in order to translate such generated knowledge from rodent to human, the similarities and differences between their microbiota profile need to be considered. In Table 2, we summarized some commonly used rodent models and their role in microbiota research.
The genome data showed that more than 85% of the genomic sequences between human and mouse are conserved, while the main difference is found in the primary sequence of regulatory elements. Cheng et al. reported that, in murine genome, half of the transcription factor binding sites may not have orthologous sequences in human genome.26 Moreover, the genomic studies have shown a significant difference in the immune system and its regulation in different species. Since gut microbiota has major impact to host innate and adaptive immune responses, the translation of findings from rodents to human should be carefully validated before drawing definite conclusions. While human and murine gut microbiota has 90% overlapping in phyla and genera levels, the composition and abundance of microbes have key discrepancies.27 For instance, the major difference is the Firmicutes/Bacteroidetes ratio, where it is significantly higher in human than mice. Particularly, human Bacteroidetes is mainly composed of Prevotellaceae and Bacteroidaceae, while mice Bacteroidetes are primarily composed of S24-7. Regarding the Firmicutes, Ruminococcaceae is the major phylum observed in human and Clostridiales is the major one observed in mice. Moreover, human and mouse each carries specific genera, such as Faecalibacterium, Megasphera, Asteroleplasma, Succinivibrio, Paraprevotella in human and Mucispirillum in mouse.28
Colonization resistance
Humans are born with and form a large community of symbiotic and pathogenic microbes, which inhabit our gut, skin, mucosal passages, and form a stable community that is resistant to external pathogens. The term “colonization resistance” was initially coined in the 1950s when Bohnhoff et al. found that mice became significantly sensitive to a specific type of bacterial infection after antibiotic treatment.29 Later, such conclusion was further applied to the phenomenon that current microbiota could provide resistance the colonization of invading pathogenic species, also from which researchers recognize. As a result, the microbiota is crucial shield in protecting us from exogenous microorganisms. Despite the fact that microbiota colonization resistance has not been fully elucidated, with the advent of GF animal models, researchers have discovered several potential mechanisms such as nutrient competition,30 antimicrobial production, and bacteriophage deployment. Another example of colonization resistance is the interaction of symbolic and pathogenic E. coli., where indigenous E. coli strains compete with pathogenic E. coli O157:H7 for the amino acid proline in consuming nutrients.In this section, we focus on the gut microbiota and colonization resistance. The vaginal and skin microbiota and their colonization resistance are also discussed.
The GI tract digests proteins as well as sugars from foods. Metabolizing polysaccharides and specific proteins requires multiple enzymes produced by various bacteria. For example, Bacteroides species in the large intestine are responsible for sugar harvest.31 Pathogenic Enterobacteriaceae also utilizes sugar and amino acids in gut.32 Freter et al. proposed a niche hypothesis which has been supported by in vitro and in vivo studies. The hypothesis states that the composition and abundance of gut microbiota is determined by one or a few nutritional substrates.33 In mouse models, when a single type of sugar is removed, both the microbiota composition and the ability of resistance to pathobiont were altered.34
Probably due to the necessity of competing with foreign bacteria, gut bacteria have developed various ways of suppressing competitors, including the secretion of diverse bacteriocins. A contact-dependent competition in the gut, namely type 6 secretion system, was originally identified in the bacteria secretion system involved with eukaryotic cells,35 which was later found relevant to intraspecies killing. The system works by contact cells delivering effectors, such as degraders of nucleotide, cell walls and membranes, into the cytoplasm.36 Moreover, this system may also contribute to the abundance of Bacteroides species in the mouse and human gut.37 Besides the type 6 system, other systems such as type 7 (or ESX system), also mediate the intra- and interspecies killing.38 Currently, the contact-dependent systems of gut microbiota inhibition and growth are being increasingly discovered. The intermediate genes, immunity and effectors may serve as amenable factors which are modifiable via bioengineering methods. Additionally, they are valuable for studying the interactions, structure, and dynamics of the gut microbiota. However, the exact role of bactericidal mechanisms remain poorly understood and further studies are still necessary.
Bacteriophage deployment is another mechanism of colonization resistance in the gut; however relevant research is still in an immature stage.32 It has been revealed that two cycles, namely the lytic cycle and the lysogenic cycle, are involved in bacteriophage infection. Phages duplicate by injecting genomic segment into the bacterial cytoplasm, after which the two cycles start to branch. Phages in lysogenic stage insert their genome into bacteria genome and render prophages, which guarantees the replication of phage DNA and entrance into the lytic cycle. In the lytic cycle, phage DNA starts replication, modification and expression, resulting in new phage assembly, cell lysis and phage spreading.39 There are several potential mechanisms to prevent bacteriophage infection including the blockage of surface receptor recognition, superinfection exclusion system and abortive infection. For infection prevention, resistant strains exhibit compositions similar to bacterial surface and thereby could serve as decoys for attacking phages.40 DNA replication could be prevented by “restriction-modification” system, mainly by methyltransferase and restriction endonucleases. This system serves as the primitive inner bacteria defense system in the human body, despite the disadvantage of this system that it also damages the host DNA.41 The defense system of bacteria also inspired the discovery of Clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9, which has been extensively reviewed.42 Later, newer defense systems, such as bacteriophage exclusion have been discovered to work by preventing DNA replication.43 The third potential mechanism is the abortive infection, where the infected cells are killed, and surrounding ones are protected. This mechanism is not yet fully elucidated, and still needs further exploration.
Besides the gut, the vaginal microbiota also plays crucial in resisting the colonization of invading pathogenic microbiomes, which is important for preventing sexually transmitted infections, urinary tract infections and vulvovaginal candidiasis.44 Traditionally, the cultivation methods suggested the vaginal microbiota as a community that lacks species that produce lactic acid (e.g., Lactobacillus species).45 Moreover, the vaginal microbial community is overabundant with anaerobic bacteria including Gardnerella vaginalis, Prevotella spp., Mobiluncus spp., Ureaplasma urealyticum, and Mycoplasma hominis.46 Later studies identified Lactobacilli as important members of vaginal microbiota. To better understand the vaginal microbiota, researchers have grouped the vaginal bacteria community into five types known as community state types (CSTs) I–V. All five communities are dominated by L. crispatus, L. gasseri, L. iners, polymicrobial flora including Lactobacillus and bacterial vaginosis-associated bacteria (BVAB), and L. jensenii. The CST I, III and IV are commonly found in women and have been extensively studied, while the other two types are rare.47 The Lactobacillus species are believed to provide protective functions by generating bactericidal and virucidal agents, including lactic acid and bacteriocins.48 As a result, the vaginal Lactobacilli is considered a risk factor of sexually transmitted infections such as human immunodeficiency virus (HIV),49 human papillomavirus,50 and herpes simplex virus infections.51 In a previous randomized clinical study, Schwebke et al. found that women treated with atypical gram positive stain smears showed lower risk for incident chlamydial genital infection.52 In another study which involves 3620 nonpregnant women, Brotman et al. found a strong association between bacterial vaginosis and elevated risk of genital infection.53 So far, only limited studies are available regarding vaginal colonization resistance, but it has been widely agreed upon that the vaginal colonization resistance plays crucial protective roles in preventing pathogenic infections.
The skin, as the largest organ in human body, is colonized by dense microbiome communities. Healthy skin with balanced microbiota is believed to contribute to colonization resistance against pathogenic infections. Changes in the skin microbiota (dybiosis) are highly associated with many common skin diseases, such as acne, a chronic inflammatory skin condition mediated by Propionibacterium acnes.54 Severity of P. acnes pathophysiology is correlated with the level of sebum secretion. As a result, acne is prevalent in teenager and a minor portion of adults. Also, the production of bacteriocins by current residing microbiome provides further protection against invading species.55 For example, S. epidermidis was suggested to destroy S. aureus biofilms via a serine protease.56 In addition, S. lugdunensis was discovered to produce lugdunin, an inhibitor of nasal colonization with S. aureus. Lugdunin also inhibits other pathogens including Enterococcus faecalis, Listeria monocytogenes, Streptococcus pneumoniae, and Pseudomonas aeruginosa.57 Overall, understanding the interactions among skin microbiota communities will be beneficial to the control of skin diseases or disorders.
The microbiota–gut–brain axis
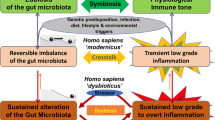
In the 1980s, with the development of brain imaging, our understanding of the critical roles of the gut–brain axis in homeostasis was established.58 Researchers then reached consensus that this axis is bidirectional. On the one hand, gut distension activates key pathways within the brain, while on the other hand, such pathways are involved with gut disorders, for example irritable bowel syndrome (IBS).59 In the past decades, gut microbiota was identified as a key regulator of the gut–brain axis. Multiple animal models as well as human studies have been used to model the gut-brain axis. The factors contributing to gut–brain axis balance are summarized in Fig. 2. A recent study by Chen et al.60 found that, due to the loss of histone demethylases (eg, KDM5), fruit flies (Drosophila melanogaster) showed intestinal barrier dysfunction and change in social behaviors such as mating. This is one of the direct pieces of evidence that mating behavior is likely relevant to the enteric bacteria. Similarly, in mouse models, Bravo et al. performed chronic feeding with lactic acid bacteria Lactobacillus rhamnosus on mice and found region-dependent alterations in the brain such as GABA gene upregulation in cortical regions and downregulation in the hippocampus, amygdala, and locus coeruleus.61 Thus, it indicates that gut microbiota could influence neurophysiology and behavior. Moreover, Buffington et al.62 reported that maternal high-fat diet induces gut microbiota shifts and physiological change in the offspring brain, such as fewer oxytocin immunoreactive neurons in the hypothalamus. Additionally, offspring social deficits and gut microbiota shifts could be prevented by co-housing with offspring of regular-diet mothers.62 This finding further supports that gut microbiota negatively impacts offspring social behavior. Gut microbiota also affects the wound-healing process. Mice fed with lactic acid bacteria Lactobacillus reuteri showed enhanced wound-healing properties via upregulation of oxytocin, which is a regulatory factor that activates host CD4 + Foxp3 + CD25 + immune T regulatory cells.63 Other studies also showed that gut microbiota impacts cognition, anxiety, depression-related behavior, and reward/addiction pathways of mice.64 Studies in chimpanzees (Pan troglodytes) revealed the other direction of microbiota in the gut-brain axis: composition of gut microbiota is impacted by various social interactions.65 Studies of gut–brain axis in humans showed similar results regarding the connection between brain physiology and gut microbial ecology. In 2016, Allen et al. performed a preclinical study on healthy volunteers to test if psychobiotic consumption could affect neurophysiological responses such as stress response, cognition, and brain activity.66 Results indicated that consumption of B. longum 1714 is associated with reduced stress and improved memory. However, in this study, the number of samples was small (N = 11) and confounding factors such as the environment, diet, lifestyles, and genetic variations were not fully considered. In another study using mouse models, gut microbiota was discovered to be necessary for motor deficits, microglia activation, and αSyn pathology.67 The authors transplanted the microbiota from Parkinson’s disease patients and found that the mice showed enhanced physical impairments compared with mice with microbiota from healthy donors. Thus, it suggests that gut microbes are potentially relevant to neurodegenerative diseases such as Parkinson’s disease and could be used as a therapeutic marker. Furthermore, researchers found there is significant difference in the component of microbes in the gut of children with and without autism spectrum disorders, a pervasive developmental disorder characterized by social abnormalities, communication impairments, and repetitive behaviors.68 This is indeed another evidence showing the relationship between GI microbiota and neurophysiology.

Bidirectional gut-brain axis interactions and the common factors contributing to the gut–brain activity
Many pathways have been proposed to mediate the communication within the gut-brain-axis. The signal passage along gut-brain-axis involves the interactions among autonomic nervous system (ANS), enteric neural system (ENS), central nervous system (CNS), immune system, and endocrine system. The ANS, which controls GI tract functions such as gut movement and mucus production, is a complex network that integrates the communication between the gut and the brain, as well as induces CNS effects in the gut since CNS is responsible for processing the visceral information.69 The ANS directly triggers neurological responses in the gut which further causes physiological changes. The ANS also mediates the interaction between the gut microbiota and ENS. ANS-triggered ENS activity results in the absorption and delivery of pre-/probiotics in the GI tract such as starches and other microbial nutrients.70 Microbes could affect the neural system via neuromodulatory metabolites including tryptophan, serotonins, GABA and catecholamines.58 Previous study on mice models have proved that gut microbial metabolite 4-ethylphenylsulfate induces mental disorders (such as anxiety-like behavior).71 Additionally, the gut microbial tryptophan metabolite indole was found relevant to the activation of the vagus nerve, the 10th cranial nerve that connects the gut and brain.72 In this study, rats with acute and high indole overproduction showed decreased motor activity, while rats with chronic and moderate indole increase showed enhanced anxiety-like behavior. Similarly, the bacteria Lactobacillus rhamnosus was found to induce information transmission in vagal afferents in the mesenteric nerve bundle. Such induction could be eliminated by vagotomy.73 Also, treatment with bacteria Lactobacillus reuteri in rats models was found to help mice with social deficits; such change was also restored in mice with vagotomy.74 Recent studies also reported potential mechanisms of microbiota–ENS interaction. As one of the major serotonin producers in human, gut microbiota is linked to ENS activation by 5-HT receptors. De Vadder et al. demonstrated the interaction by pharmacological modulation of 5-HT receptors and depletion of endogenous 5-HT.75 The presence of 5-HT receptor antagonist negatively affects ENS activity. The gut microbiota also communicates with another major neuroendocrine system, the hypothalamic–pituitary–adrenal (HPA) axis, which is known to coordinate stress response.76 Signal molecules generated in HPA are distributed throughout the body and affect gut microbiota. To illustrate the connection between HPA and gut microbiota, GF mice are used. Studies revealed that GF mice exhibited elevated plasma corticosterone, indicating hyperresponsive HPA axis and the regulatory effect of gut microbiota.77 In human, it has been reported that bowel syndrome patients (with gut microbiota changes) tend to have exaggerated adrenocorticotrophic hormone response to corticotrophin releasing factor infusion.78 Despite there have been numerous of studies on the bidirectional pathways between the gut-microbiota and the brain, it is still difficult to fully understand the mechanisms.
Numerous influencing internal and external factors have been discovered to modulate the gut-brain axis of the host, including genetics, socioeconomic status, diet, medications, and environmental factors.79 Genetics and epigenetics are important in understanding the brain as well as gut health. An increasing number of studies have been performed on the relationship between host (human or mice) and microbiota genetics. One of the important components of microbiota-host genetic interaction is via the modulation of RNAs. For example, in GF mice models, researchers found that microRNAs were dysregulated in GF mice in certain brain regions, amygdala and prefrontal cortex, which suggests a close relationship between gut microbiota and brain physiology.80 In another study of gut microbiota and hippocampal RNAs using GF mice, Chen et al. found that gut microbiota significantly regulates the expression level of hippocampal microRNAs and mRNAs. Specifically, re-colonizing the gut microbiota in GF mice did not reverse the behavioral change such as less latency to familiar food, but microRNAs and mRNAs were significantly restored.81
As mentioned before, lifestyles, especially diet, have been shown to be among the most critical factors in modulating the gut-brain axis. For example, a high-fat diet with only animal products will shift the microbiota composition profoundly. Specifically, animal models with high-fat diet showed reduction in Bacteroidetes levels and an increase in both Proteobacteria and Firmicute levels.82,83 Proteobacteria (Bilophila wadsworthia) abundance was also observed in another study of high-fat diet-fed animals.84 On the contrary, the Mediterranean diet composed of whole grains, nuts, vegetables, fruits, and only certain animal products (fish and poultry) showed beneficial results in hosts. In human intervention studies of diet, consumption of the Mediterranean diet has been shown to significantly reduce the occurrence of neurovegetative disorders, psychiatric conditions, cancer, and cardiovascular diseases.85 Mediterranean diet also showed correlation with reduced risk of depression.86 Though strong evidence showed that the Mediterranean diet is beneficial to the hosts, further mechanism studies are still necessary to illustrate the regulatory mechanism of such diet on the gut–brain axis. Another type of diet with high fat and low carbohydrate, namely the ketogenic diet, is popular because it forces the consumption of the body’s reserved fat. Ketogenic diet was considered to be able to inhibit apoptosis in neurodegenerative diseases because of the increase in serum ketones, which has been shown to improve mitochondrial activity.87 Studies have shown that the ketogenic diet also causes shift in microbiota abundance in the gut. Specifically, Akkermansia, Parabacteroides, Sutterella, and Erysipelotrichaceae levels were significantly higher in mice on ketogenic diets.88 Moreover, mice on ketogenic diets were better protected from acute epileptogenic seizures compared with the control group on a normal diet. Furthermore, colonization with increased microbiomes in GF mice also showed correlation with seizure protection, as well as alterations in hippocampal metabolomic profiles. All above studies support the conclusion that changes in lifestyles have marked impacts on the gut microbiota.
Finally, medications, especially antibiotics, will directly affect the gut microbiota and subsequently the gut–brain axis. Besides antibiotics, a growing number of studies also showed that nonantibiotic drugs can change the gut–microbiota composition, as well as neurophysiology and behavior.89 In a large-scale gut-microbiota project named the Belgian Flemish Gut Flora Project, antibiotics, osmotic laxatives, hormones, benzodiazepines, antidepressants, antihistamines, and inflammatory bowel disease drugs were found to be highly relevant to the variation of gut microbiota.90 Other studies also showed that proton pump inhibitors, metformin, and statins can impact gut microbiota.91 Moreover, due to the rise of interest in the gut–brain axis, more and more psychotropic medications were discovered to have antimicrobial activities. Examples are serotonin antagonists such as sertraline, paroxetine, and fluoxetine, which have antimicrobial activity against gram-positive bacteria such as Staphylococcus and Enterococcus.92 These findings indicate the potential impact of medications on the gut–brain axis.
Microbiota in the development of immune systems
Microbiota in different organs exhibits distinct characteristics and compositions. As a result, microbiota interacts with multiple biological processes of the host. In this section, we introduce the interactions between human microbiota in gut, oral cavities, lungs, skin, vagina, and the development of immune systems.
The human immune system is comprised of innate and adaptive immune responses, both of which have been shown to extensively interact with microbiota. The innate immune response has critical role in maintaining a homeostatic environment by eliminating pathogenic bacteria and regulating the adaptive response to microbiota. These effects are mediated by factors such as secretory IgA (sIgA), toll-like receptor 5 (TLR5), autophagy, and inflammasomes.93 For instance, slgA can bind and form complexes with commensal bacteria, which selectively presents the bacterial components to tolerogenic dendritic cells. As an anti-inflammatory molecule, slgA can reduce the inflammatory response that could result from the immense bacteria load in the organs. On the other hand, dysbiosis of microbiota can alter the sIgA response and lead to unregulated bacterial growth. Hapfelmeier et al. showed that microbiota-specific sIgA response was observed in GF mice using reversible microbial colonization system.94 The sIgA induction was confirmed as a gradual response to current bacterial exposure, suggesting a crosstalk between microbiota and immune system. The adaptive immune response is another important part to maintain a healthy microbiota and immune balance. Particularly, the education of adaptive immune response is achieved by differentiation and maturation of B and T cells and establishment of immune tolerance to microbiota.95 Depending on the bacteria species, the CD4 T cell responses vary significantly, which leads to the differentiation into distinct subsets and the subsequent pro-inflammatory cytokine release such as interferon-γ and interleukin IL-17A. The crosstalk between microbiota and adaptive immune response will be further discussed in the following sections.
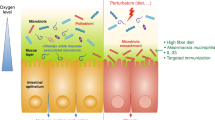
The GI tract hosts a large number of immune cells, which constantly communicate with the gut microbiota. The maturation of the immune system needs the development of commensal microorganism. One of the mechanisms of gut microbiota affecting the immune system is by mediating neutrophil migration, which subsequently impacts T cell differentiation into various types such as helper T cells (Th1, Th2, and Th17) and regulatory T cells.96 Disorders in microbiota development during the maturation of the immune system could lead to deteriorated immunological tolerance and autoimmune diseases.97 Additionally, heterogeneous molecules produced by microbiota may induce immune response and stimulate inflammation or chronic tissue damage.98 The general interactions of microbiota and host immune response during healthy and disease states are depicted in Fig. 3.

Factors affecting microbiota-associated chronic inflammation in healthy and disease state
Human immune system is closely related to the microbiota as a complex symbolic relationship during the co-evolution of vertebrates and microbiota.99 The vertical transmission from the mother’s microbiota to the child at birth is considered the initial introduction of microbiota to the child. As a result, infants born by Cesarean section are colonized with bacteria of the epidermal origin, which might link to higher risk of developing allergies and asthma compared with infants who received initial microbiota from the maternal vaginal flora.100 Such difference in immune system and microbiota would be gradually eliminated during growth. As mentioned before, the infant’s microbiota stabilizes at ~1-year-old and resembles that of adults. The neonatal immune system also rapidly develops under the impact of dynamic microbiota.101 In addition to the microbiota transmission during birth, breastfeeding also plays crucial roles in the establishment of infant immune system as well as microbiota. Besides the required nutrients and antimicrobial proteins, breast milk provides slgA, which is specifically shaped by the maternal microbiota. As a result, infants’ microbiota is seeded not only by the maternal epidermal or virginal origin but reinforced by the sIgA shaped by maternal microbiota. Moreover, before the solidification of infant immune system, the sIgA significantly protects the newborn against pathogens.99 To summarize, the maternal-neonatal microbial bond supports the close relationship between microbiota and the immune system.
The gut microbiota has been closely connected to immunological response due to the fact that enteric microorganisms may promote macromolecules and antigens through the gut epithelium.102 The principal component of bacterial flagellum, namely flagellin, elaborates the relationship between gut epithelial integrity and host immunity. Flagellin is recognized by TLR5, which is found actively expressed in B-cells and CD4 + T-cells. Differentiated B-cells produce IgA, which neutralizes the pathogen and potential subsequent infection.103
The gut microbiota contributes to the development of immune system via the gut-associated lymphoid tissues composed of Peyer’s patches (PPs), plasma cells, and lymphocytes. Previous studies have shown that the gut bacteria interact with mucosal antibodies that are taken up by CD11 + dendritic cells in the PPs. Studies also showed that the luminal microbiota bound to SIgA increased their presence in PPs.104 The CD8 + T cells are mostly found in the intraepithelial intestine compartment, and the microbiota plays important roles in maintaining the function of CD8 + T-cells. This is supported by previous studies showing that GF mice exhibit reduced intestinal CD8 + T-cells.105 In all, understanding the relationship between the microbiota and the immune system is a critically important topic in health sciences. However, due to our innate understanding of the network of gut-immune system, greater attention will be necessary to further promote our knowledge in immune homeostasis and novel immune-microbe therapies.
The oral cavity is another important habitat where the microbiota could colonize. Different from the gut environment, the oral cavity contains both hard surface of teeth and epithelial surface of mucosal membrane. Approximately 50 species (1000 sub-species) exist in the oral microbiota. Due to the constant exposure to saliva, oral microbiota acquired the feature of avid adherence, which guarantees their colonization and resistance to the forces of fluid flow.106 Oral microbiota contains complex polymicrobial communities which have complicated interactions with the host’s diet and immunity. The colonization resistance in oral microbiota is affected by not only the lack of a single treatment for therapeutic intervention, but also due to the presence of a fluid phase which could inactivate bioactive molecules. The number of different oral sites where disease can occur and the poor retention of topical application of therapies are also hurdles to the treatment of oral disease caused by pathogens. Oral pathogens exert the ability to trigger immune response such as pro-inflammatory responses. On the other hand, alterations in host immune system also affects the oral microbiota community. For example, gingivitis, a common disease in humans, is caused by immune-inflammatory responses where neutrophils are recruited to the gingival tissues.107 In periodontal disease, inflammation has been found to be an important driver for the growth of pathogenic microorganisms since inflammation can cause tissue destruction, which provides nutrient to microbiomes.108 However, inflammation could subsequently trigger bactericidal activity of the immune system. Thus, there exists such a paradox in dysbiosis that if the host immune system was downregulated, microbiomes will starve due to lack of nutrients. Periodontitis-associated bacteria such as the P. gingivalis is able to tackle the conundrum by triggering the host immune response without coupling bactericidal activity. Such function has been demonstrated in mice models where P. gingivalis intervened the host-microbiota homeostasis and contributed to the development of periodontitis.109 The special manipulation of the host immune system by P. gingivalis has been revealed to involve C5a receptor 1 (C5aR1) and TLR2. In human and mouse neutrophils, P. gingivalis was able to initialize a C5aR1-TLR2 signal which separates a TLR2-MyD88 pathway from a TLR2-MyD88-Mal-PI3K pathway, leading to inflammation and blocked phagocytosis. In summary, the oral microbiota could be both beneficial by potentially stabilizing the microbial diversity and harmful to cause collective pathogenic outcomes.
Like gut and oral tissues, the lungs also present a complex bacteria community. The lung microbiota is relatively dynamic as a result of the microbiome immigration and elimination via aspiration, cough, or mucociliary clearance.110 The majority of microbes in lungs belong to Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria families.111 The lung microbiota is responsible for the state of immune tolerance that protects the host from undesired inflammatory response.112 This function is mediated by the interaction between commensal bacteria and lung immune cells. Given the important role that lung microbiota plays in maintaining lung homeostasis, the lung microbiota composition is useful in monitoring lung health conditions.113 The interactions between lung microbiota and local immune cells are closely relevant to the pattern recognition receptors (PRRs), which are responsible for the recognition of microbial molecules. The above-mentioned TLRs also belong to PRRs. Activation of PRRs could stimulate the engagement of ligands and further induces immune-related genes expression, which promotes the immune response against pathogens.114 Additionally, the lung microbiota was also found to regulate antigen-presenting cells and regulatory T cells. In mice models, it was found that newborn mice showed excessive airway eosinophilia, Th2 cytokine release, and hyper-responsiveness after exposure to allergens. With the bacterial load increasing during the following two weeks, the microbiota composition was shifted (Gammaproteobacteria and Firmicutes toward Bacteroidetes) and responsiveness to allergens was decreased. The major mechanism includes the appearance of the Helios-regulatory T cells subset, which is promoted by changes in lung commensal bacteria community.115 Furthermore, infant mice without proper lung microbiota would suffer from excessive sensitivity to allergens until adulthood.
The human skin, like gut, is also colonized by a dense community of microbiomes composed of highly diverse communities. It has been discovered that the skin microbiota is composed of prokaryotes (bacteria and archaea) and eukaryotes (fungi, metazoic parasites). Similar to the gut microbiota, skin microbiota is also involved in the development of the innate immune system. For example, S. epidermidis produces lipoteichoic acids which prevent skin from injury-induced inflammation. The potential mechanisms include the inhibition of cytokine release and TLR2-based immune responses.116 Interestingly, S. epidermidis also promotes the expression of certain antimicrobial peptides like human β-defensins (hBDs) which enhances the skin defense.117 Moreover, S. epidermidis was believed to strengthen the function of skin lymphocytes, thereby contributes to increased skin immunity.118 In summary, as a primary part of the human immune system, the skin harbors a wide range of cells that perform functions of immunity such as macrophages, dendritic cells, lymphocytes and various T-cell populations. Moreover, due to the advent of high-throughput sequencing, researchers are able to perform in-depth taxonomic analysis of the skin microbiota, which further boosts our understanding of roles that the skin microbiota plays in human wellness.
As mentioned above, vaginal microbiota is critical in protecting the host from invading pathogens via colonization resistance. It was also revealed that vaginal microbiota drives the innate immune response. Specifically, the vaginal microbiota stimulates the PRRs in and on epithelial cells lining the vagina and upper genital tract and initializes cytokine signaling cascades.119 For example, the release of interleukin (IL)-1β/6/8 and Tumour Necrosis Factor alpha (TNF-α) recruits or activates immune cells like Natural killer (NK) cells, macrophages, CD4 + helper T-cells, CD8 + cytotoxic T-lymphocytes and B-lymphocytes.120 Bacterial vaginosis (BV) is one of the most common vaginal dysbiosis due to the displacement of Lactobacillus spp and the increased concentration of BVAB. Pathogenic microbiomes in BV such as G. vaginalis and P. bivia have been found to inhibit the host inflammatory response in the vaginal epithelium.121 However, only limited number of studies examine the mechanisms of how BVAB interacts with the host immune system. Previous studies revealed that G. vaginalis infection does not trigger changes in the level of pro-inflammatory mediators including IL-1β, IL-6, MIP-3α, or TNFα,122 while A. vaginae induces a broad range of pro-inflammatory cytokines, chemokines, and antimicrobial peptides including IL-1β, IL-6, IL-8, MIP-3α, TNFα, and hBD-2; whereas P. bivia induces fewer types of immune factors including IL-1β and macrophage inflammatory protein (MIP)-3α.123 However, contradictory results have been reported that P. bivia suppressed the host immune responses.124 In all, further studies are still necessary to better understand the interaction between vaginal microbiota and host immune system.
Microbiota in the development of diseases
Microbiota are complex systems consisting of trillions of microorganisms. With advanced sequencing technologies and bioinformatics, most of microbiota–related research is focusing on the relationship between microbiota compositional changes and various disease states. When subjected to external changes, the balance of microbiota community can be affected, leading to dysregulation of bodily functions and diseases125 as summarized in Fig. 4. To date, mounting evidence has confirmed that microbiota is associated with the development of CVDs, cancer, respiratory diseases, diabetes, IBD, brain disorders, chronic kidney diseases, and liver diseases. Due to the limited studies on non-bacterial species in disease development, we majorly focus on the bacterial element of the microbiota in this section. The disease-related pathogens and the signaling pathways are summarized in Table 2 and are discussed in detail in each section.

Human microbiota dysbiosis contributes to various diseases
Cardiovascular diseases
CVDs are the leading cause of morbidity and mortality worldwide, including coronary heart disease, cerebrovascular disease, peripheral arterial disease, etc. While the general risk factors include atherosclerosis, hypertension, obesity, diabetes, dyslipidemia, and mental illness, growing evidence has suggested that microbiota play a role in maintaining cardiovascular health and its dysregulation may contribute to CVDs.126 Particularly, studies on microbiota transplantation, microbiota-dependent pathways, and downstream metabolites have all shown that microbiota may influence host metabolism and CVDs through multiple metaorganism pathways. Here, we present the potential pathogenesis of microbiota in CVDs.
Oral microbiota
Periodontal diseases, which are initiated and propagated through dysbiosis of oral microbiota, have been shown to be associated with increased risk for CVDs. In 1993, DeStefano et al. reported that subjects with periodontitis had a 25% increased risk of CVDs compared with those with minimal periodontal disease, indicating a correlation between oral microbiota with CVDs.127 Multiple bacterial phylotypes were found in both the oral cavity and atherosclerotic plaques, suggesting an association between oral microbiota and atherosclerosis. Schenkein et al. presented two major mechanisms linking periodontitis with atherosclerosis.128 The first is that some microorganisms can invade endothelial and phagocytic cells within the atheroma, causing pathogenic changes and progression of the lesion, and the second includes the release of inflammatory mediators such as C-reactive protein (CRP), fibrinogen, metalloproteinases from periodontal lesions to systemic circulation. A cohort study by Lise et al. reported that the antibody levels to periodontopathogen T. forsythia were inversely related to the increased risk for CVD mortality. Indeed, other studies also linked periodontitis with several cardiovascular risk factors. A randomized controlled trial (RCT) showed that intensive periodontal treatment achieved a reduction in systemic inflammatory markers including IL-6 and CRP, and a decreased systolic blood pressure and an improvement in lipid profiles.129 IL-6 can cause cardiac hypertrophy through the IL-6 signal-transducing receptor component, glycoprotein 130. Moreover, IL-6 is able to induce the hepatic synthesis of CRP. CRP is suggested to directly influence vascular vulnerability through several mechanisms including regulating the local expression of adhesion molecules, downregulating the endothelial bioactivity of nitric oxide, altering the low-density lipoprotein ingestion of macrophages. Ramirez et al. found that, compared with the control group, periodontitis patients had higher levels of E-selectin, myeloperoxidase, and ICAM-1, which are important risk markers for CVDs.130 E-selectin is a receptor of carbohydrate ligands on the surface of leukocytes. It functions by binding with leukocytes, drawing them from the circulation toward the surface of the endothelium. Subsequently, the transmembrane glycoproteins ICAM-1 and VCAM-1, interact with integrins on the surface of the leukocyte to promote its strong binding to the endothelium, thereby contributing to CVD through their inflammatory effects on the vascular endothelium.
Gut microbiota
It is not surprising that the gut microbiota, is considered the largest endocrine organ in the body, can affect the cardiovascular system and contribute to CVDs. Gut microbiota is involved in the metabolism of choline, phosphatidylcholine, and carnitine, which eventually produce trimethylamine-N-oxide (TMAO). TMAO has been suggested to not only regulate cholesterol balance and bile acid levels but is also associated with early atherosclerosis and high long-term mortality risk of CVDs.131 Mechanistically, TMAO can activate the mitogen-activated protein kinase (MAPK) and NF-κB signaling pathways in endothelial cells and smooth muscle cells.132 The MAPK signaling pathway can be stimulated by growth factors, pathogen-associated molecules, and inflammatory cytokines, which follows by a MAPKKK-MAPKK-MAPK-TFs signaling cascade and results in the expression of inflammatory cytokines IL-6, IL-8, and TNF-α. It is well-established that NF-κB is an important mediator that regulates the activation, differentiation, and effector function of inflammatory immune cells. Therefore, the dysregulation of NF-κB may contribute to pathogenesis of atherosclerosis by promoting monocyte recruitment.133 Another inflammation mediator lipopolysaccharide (LPS), also known as endotoxin, is a component of Gram-negative bacteria that are mainly distributed in gut and oral cavity. Recent studies have shown that LPS can induce vascular oxidative stress by activating the TLR4 pathway, leading to endothelial dysfunction and vascular inflammation. A retrospective analysis conducted by Yoshida et al. suggested that patients with CVDs have higher fecal LPS levels compared with those without CVDs. It is interesting that the structures of lipid A moieties of LPS differ in bacteria, which may determine LPS activity.134
Gut microbiota is able to metabolize polysaccharides and proteins into short-chain fatty acids (SCFAs), another class of metabolites that is linked to CVDs. Most SCFAs are acetates, butyrates, or propionates. A large proportion of acetates is subjected to lipogenesis in adipose tissue and oxidize in muscle, with some being converted to butyrates by bacteria. As shown in Fig. 3, butyrates are involved in mediating the integrity of the intestinal barrier and are suggested to have direct salutary effects on intestinal epithelial cells.135 Propionates are mainly oxidized or metabolized in the liver. The potential role of these major SCFAs in CVDs has been extensive studied in animal models. SCFAs, particularly propionates and butyrates, were shown to protect the host from hypertensive cardiovascular damage.136 The propionate is suggested to regulate the balance of effector T cells and regulatory T cells, which is critically important in hypertension and hypertension-induced organ damage.137 Moreover, propionates reduced lateralization of gap junction protein connexin 43 in cardiomyocytes, thereby reducing susceptibility to ventricular tachycardia.136 The butyrate has been shown to modulate blood pressure by inhibiting expression of renal prorenin receptors and renin in animal models.138 Recently, accumulating evidence has shown that SCFAs can act on G-protein-coupled receptors Gpr41, Gpr43, and Olfr78 to mediate blood pressure. Olfr78, expressed in smooth muscle cells of vasculature, is an olfactory receptor that mediates renin secretion in response to SCFAs. Gpr41 and Gpr43 are widely expressed in the body, which will be activated upon SCFAs binding. It is established that Olfr78 and Gpr41/43 response to SCFAs through different G protein subunits and second-messenger systems. Olfr78 will activate adenylate cyclase type 3 and Golf in the olfactory signaling pathway to induce cAMP production; while Gpr41/43 activates Gαi and/or Gαo to decrease cAMP.139 Therefore, activation of Olfr78 increases hypertension by facilitating the release of renin, while activation of Gpr41 and Gpr43 counteract the hypertensive effect of Olfr78.140 These data reinforce the important role of microbiota in blood pressure control and CVD progression.
Cancer
Cancer is a disease characterized by the rapid proliferation of abnormal cells that grow uncontrollably, which can occur in almost all regions of the body. Currently, cancer is a leading cause of mortality worldwide, causing over 10 million deaths in 2020.141 Generally, the development of cancer is due to gene mutations that disrupt the cell growth or metabolic activities, and more than 100 human carcinogens are listed by the World Health Organization. Although carcinogenesis is a multifactorial process, it has been well established that tobacco, bacteria and viruses, obesity, alcohol, and radiation are the major risk factors for cancer.142 While the role of microorganisms was disregarded in cancer for a long time, the focus has shifted largely due to the finding that H. Plyori contributes to gastric cancer initiation in 1994.143 Surprisingly, recent studies have shown that microbiota plays an important role in carcinogenesis, mainly through 1) influencing the host cell proliferation and death, 2) altering immune system activity, and 3) affecting host metabolism.
Oral microbiota
Researchers have found that periodontitis, characterized by dysbiosis of oral microbiota, is involved in the initiation and progression of oral, pancreatic, genitourinary, and gastrointestinal cancers. Farrell et al. found a significant variation between the salivary microbiota of pancreatic cancer patients and healthy subjects in a retrospective case–control study.144 In cancer patients, the levels of N. elongate, and S. mitis were significantly decreased, while the level of G. adiacens was elevated compared with healthy subjects. A prospective cohort study conducted by Michaud et al. revealed that individuals with high P. gingivalis antibody levels had a twofold increased risk of pancreatic cancer compared with those with low antibody levels.145 Similarly, Fan et al. suggested that P. gingivalis was correlated with higher risk of pancreatic cancer, while Fusobacteria were associated with a decreased risk.146 Studies revealed that the abundance of T. forsythia, P. gingivalis, and F. nucleatum are significantly higher in esophageal cancer tissues compared with normal tissues.147 It is suggested that oral microbiota promote carcinogenesis by inducing chronic inflammation and producing oncometabolites.148 Since many bacteria share similar carcinogenic mechanisms, we use P. gingivalis, a pivotal periodontal bacterium, as an example to introduce the pathogenesis. Oral squamous cell carcinoma (OSCC) is the most common cancer in the head and neck region. Firstly, the presence of P. gingivalis has been shown to increase the risk of OSCC by dysregulating tissue integrity and host immune response. Cao et al. suggested that P. gingivalis can bind to TLR4 receptor, which in turn activate NF-κB pathway and overstimulate the downstream JAK1/STAT3 signaling pathway, leading to inhibition of cell apoptosis.149 TLRs are characterized as primary sensors that response to microbial components and trigger immune response. All TLR signaling pathways eventually activate NF-κB pathway, which controls the expression of a wide range of inflammatory cytokines. Secondly, studies have shown that P. gingivalis can stimulate the proliferation of epithelial cells by interfering with the cell cycle regulation. Kuboniwa et al. reported that P. gingivalis can affect signaling pathways involving cyclins, p53, and PI3K.150 Cyclins are subunits of CDK complexes that regulate the progression of cell cycle and thus proliferation. p53 is a tumor suppressor gene that has been well-established in the cause of cancer. Activation of p53 can cause cell cycle arrest and apoptosis, thus mutation or loss-of-function of p53 may lead to uncontrolled cell growth.151 Moreover, P. gingivalis has been shown to interact with β-catenin, a key protein in regulating cell proliferation and tumorigenesis. The Wnt/β-catenin signaling is a versatile pathway that involved in many human diseases. Aberrant activation of Wnt/β-catenin pathway results in the accumulation of β-catenin in the cells and thus upregulating the expression of oncogenes including CyclinD-1 and c-Myc. Zhou et al. suggested that P. gingivalis can induce noncanonical activation of β-catenin and dissociation of the β-catenin destruction complex via gingipain-dependent proteolytic processing.152 Thirdly, P. gingivalis may induce chronic inflammation by increasing levels of cytokines including IL-8, TGF-β1, and TNF-α. IL-8 and TGF-β1 can enhance the invasiveness of tumor cells by upregulating matrix metalloproteinases.153 TNF-α can lead to gene mutations through the generation of reactive oxygen species (ROS) or reactive nitrogen (RNS) intermediates as well as induce epithelial–mesenchymal transition, which stimulates tumor angiogenesis.154 Lastly, P. gingivalis can produce oncometabolites such as acetaldehydes and oxygen radicals. Accumulation of these metabolites are known to promote chronic inflammation and cause DNA damage and mutagenesis, leading to cancer development. Recent studies also suggested that intestinal colonization of oral microbiota contributes to several health issues including carcinogenesis.155 F. nucleatum, a periodontal pathogen, has been extensively studied in colorectal cancer (CRC). By comparing cancer and adjacent normal tissues, it was found that F. nucleatum was significantly enriched in tumor tissues and may promote CRC progression by increasing tumor multiplicity and selectively recruiting tumor-infiltrating myeloid cells.156
Respiratory microbiota
The focus of respiratory microbiota and cancer is largely on lung cancer. The lung, which was once considered sterile, is colonized by different microbiota throughout the respiratory tract. In healthy individuals, the core microorganisms in the lung are Pseudomonas, Streptococcus, Prevotella, Fusobacterium, Haemophilus, Veillonella, and Porphyromonas. In a systematic review, Perrone et al. summarized that levels of Actinomyces, Veillonella, Streptococcus, Megasphaera, and Mycobacterium were more abundant in lung cancer patients compared with healthy individuals. In addition, Gomes et al. reported that a squamous cell carcinoma subcluster with the worst survival was correlated with several Enterobacteriaceae.157 Another study suggests that in the microbiota of patients with lung cancer, unlike in the control group, has high levels of Streptococcus, indicating it may be a possible diagnostic marker. Interestingly, Peter et al. found no relationship between tumor tissue microbiota with lung cancer recurrence, while the higher richness and diversity in adjacent normal tissue was associated with worse outcome.158 Although the underlying carcinogenesis mechanisms are not fully elucidated, dysbiosis of lung microbiota increases inflammation and host immune modulation, which are two important pathways related to cancer. Jin et al. reported that microbiota induced inflammation associated with lung adenocarcinoma via activation of lung-resident γδ T cells, facilitating the proliferation of tumor cells.159 Interestingly, previous studies suggested that γδ T cells are able to recognize cancer cells and initiate anticancer activity, largely related to cytotoxicity and interferon-γ production. Therefore, it remains inconclusive how the immune system response to lung microbial. In an epithelial cell model, exposure to Streptococcus, Prevotella, and Veillonella led to the upregulation of PI3K and ERK1/2 signaling pathways, which mediates cell proliferation, differentiation, and survival.160 PI3K/Akt/mTOR is one of the most important cell signaling pathways and a well-established mediator of cancer. Activation of PI3K/Akt/mTOR pathway, by gene mutation, inactivation of PTEN, or activation of upstream oncogenes, contributes to the development of tumor and resistance to therapeutics. The same research group used an in vivo non-small cell lung cancer mouse model to show that microbiota dysbiosis led to upregulation of PI3K/AKT, ERK/MAPK, IL17A, IL6/8, and inflammasome pathways, suggesting that microbiota can contribute to the pathogenesis of lung cancer.161
Gut microbiota
Increasing evidence suggests that gut microbiota is associated with the initiation and progression of CRC. Studies have shown that dysbiosis of gut microbiota can trigger inflammation and immune response that are indirectly related to carcinogenesis.162 Grivennikov et al. reported that microbial products may induce epithelial barrier deterioration, which trigger tumor-elicited inflammation and drive initiation and progression of CRC.163 It is suggested that gut microbiota can promote CRC progression by affecting certain signaling pathways including E-cadherin/β-catenin, TLR4/MYD88/NF-κB, and SMO/RAS/p38 MAPK.164 Chen et al. suggested that both commensal and pathogenic bacteria facilitate CRC progression via 1) exploiting tumor surface barrier defects, 2) invading normal colonic tissue and inducing local inflammation, and 3) producing genotoxic metabolites to induce oncogenic transformation of colonic epithelial cells.165 It has been characterized that the major bacteria that contribute to CRC are E. faecalis, E. coli, B. fragilis, S. bovis, F. nucleatum, and H. pylori.166 These bacteria are able to produce genotoxic substances such as colibactin, B. fragilis toxin, and typhoid toxin that cause host DNA damage. For example, intestinal F. nucleatum was evaluated in several CRC studies.167 The abundance of F. nucleatum was significantly higher in mucosal and fecal samples of CRC patients compared with healthy controls. These studies also indicated that F. nucleatum can invade CRC tumor cells, leading to the presumption that F. nucleatum may influence tumorigenesis.
Diabetes mellitus
Diabetes mellitus (DM) refers to a group of diseases that affect glucose regulation. DM can be classified as type 1 diabetes mellitus (T1DM), Type 2 diabetes mellitus (T2DM), and gestational diabetes mellitus (GDM). T1DM is caused by the autoimmune response against pancreatic β cells, while T2DM is characterized as the inability of the body to produce or utilize insulin properly. GDM is one of the most prevalent pregnancy complications and is associated with increased risk of maternal and fetal metabolic disorders. The relationship between microbiota and DM has been extensively studied and the correlation between microbiota dysbiosis and onset of DM is well established.
Type 1 diabetes mellitus
In T1DM, the microbiota is an attractive research field due to its close relationship with chronic inflammation and immune response. The composition of oral and fecal microbiota appears to be distinct in T1DM patients in multiple studies. Groot et al. found that Christensenella and Bifidobacteria were enriched in fecal samples.168 Oral Streptococcus was positively associated with T1DM, while fecal Streptococcus was inversely correlated with T1DM. In addition, T1DM patients may exhibit decreased levels of SCFA butyrate-producing bacteria, which are key factors in decreasing chronic inflammation and maintaining intestinal homeostasis.169 This finding was reinforced by other case-control studies which showed that the levels of R. faecis, F. prausnitzzi, Intestinimonas were significantly lower in T1DM patients than in healthy controls.170 However, inconsistent results were reported with other bacteria, suggesting that further research is required.171 Interestingly, Vatanen et al. analyzed data from TEDDY study and showed that healthy children contained more genes related to fermentation and the synthesis of SCFAs without significant association to specific taxa.172 Therefore, it is possible that the T1DM-related microbial factors are taxonomically diffuse but functionally coherent. The research on microbiota in T1DM was conducted mainly in animal models, therefore, the pathogenic mechanisms require further validation in human. It was first introduced that the development of T1DM may be dependent on microbiota in 1987 by Suzuki et al.173 It was suggested that microbiota can contribute to T1DM mainly through immune response modulation. T1DM is defined as an autoimmune disease by chronic inflammation of the pancreatic islets of Langerhans. Since microbiota is involved in the initiation of chronic inflammation, it is not surprising that its dysregulation may contribute to T1DM. Higuchi et al. reported that the plasma levels of IL-6 were significantly higher in T1DM patients than in healthy controls, which was correlated with the abundance of Ruminococcaceae and Ruminococcus. Leiva-Gea at al also reported that T1DM patients had increased levels of proinflammatory cytokines IL-1β, IL-6, and TNF-α and decreased levels of anti-inflammatory cytokines IL-10 and IL-13, which were significantly correlated with the abundance of different bacteria.170 In addition, an upregulated level of LPS was observed, which is known to induce the release of proinflammatory cytokines and impair pancreatic β cells.174 Clinical data showed that T1DM patients have significantly elevated levels of TLR2 and TLR4 ligands, indicating increased TLR2 and TLR4 activity.175 As described above, TLRs play an important role in innate and adaptive immunity, which protect the body from infectious microorganisms. The role of TLR4 was further evaluated in mouse models. Elke et al. reported that TLR4 accelerates the development of diabetes, suggesting that TLR4 is involved in the progression of insulitis.176 The TLR4/MyD88 pathway regulates the activation of NF-κB and the levels of pro-inflammatory cytokines such as IL-6 and TNF-α.177 Wen et al. established an MyD88-negative Non-Obese Diabetic mice and found that the mice lacking MyD88 protein do not develop T1DM. Taken together, it suggests that microbiota may facilitate the progression of T1DM via TLR4/MyD88 signaling pathway.
Type 2 diabetes mellitus
In terms of T2DM, gut microbiota has been linked to disease development. Numerous studies have confirmed that the composition of gut microbiota is altered in T2DM patients.178 Larsen et al. reported that the abundance of Firmicutes and Clostridia were significantly decreased in T2DM patients compared with the control group. In addition, the ratios of Bacteroidetes to Firmicutes, Bacteroides-Prevotella group to C. coccoides-E. rectale group were positively correlated with blood glucose level.179 Almugadam et al. showed that the abundance of SCFA-producing bacteria Facalibacterium and Roseburia were significantly decreased in T2DM patients.180 Antidiabetic agents were able to improve the diversity and richness of gut microbiota and enriched gut ecosystem with beneficial bacteria. The underlying molecular mechanisms of gut microbiota contributing to T2DM may include modulation of inflammation, gut permeability, and glucose metabolism. Generally, T2DM is associated with increased levels of pro-inflammatory molecules. LPS is well documented to promote low-grade inflammation. Several studies have suggested that T2DM patients have increased level of LPS in peripheral circulation.181 LPS can bind to TLR4, triggering macrophage aggregation and activating the NF-κB signaling pathway. This interaction leads to the release of inflammatory factors, resulting in the inhibition of insulin secretion. Gut microbiota can metabolize primary bile acids into secondary bile acids. Secondary bile acids bind to the farnesoid X receptor and release fibroblast growth factor, FGF19/15, which is able to promote insulin sensitivity and glucose tolerance.182 Therefore, dysbiosis of gut microbiota may lead to abnormal bile acid metabolism by affecting glucose metabolism. Another class of important metabolites from gut microbiota are SCFAs. Studies suggest that SCFAs play an important role in mediating glucose metabolism and insulin sensitivity via multiple signaling pathways. For instance, SCFAs can bind to Free Fatty Acid Receptor FFAR2 or FFAR3 on intestinal L cells, stimulating the release of glucagon-like peptide-1 (GLP-1) and peptide YY, which are known to promote insulin secretion and reduce glucagon.183 In addition, butyrates can protect the integrity of the intestinal barrier, which may be damaged in T2DM patients due to low-grade inflammation. Moreover, SCFAs are important anti-inflammatory mediators that can limit autoimmune response by promoting the production of regulatory T cells.184 Therefore, the reduced abundance of SCFA-producing bacteria may contribute to the development of T2DM.
Oral microbiota may also play a role in T2DM. Oral bacteria can translocate to the gut, changing the composition of gut microbiota and potentially mediating immune response.185 Several studies have identified significant alterations in oral microbiota composition between T2DM patients and healthy controls. Interestingly, Xiao et al. reported that T2DM induces a shift in oral microbiota composition with enhanced IL-17 level.186 By transferring to GF mice, the DM-modified oral microbiota is more pathogenic, indicating that DM can increase the risk and severity of periodontal disease.
Gestational diabetes mellitus
In GDM, several studies reported that gut microbiota mediates insulin resistance and inflammation during pregnancy. Metabolic disorders are commonly seen in GDM women, including enhanced insulin resistance and downregulated insulin secretion.187 During pregnancy, the composition of gut microbiota undergoes substantial changes, which may account for the development of GDM. For example, positive correlation has been identified between insulin and Collinsella, gastrointestinal polypeptide and Coprococcus, and adipokine with Ruminococcaceae and Lachnospiraceae.188 Moreover, Koren et al. demonstrated that gut microbiota changed from first to third trimesters, with increased diversity and decreased richness.189 It was shown that GDM patients had increased Firmicutes to Bacteroidetes ratio, an important factor that facilitates obesity and aggravates inflammation.190 The abundance of SCFA-producing bacteria was significantly lower in GDM pregnancies compared with healthy controls, indicating that the elevated blood glucose levels may be caused by microbiota alteration.191 Studies also revealed that the gut microbiota composition in the offspring of GDM mothers were different from those in non-GDM mothers. Ponzo et al. reported that the abundance of proinflammatory bacteria was higher in GDM infants than in healthy controls.192 Other studies confirmed this finding that the GDM infants had lower α-diversity compared with the control group, and the abundance of certain lactic acid bacteria may be affected by maternal GDM status.193 Therefore, gut microbiota may play a critical role in the development of GDM and may also affect GDM infants.
Respiratory diseases
Respiratory diseases are a group of diseases that affect the lungs and other parts of the respiratory system and include chronic diseases (asthma and chronic obstructive pulmonary disease (COPD), pulmonary fibrosis) and pneumonia. Extensive studies have suggested that oral, lung, and gut microbiota are associated with the development of respiratory diseases. In this section, we will discuss the major findings demonstrating a connection between the microbiota and the development of respiratory diseases.
Chronic respiratory diseases
COPD and asthma are the two most frequently diagnosed chronic respiratory diseases. COPD is defined as a disease state characterized by the presence of airflow limitation associated with chronic bronchitis or emphysema. Asthma is a heterogeneous syndrome of chronic airway inflammation characterized by bronchial hyper-responsiveness to environmental triggers and by symptoms including wheezing, shortness of breath, and chest tightness. Accumulating data suggest that lung microbiota is actively involved in the development of chronic respiratory diseases. The composition of lung microbiota was found to be distinct between patients and healthy individuals. Using 16s rDNA sequencing technology, studies have identified that asthma patients had higher bacterial load and diversity, increased abundance of Proteobacteria, and decreased abundance of Bacteroidetes and Firmicutes.110 In addition, Woerden et al. found a different pattern of fungi, particularly Malassezia pachydermatis, in the sputum samples of asthma patients and controls. However, the research on fungi is limited and remains inconclusive.194 Other studies identified altered abundance of Pseudomonas, Moraxella, Lactobacillus, and Haemophilus during COPD exacerbations.195 Recent studies also found a potential relationship between pulmonary fibrosis and viral and bacterial infection. A clinical trial reported that the progression of pulmonary fibrosis is associated with specific Staphylococcus and Streptococcus bacterial species.196 Chronic inflammation induced by lung microbiota may be the key process in the initiation of chronic respiratory diseases. In asthma patients, Proteobacteria has been associated with hyper-responsiveness and Th17/IL-17-mediated inflammation.197 H. influenza, the most isolated pathogen from asthma patients, can induce steroid-resistant neutrophilic allergic airway diseases.198 Alnahas et al. demonstrated that Proteobacterium M catarrhalis can exaggerate allergic airway diseases by triggering a strong immune response characterized by neutrophilic infiltration, high levels of IL-6 and TNF-α, and moderate levels of Interferon (IFN)-γ and IL-17 in a mouse model.199 Furthermore, Garcia-Nuñez et al. reported that the bronchial microbe Proteobacteria may induce chronic inflammation and predict high disease severity.200 These studies highlighted that lung microbiota dysbiosis may potentially be associated with the development of chronic respiratory diseases.
Gut microbiota is a potent modulator of pro-inflammatory and autoimmune responses, leading to different inflammation-related diseases. Multiple studies have linked the dysbiosis of gut microbiota early in life to increased risk of asthma later in life, known as the gut-lung axis. The gut-lung axis in chronic respiratory diseases has been extensively studied and reviewed.112 It is suggested that gut microbiota dysbiosis in early life may lead to the development of respiratory diseases, since gut microbiota plays an important role in immune cell maturation and pathogen resistance.201 Indeed, Roussos et al. and Rutten et al. demonstrated that patients with chronic GI diseases have higher prevalence of chronic respiratory diseases including asthma and COPD, while the mechanisms are still unclear.202,203 Sprooten et al. reported that patients with acute COPD exacerbations had increased GI permeability, suggesting that gut microbiota is involved in exacerbations.204 Another study demonstrated that the increased levels of gut microbiota-dependent circulating TMAO were associated with all-cause mortality in COPD patients.205 Arrieta et al. discovered that the abundance of Veillonella, Faecalibacterium, and Lachnospira were significantly decreased in children at risk of asthma.206 It is suggested that the gut bacterial metabolites may contribute to asthma through its immune modulation. For example, Roduit et al. reported that children with high level of SCFAs are less likely to have asthma at later stage.207 SCFAs have been shown to promote peripheral regulatory T-cell generation208 and ameliorate inflammation in allergic asthma models.209 In addition to bacterial metabolites, it is suggested that lymphocytes with altered homing properties may contribute to asthma.210 Under normal situation, lymphocytes are thought to exhibit tissue specificity to the site where they first encounter the antigen. However, intestinal lymphocytes from IBD patients are known to lack tissue specificity and may account for the presence of inflammation in organs other than the gut. Huang et al. reported that innate lymphocytes were recruited from the gut to the lungs following inflammatory signals from IL-25.211 Interestingly, some data indicates that the gut-lung axis may have a bidirectional interaction. Perrone et al. showed that pneumonia induced intestinal epithelial apoptosis212 and decreased intestinal epithelial proliferation213 in mice.
Oral microbiota has been associated with chronic respiratory diseases due to the contiguous anatomic structure and microaspiration.214 Early studies found significant similarity between the oral and lung microbiota, while the nasal microbiota shares less similarities with lung microbiota.215 It is hypothesized that the oral microbiota may contribute to chronic respiratory diseases through aspiration and systemic inflammation. It is possible that aspiration of oral bacteria into the lung leads to lung microbiota dysbiosis and inflammation. Segal et al. reported that the enrichment of oral bacteria Veillonella and Prevotella in bronchoalveolar lavage samples has been associated with subclinical inflammation, characterized by increased neutrophils and lymphocytes.216 A RCT showed that bronchial microbiome of asthmatic subjects was uniquely enriched with two periodontal pathogens, Fusobacterium and Porphyromonas.217 Many periodontitis-related inflammatory cytokines have also been detected in chronic respiratory diseases. Aaron et al. reported that TNF-α was increased in the sputum of COPD patients.218 Substantial studies have shown that TNF-α can stimulate the generation of ROS in pulmonary tissues, accompanied by the generation of various adhesive and proinflammatory molecules such as VCAM-1, ICAM-1 and RAGE. TNF-α is also suggested to function as a pro-inflammatory cytokine in asthma that recruits neutrophils and eosinophils.219 Periodontitis is related to high levels of systemic inflammatory markers, such as CRP and IL-6. Jousilahti et al. reported that the level of CRP was significantly associated with asthma prevalence.220 However, the oral-lung axis has not been fully understood and deserves further investigation.
Pneumonia
The normal respiratory tract and gut microbiota protect against pneumonia by preventing pathogenic bacteria colonization and by modulating immune responses. Therefore, it is not surprising that the dysbiosis of respiratory tract microbiota is considered a risk factor of pneumonia.
The upper airways are the main source of microbes to the lower airways. Recently, researchers have shown that the reduction of nasal microbiota diversity increased susceptibility to pneumonia. Particularly, three microbiota profiles dominated by Lactobacilli, Rothia, and Streptococcus were significantly associated with pneumonia.221 In neonates, the pathogenic bacterial colonization of the airways with S. pneumonia, H. influenza, and M. catarrhalis were associated with increased risk of pneumonia and bronchiolitis.222 Regarding the lower airway microbiota, studies suggested that increased abundance of Prevotella and Veillonella predisposed pneumonia in HIV patients.223 In addition, altered immune response due to microbiota dysbiosis may increase the risk of pneumonia. For example, dysregulation of SCFA-producing bacteria may contribute to the development of pneumonia. Segal et al. suggested that pulmonary SCFAs correlated with increased anaerobic bacteria.224 Indeed, SCFAs have a direct inhibitory effect on immune response via suppression of IFN-γ and IL-17A pathways. During bacterial infection, neutrophils are rapidly migrated to lung parenchyma and alveolar. The IFN-γ released by neutrophils regulates bacterial clearance, therefore the level of IFN-γ is critical for host defense during pneumonia.225 Similarly, Th17 cells and its signature IL-17A signaling is an important immune response against pneumonia. During infection, IL-17A acts on nonimmune cells to trigger the release of antimicrobial proteins, cytokines, and chemokines, thus enhance innate immunity during microbial infection.226 By inhibiting the IFN-γ and IL-17A pathways, it allows the lung bacteria reproduction and worsen inflammation. Salk et al. reported that the influenza-specific lgA production is significantly associated with levels of Lactobacillus, Prevotella, Veillonella, Bacteroide, and Streptococcus.227 Interestingly, recent studies suggested that commensal microbes can play a crucial role in the development of pneumonia. Recently, the global pandemic COVID-19 has become a major research area in respiratory disease. Emerging data are now connecting the COVID-19 mortality with microbiota dysbiosis. Fan et al. investigated the lung microbiota characteristics from 20 deceased COVID-19 patients.228 It is suggested that the dysbiosis of lung microbiota is characterized by increased abundance of Acinetobacter spp., which are related to multidrug resistance and mortality. In addition, Cryptococcus was the dominant fungi in the lung fungal communities, along with Issatchenkia, Cladosporium, Candida, etc. Han et al. reported that COVID-19 may induce severe dysbiosis of lung microbiota, particularly with increased abundance of Klebsiella oxytoca, Faecalibacterium prausnitzii, and Rothia mucilaginosa.229 Segal et al. revealed that the enrichment of lower airways with oral bacteria Mycoplasma salivarium was associated with poor clinical outcome.230 However, no significant connection was found between increased mortality and secondary respiratory pathogens.
Gut microbiota is another major subject when studying pneumonia-microbiota interaction. Schuijt et al. identified that the gut microbiota plays a protective role against S. pneumoniae infection.231 Compared with the control group, S. pneumoniae infection in gut microbiota depleted C57BL/6 mice demonstrated increased bacterial dissemination, inflammation, organ damage and mortality. In addition, depletion of gut microbiota was associated with the upregulation of metabolic pathways, leading to reduced responsiveness to inflammatory cytokines. In accordance, Felix et al. showed that the commensal gut segmented filamentous bacteria protected immunodeficiency mice from S. pneumoniae infection. It is likely that the bacteria promoted a shift in lung neutrophil phenotype from inflammatory to pro-resolution, which is similar to heat-inactivated S. pneumoniae treatment. Recent data also suggested that gut microbiota composition may reflect disease severity in COVID-19 patients.232 In COVID-19 patients, decreased abundance of several gut commensals was observed, including Bifidobacteria, Eubacterium rectale, and Faecalibacterium prausnitzii. The dysbiosis gut microbiota was positively associated with disease severity, with elevated levels of inflammatory cytokines and blood markers such as CRP, aspartate aminotransferase, and lactate dehydrogenase. Therefore, unlike in chronic respiratory diseases, the gut-lung axis may provide additional protection for the host against pneumonia by regulating the immune response.
Inflammatory bowel disease
IBD is a chronic and remittent inflammatory condition of the GI tract, encompassing several diagnoses including Crohn’s disease (CD) and ulcerative colitis (UC).233 While UC is known as continuous, diffuse, and superficial inflammation of the colon, CD is characterized by discontinuous, transmural lesions affecting different regions of the GI tract.234
Although the development of IBD is due to complex multifactorial mechanisms, several risk factors have been extensively studied and are now well documented. The pathogenesis of IBD involves dysregulated immune response, genetic mutations, and environmental factors.235 The intestinal barrier plays an important role in maintaining homeostasis; dysfunction of the barrier may lead to ulceration. Specifically, the intestinal barrier would be susceptible to pathogen invasion without the secretion of antimicrobial peptides (AMPs) or tight junction proteins.236 During initial disease in genetically susceptible individuals, the immune response is altered, leading to loss of immune tolerance to intestinal antigens. This subsequently stimulates the differentiation of helper T cells and release of chemokines and proinflammatory cytokines, which induce chronic inflammation of the intestine.237 In addition to immune dysregulation, genetic factors are involved in determining IBD development. For example, genetic mutations associated with CD include polymorphisms for the Nucleotide Oligomerization Domain Containing 2 (NOD2/CARD15), Immunity-elated GTPase family M (IRGM), and autophagy-related 16 Like 1 (ATG16L1).238
Studies have shown that gut microbiota are highly associated with the development of IBD. Mechanistically, microbiota dysbiosis is linked to IBD through its impact on inflammation as well as the intestinal barrier. As described before, microbiota dysbiosis can induce chronic inflammation, which is associated with the development of multiple diseases such as cancer, diabetes, and heart diseases. Importantly, it is postulated that microbiota can interact with intestinal barrier and lead to IBD. For example, Kleessen et al. reported that bacterial invasion of the mucosa was detected more in IBD patients than in controls.239 It was also reported that the abundance of adherent-invasive E. coli was significantly increased in CD patients, suggesting that the pathogenic bacteria may affect the permeability of the intestine, the composition of gut microbiota, and eventually induce intestinal inflammation.240 In healthy individuals, the predominant phyla are Firmicutes and Bacteroidetes, followed by Proteobacteria and Actinobacteria. Multiple studies have revealed that the composition of gut microbiota is different between IBD patients and healthy controls.241 For example, the ratio of Bacteriodetes to Firmicutes is decreased while the abundance of gammaproteobacterial increased in IBD patients.242 The protective and normal bacteria, Bacteroides, Eubacterium, and Lactobacillus are significantly reduced in CD and UC patients.243 A meta-analysis study suggested that enterohepatic Helicobacter species, but not intestinal H. pylori infection, was significantly related to IBD.244 It should be noted that, although many studies provided the association between microbiota dysbiosis and IBD, the causation remains to be determined.245 It is possible that the microbiota dysbiosis can be considered a response to the environmental changes due to intestinal inflammation. The possible role of fungi and viruses in IBD are also being studied and reviewed, but no link has been established thus far.234,245
While most of the studies are focusing on gut microbiota, oral microbiota is gaining attention with the characterization of the oral-gut axis. Kitamoto et al. showed that pathobionts and pathogenic T cells of oral origin were able to translocate and colonize in intestines, causing IBD in periodontitis mouse models.246 Derrien et al. concluded that the bacteria residing in the oral cavity and GI tract maintain intimate relationships,247 supporting the notion of an oral-gut axis. Recently, a meta-analysis by She et al. demonstrated that periodontitis was significantly associated with IBD, while the mechanisms are undetermined.248 Another study by Kimura et al. suggested that, in the salivary microbiota of IBD patients, the abundance of Bacteroidetes was significantly increased with a concurrent decrease of Proteobacteria.249 They also found a significant correlation between inflammatory cytokine levels and the abundance of Streptococcus, Prevotella, Veillonella, and Haemophilus, implicating a possible relationship between dysbiosis of oral microbiota with inflammatory response in IBD patients. A case-control study by Vavricka et al. showed that both periodontitis and gingivitis marker levels were increased in CD patients compared with healthy controls.250 Although these studies have established an association between oral diseases with IBD, data regarding oral microbiota in IBD is still limited and require further investigation.
Brain disorders
Neuropsychiatric and neurodegenerative disorders of the brain, along with many other comorbidities, were known to be responsible in causing significant mortality in different population subsets. Extensive research over the years have shown to implicate the role of microbial diversity in brain disorders by modulating the factors linked with the development of these disorders. One example is data from a meta-analysis study which shows that depression is responsible for an increase in relative risk of mortality from all causes, specifically about 1.86 times more than non-depressed patients.251 Microbiota-induced hyperactivity of the HPA axis and inflammation are also shown to be associated with provoking depression.252
Neuropsychiatric disorders
Gut microbiota is believed to play a vital role in mediating neuronal behavior via gut-brain axis.253 Preclinical studies have established that gut microbiota affects cognitive performance, repetitive behaviors, and social interactions in different animal models.62,254,255 One of the plausible hypotheses about gut microbiota’s involvement in affecting neuronal disorders is described by stress-induced intestinal permeability, permitting endotoxins to enter the blood circulation, thereby triggering an immune response.256 This peripheral inflammation can also influence mental health by promoting the entry of neurotoxins into the brain and also by obstructing neurotransmitter systems.257 Although the direct mechanism of gut bacteria influencing neuropsychiatric disorders was clearly not studied, many studies believe that gut-induced stress has a vital role along with disrupted gut microbiome and various other factors in causing depression, anxiety, and other psychological disorders. Recently, Jiang et al. has demonstrated that fecal samples from patients with major depressive disorder have shown increased Bacteroidetes, Protobacteria and Actinobacteria along with less Firmicutes when compared with fecal samples from healthy controls.258 Decreased expression of certain families such as Lachnospiraceae and Ruminococcaceae within the phylum Firmicutes was reported, and this is believed to be correlated with behavioral changes caused by stress. Some bacterial genera such as Roseburia, Blautia, Lachnospiraceae, and Ruminococcaceae are associated with synthesis of SCFA (responsible for barrier integrity) and anti-inflammatory properties. It was believed that there was an extensive correlation between diversity of gut microbiota and mood-related behaviors, especially depressive disorder.259
Both major brain disorders, depression and anxiety are indicated to be influenced by stress-regulated HPA axis pathway, which is believed to be strongly modulated by gut microbiota composition.257,260 An epigenetic study using GF mouse models had demonstrated that GF mice showed significant difference in gene expression in the brain systems when compared with control mice, notably in the areas of cortex, cerebellum, striatum, and hippocampus.261 It is suggested that the gut-brain axis may be affected by regulation of stress hormones and the establishment of neuronal circuits. Based on this initial finding, many studies have investigated the substantial changes observed in GF mice compared with the wild-type controls. In GF rats, increased levels of neurotransmitters such as norepinephrine, dopamine, and serotonin were reported in the striatum, whereas dopaminergic turnover was found to be decreased in the frontal cortex, striatum, and hippocampus of GF rats.262 Few other study findings using GF mice models have demonstrated a reduction in brain-derived neurotropic factor and nerve growth factor-inducible protein A in several brain regions, and an increase in synaptophysin and post synaptic density (PSD-95) proteins in GF mice brain systems when compared with controls.261 When it comes to autism spectrum disorder (ASD), several pre-clinical studies have reported that gut dysbiosis induced significant neurodevelopmental changes in mouse models of ASD.263 Species like Clostridium and Ruminococcus was found to be different when compared between autism children and controls.264 Adams et al. demonstrated that symptoms of GI discomfort were correlated with severity of autism in children.265 A small pilot scale study conducted by Kang et al. using fecal transplantation of standardized gut microbiota to children diagnosed with autism spectrum disorder has enhanced GI function and decreased behavioral ASD scoring.266
Neurodegenerative disorders
Lately, accumulating body of evidence from various studies are emphasizing the importance of gut microbiota in the progression of various neurological disorders such as Alzheimer’s disease (AD), cerebrovascular stroke (CVS), Parkinson’s disease (PD), and schizophrenia etc. It has been evident that gut microbes are involved with regulation of brain function via its effect on host innate immunity.267 The community composition is also found to be varied depending on the way of newborn delivery. Vaginally delivered newborns are colonized with microbiota from the maternal genital tract and is more heterogenous compared with newborns delivered by Cesarean section.268 Cesarean delivered newborns displayed less brain electrical activity, which is supported by in vivo studies using Cesarean-delivered rats, where these rats exhibited pre-pubertal alterations in the development of cortex and hippocampus.269
During the presymptomatic stages of PD, α-synuclein-mediated Lewy body pathology was observed in the ENS and dorsal motor nucleus of the vagus nerve. A total of 38 human fecal samples were analyzed using 16s rRNA sequencing, displaying significant differences in bacterial composition such as decreased Blautia, Faecalibacterium and Ruminococcus and increased Escherichia-Shigella, Streptococcus, Proteus, and Enterococcus.270 Li et al. in a study conducted in China, has identified significant diversity in different taxa such as increases in Prevotella, Akkermansia and decreased abundance in Lactobacillus species in PD patients when compared with healthy controls.271 These species play a prominent role in affecting the harmony of gut homeostasis, for example, increased numbers of Akkermansia were shown to be responsible for increased intestinal permeability and facilitating pathogen entry.272 Increased level of certain Prevotella species is associated with mucin synthesis in the gut mucosal layer and production of SCFAs, which are shown to mediate neuroinflammation in mouse models of PD.67 However, another study has reported a decreased abundance of Prevotella in PD patients compared with healthy controls, which led to the need of additional studies or bigger sample size to understand the specific role of Prevotella and its family in progression of PD.273 Differences in genotype, diet, and lifestyles of the population subsets might be a potential reason for this disparity in reports. Pathological features of AD were characterized by the presence of amyloid-β plaques and intracellular tau based neurofibrillary tangles (NFT). Vogt et al. have reported significantly less microbiome diversity in AD patients compared with healthy controls. Particularly in this study, a decrease in phylum like Firmicutes, Actinobacteria (member of Bifidobacterium) and an increase in phylum of Bacteroidetes and Proteobacteria were observed in the AD group.274 Reduction in Firmicutes and Bifidobacterium has been well studied for their association with T2DM and inflammation, which are identified as major risk factors for AD.179 Inflammatory intestinal bacterial taxa are found to be associated with high level of inflammatory cytokines, including IL-6, TNF-α, CXCL2, NLRP3, and brain amyloidosis in a study conducted in older people suffering from cognitive disorders.275 In the same study, increased levels of pro-inflammatory cytokines were observed with increased numbers of Escherichia/Shigella. Colonization of certain pathogenic bacterial strains such as Toxoplasma and Chlamydiaceae pneumoniae has also been suggested for their roles in chronic neuroinflammation and NFT in AD.276
CVS is a major neurological condition associated with neurological defects and impairment in cognitive functions leading to disability and mortality. Acute middle cerebral artery occlusion-induced stroke mouse models have shown reduced species diversity and increased growth of Bacteroidetes in mice, while fecal transplantation of normal gut microbiota normalized brain lesion-induced dysbiosis and improved stroke outcomes.277 Another preclinical study using mice has reported a significant change in cecal microbiota, such as Prevotellaceae and Peptococcaceae, the former of which is a core part of microbiota in mice, although their functionality in humans is yet to be identified.278 Additionally, experiencing stress before stroke might increase the bacterial translocation from the intestine to the blood stream, triggering immune responses.279 Despite the number of studies done in strengthening the idea of gut microbiota involvement in orchestrating neuronal harmony, additional studies are required to identify the clinical benefits of targeting specific microbiota in treating these conditions.
Oral and respiratory microbiota in brain disorders
Oral microbiome is another key contributor to the development of neurological disorders, as improper maintenance of oral health can influence the growth of complex communities on the surface of teeth, tongue, or under the gum.280 Hicks et al. has reported that salivary microbiome analysis has shown wide differences between ASD, typically developing, and non-developmentally delayed groups of children.281 In a depression-related study, 1S rRNA gene-based next-generation sequencing was used to profile the bacterial composition of saliva in depressed patients compared with young adults; it has shown diversification but importantly, increased Prevotella nigrescens and Neisseria was observed in depressed individuals.282 Smoking and alcohol consumption are two major factors that induce dysbiosis in oral microbiota and promote growth of pathogenic bacteria.280 A meta-analysis study has revealed that drinking alcohol is associated with pathogenesis of AD and also with significantly decreased level in Firmicutes phyla and an increased level in Bacteroides phyla in these patients.283 In a cross-sectional case control design study on PD, around 16 bacterial families were found to be altered in early-stage PD patients.284 Among them, variation in families like Bifidobacteriaceae, Saccharomycetaceae and Lactobacillaceae were studied extensively for their role in progression of PD. A cohort study with 68 patients comprising of AD and control groups have reported differential abundance of two specific taxa Pasteurellaceae and Lautropia mirabilia, which were found to be associated with mild cognitive impairment.285 Additionally, another study conducted in 78 patients have revealed increased relative abundance of Moraxella, Leptotrichia and Sphaerochaeta and decreased Rothia in saliva of AD patients when compared with healthy controls.286 Unfortunately, not many studies have been reported about respiratory microbiota for its role in neurological disorders, and the research concerning respiratory microbiota is still at infancy stage. Although the affinity of microbial dysbiosis in many neurological disorders is being extensively studied, currently there is no gold standard to interlink the changes in microbial environment with the pathogenesis of these disorders. More preclinical and clinical studies targeting microbiome are required to understand the extent and complex nature of microbiome’s association with the development of several brain disorders.
Chronic kidney diseases
Around 9% of the global population suffer from chronic kidney disease (CDK).287 Co-morbidities like diabetes, hypertension and heart disease are considered some of the major risk factors for CKD.288 CKD is physiologically identified as a decrease in glomerular filtration rate (GFR) < 60 ml/min per 1.73 m2 or by the existence of albuminuria for 3 or more months. Gradual loss of kidney function and irreversible renal structural changes are the main characteristics observed in CKD patients.
Gut-kidney axis communication and gut microbiota
Differences in microbial ecosystems were studied persistently for their involvement in the progression of CKD.289 Recently, oral microbiota were extensively studied for the role in mediating chronic systemic inflammatory dysregulation. It has been reported that conditions affecting oral microbiota like periodontitis indirectly affects CKD by augmenting systemic inflammation.290 Biomarker-based human studies have reported that elevated IgG levels due to the presence of elevated periodontal pathogen species like P. gingivalis, T. denticola, S. noxia, A. actinomycetemcomitans, and V. parvula are connected to detrimental kidney function.291 Bastos et al. has reported that higher frequency of Candida albicans, P. gingivalis, T. forsythia, and T. denticola was associated with the development of chronic periodontitis in CKD patients, thereby indicating a bidirectional relationship between changes in oral microbiota and CKD.292 A large 10-year cohort study with CKD patients suffering with periodontitis had demonstrated an increase in mortality rate from 32% to 41% in those patients.293 However, data is lacking to establish a solid confirmation on the role of oral microbiota in the pathogenesis of CKD.
The gut-kidney axis functionality is based on metabolic and immune pathways being interlinked with each other.288 The metabolic pathway is mostly focused on gut microbiota-produced metabolites that mediate host physiological functions, whereas the immune pathway depends on several other components like monocytes, lymphocytes, and cytokines, which facilitate the communication between the gut and kidney.294 Recently, involvement of dysbiosis in the gut microbial environment leading to CKD has garnered attention, as there are implications of cross functionality between the gut and renal system.295 Numerous studies have been conducted to link the qualitative and quantitative changes in intestinal microbiota with the pathogenesis of CKD and end-stage renal disease (ESRD).296,297 However, there is no solid evidence confirming the presence of altered gut microbiota in CKD patients.297 Factors such as increased protein absorption, reduced dietary fiber intake, slower intestinal transit, and frequent oral intake of iron supplements and antibiotics resulted in altered intestinal microbial environment, leading to systemic inflammation and accumulation of uremic toxins. Both inflammation and uremic toxins substantially contribute to the progression of CKD and CKD-associated complications.298 Vaziri et al. showed that continuous loss of kidney function augments intestinal dysbiosis in CKD and ESRD patients.299 A comparative study between fecal samples comparing healthy subjects with CKD patients have exhibited that CKD patients show reduced abundance of Actinobacteria phylum and Akkermansia genera, where the latter is correlated with regulating levels of IL-10, denoting its importance in systemic inflammation.300 Another clinical study conducted using 73 subjects have identified 31 phylotypic differences between CKD and control groups with phylotypes like Bacteroides, Parabacteroides, R. gnavus, R. torques, Flavonifractor, Weissella, Ruminiclostridium, Erysipelatoclostridium, Eggerthella, and Sellimonas being predominant in CKD patients.301 ESRD patients have shown an increase in Actinobacteria, Proteobacteria and Firmicutes and a decrease in Bifidobacteria and Lactobacilli compared with the control group.299 Another study has demonstrated that changes in gut microbiota is also shown to be an important factor in contributing to inflammation along with oxidative stress by increasing accumulation of gut-derived uremic toxins such as indoxyl sulfate, amines, ammonia, p-cresyl glucuronide (PCG), p-cresyl sulfate (PCS) and TMAO in CKD patients.302 Dietary intervention is also an additional variable that induced post-translational modification of uremic toxins, indirectly contributing to CKD progression.303 Fiber-rich diet is a main contributor to colonic bacterial fermentation, and CKD patients often have a low fiber intake diet to limit the potassium intake.304 A meta-analysis study has reported that one-third of CKD patients exhibit higher levels of pathogenic bacteria like E. coli and Enterobacter, and mild CKD patients have shown increasing presence of uremic toxin-producing bacteria.305 In vivo studies using collagen type 4α3 (Col4a3)–deficient mice demonstrated that uremia is associated with intestinal dysbiosis and intestinal barrier dysfunction, causing persistent systemic inflammation in CKD.306 Human studies conducted with CKD patients have shown higher levels of PCS and PCG in general with PCS reaching levels around 200-fold higher than PCG.307,308 Mutsaers et al. have demonstrated that PCS and PCG affect renal tubular function while simultaneously affecting the activity of MRP4 (PCS and PCG) and BCRP (PCG) transporters.309 In vivo studies have shown that PCS-administered rats at a dose of 50 mg/kg for 4 weeks induced renal tubular cell damage.310 Various in vitro studies have also shown that indoxyl sulfate is responsible for inducing inflammatory and profibrotic responses in tubular cells.311,312 Increased levels of TMAO are associated with increased risk prediction of CVD, systemic inflammation, and mortality in CKD patients.313 A trial study using samples obtained from CKD patients displayed higher plasma levels of TMAO in CKD vs non-CKD patients.314 Persistent low-grade inflammation is augmented due to translocation of bacteria and bacterial products from the gut lumen to blood via increase in intestinal permeability. Decreased levels of certain microbiota metabolites like butyrate and vitamin K, which are nephroprotective, were also observed.308 These studies show strong evidence for involvement of various disturbances in gut-renal system communication via dysbiosis in microbiota in the progression and pathogenesis of kidney diseases.
Chronic liver diseases
Liver diseases remain one of the leading causes of morbidity and mortality worldwide. Nonalcoholic fatty liver disease/nonalcoholic steatohepatitis (NAFLD/NASH) and alcoholic liver disease (ALD) are the most common chronic liver diseases that often lead to liver cirrhosis and cancer.315 NAFLD comprises a wide span of liver damages from benign steatosis to steatohepatitis with hepatocellular inflammation and damage.315 ALD may take the form of chronic disease state (steatosis, steatohepatitis, fibrosis, or cirrhosis) or acute involvement (alcoholic hepatitis).316 Cirrhosis is the end stage of all chronic liver diseases, characterized by tissue fibrosis and the transformation of normal liver architecture to abnormal nodules. Recent studies have suggested the roles that oral and gut microbiota play in the pathogenesis of chronic liver diseases.
Gut microbiota in liver diseases
Mounting evidence supports the bidirectional gut-liver axis, due to the fact that liver secretes bile acids into the biliary tract and receives blood supply via the portal vein.317 Therefore, gut microbiota may contribute to liver diseases by delivering pathogens or metabolites into the liver through the portal vein. Currently, clinical data demonstrating the relationship between gut microbiota dysbiosis and liver diseases are still limited. Mouzaki et al. reported that patients with NASH have lower level of Bacteroidetes compared with healthy controls.318 Raman et al. suggested a compositional shift in the gut microbiota of obese NAFLD patients. Analysis of fecal microbiome showed an increased abundance of Lactobacillus and selected members of Firmicutes.319 Wong et al. reported an increased fecal abundance of Parabacteroides and Allisonella but decreased levels of Faecalibacterium and Anaerosporobacter.320 In liver cirrhosis patients, Chen et al. showed that abundance of Bacteroidetes was significantly reduced, while Proteobacteria and Fusobacteria were enriched compared with healthy controls.321 However, the investigation of gut microbiota with liver diseases is mainly conducted in preclinical studies. More evidence is required to conclude whether dysbiosis contributes to liver diseases or is a consequence of the disease state.
Researchers have postulated several mechanisms linking gut microbiota to liver diseases, including regulation of bile acid metabolism, intestinal permeability, chronic inflammation, and immune response. The gut microbiota plays an essential role in the metabolism of bile acids by converting primary bile acids into secondary bile acids. Deoxycholic acid, a major secondary bile acid, has been suggested to activate NF-κB stress response pathway by generating ROS.322 In addition, recent studies have established the crosstalk between ROS and NF-κB signaling pathway. While high ROS level usually results in cell damage, NF-κB pathway is known to promote cell proliferation.323 Therefore, it is likely that deoxycholic acid can induce cytotoxicity by promoting the generation of ROS, and simultaneously activates NF-κB pathway to allow damaged cells to resist apoptosis. Hence, microbiota dysbiosis may affect to bile acid homeostasis, leading to pathogenesis of chronic liver diseases such as NAFLD/NASH. Moreover, gut microbiota is involved in the metabolism of choline, and its deficiency usually leads to hepatic steatosis.324 There are multiple mechanisms established to explain choline deficiency and liver diseases, including 1) accumulation of DNA damage during choline depletion, 2) overproduction of free radicals in choline deficient hepatocytes, and 3) induction of inflammatory response due to death of hepatocytes.324 Spencer et al. showed that the abundance of Gammaproteobacteria and Erysipelotrichi were significantly associated with choline deficiency-induced fatty liver.325 Impaired intestinal permeability allows the translocation of gut bacteria and their component, which is associated with chronic liver diseases. For example, the Gram-negative bacteria structural element LPS was suggested to be elevated in portal vein in ALD and cirrhosis.326 Mechanistically, Seki et al. described that, upon LPS binding, TLR4 upregulates chemokine secretion and downregulates TGF-β pseudoreceptor Bambi to enhance TGF-β signaling pathway.327 The study also suggested that the effect of LPS is mediated by MyD88-NF-κB-dependent pathway, as MyD88-deficient mice had decreased hepatic fibrosis. Gut microbiota-mediated chronic inflammation and immune activation is central to the pathogenesis of multiple diseases. Recent studies also suggested such mechanisms in the development of NAFLD/NASH.328 In conclusion, current research highlights the potential role of gut microbiota in liver diseases, but further study is needed to confirm their relationship.
Oral microbiota in liver diseases
As mentioned in other diseases, it has been demonstrated that oral microbes or their metabolites are able to invade other sites of the body. Although the correlation between periodontal and liver diseases is yet to be established, recent studies implicated that periodontal bacteria may be involved in the progression of NAFLD, NASH, and cirrhosis.329 Yoneda et al. reported that P. gingivalis (one of the most common periodontal pathogens) may influence the pathogenesis of NAFLD/NASH in a mouse model.330 In addition, they found that P. gingivalis infection was mostly observed in NAFLD patients compared with control subjects. Clinical data also support the notion that periodontitis may serve as a risk factor in the progression of NAFLD/NASH.331
Mechanistically, this process may be attributed to many factors including pro-inflammatory mediators, oxidative stress, and pathogen invasion. The migration of periodontal bacteria as well as their metabolites (LPS, peptidoglycans, etc.) into the systemic circulation is usually recognized by TLRs, which leads to the activation of T cells and the release of pro-inflammatory cytokines, chemokines, and ROS/RNS.332 This may indicate the oral-gut-liver axis in the inflammation pathway as suggested by Acharya et al.333 They proposed that the gut with impaired intestinal permeability may act as an intermediate between oral microbiota and the liver. Therefore, after the bacteria and metabolites enter the systemic circulation, they can reach the liver and bind to innate TLRs of hepatocytes and Kupffer cells, inducing inflammation and causing liver diseases.334 In addition, Silva Santos et al. observed that cirrhotic patients exhibited numerous oral diseases other than periodontitis, such as candidiasis, xerostomia, and petechiae.335 Moreover, dysbiosis of oral microbiota has been suggested to promote the pathogenesis of hepatitis B virus (HBV)-induced chronic liver disease. HBV-associated oral bacteria including Fusobacterium, Eubacterium, and Treponema may invade and contribute to the dysbiosis of gut microbiota as opportunistic pathogens, which subsequently participate in the formation of liver diseases.336
Microbiota and disease treatment
With the gradual understanding of microbiota, the potential of treating diseases through manipulating microbiota has attracted people’s attention. Because the human gut is involved in a wide range of physiologic functions, its modulation is expected to prevent or treat the corresponding diseases. Therefore, climbing number of clinical trials are ongoing to investigate this possibility (Fig. 5a). The majority of clinical trials focusing on efficacy of fecal microbiota transplantation (FMT) is various diseases. Since C.Difficile infection, cancer, and IBD has the highest number of trials, we also summarized the data from pubmed (Supplemental Table S1). As shown in Fig. 5b, c, FMT treatment in IBD and C.Difficile infection showed a significant response rate compared to placebo treatment. Similarly, probiotics treatment as an adjuvant therapy in cancer patients also demonstrated an optimal result (Fig. 5d). It should be noted that while the response rate is promising, the trials are mainly pilot studies with small sample size. In addition, the underlying mechanism for complete response required further investigation to optimize the experimental design and to personalize the treatment. Diet is considered the main short-term and long-term regulator of the gut microbiota,337 along with healthy lifestyle habits. As shown in Fig. 6, this section will present the latest clinical interventions targeting the gut microbiota, including microbiota modulations, FMT, and bacteria engineering. We will also discuss the pharmacological microbiota–drug interactions in clinical settings.

Current number of microbiota-related clinical trials by regions and study phases. Data are updated until October 2021

Strategies to modify gut microbiota for disease treatment
Microbiota modulations
Probiotics and prebiotics
Generally, probiotics and prebiotics are the most popular topic in microbiota modulation research. They are often used as a dietary supplement for clinical intervention by oral administration. Differences in dosage form and host are considered to be the main factors affecting the effectiveness of probiotics and prebiotics.338 Probiotic administration is suggested to restore microbial dysbiosis and maintain intestinal microbial balance by occupying host tissue and preventing colonization of pathogenic bacteria. Extensive research has indicated the potential mechanism of probiotics in disease treatments, including differences in various probiotic strains and mucosal immune system, regulation of host metabolism or altering intestinal neuromuscular function.339 However, clinical data is insufficient to support their role. Several clinical trials denied the benefits of probiotics in cancer treatment.340 Importantly, patients with damaged intestinal barrier and/or compromised immune systems might have a probiotic translocation. Some published case reports associate negative effects of probiotics with conditions such as bacteremia, fungemia, endocarditis, liver abscess and pneumonia,341 which compels us to ponder the actual effects of probiotics.
Prebiotic was recently redefined as “a substrate that is selectively utilized by host microorganisms conferring a health benefit” in 2017. Prebiotics were originally used to study the stimulating effect of probiotics. The most well-known prebiotics are inulin, fructo-oligosaccharides (FOS), lactulose, and galacto-oligosaccharides (GOS). Prebiotics are mainly used to modulate the strains of Bifidobacterium and Lactobacillus, which produce lactic acid and acetate, and to maintain the health of the host by fermenting prebiotics.342 Studies have confirmed that taking prebiotics can stimulate the selective enrichment of probiotics in the intestinal tract, thereby regulating immune response and preventing pathogens.70 Although some basic studies have confirmed that prebiotics can inhibit the colonization of pathogens by mimicking glycoconjugates of microvilli,343 and even directly act on the intestinal tract to regulate immunity,344 the applicability of prebiotics as a clinical intervention is still debatable. Belcheva et al. suggested that supplementation with prebiotic/butyrate could promote tumor progression due to genetic variation in individuals.345 Their findings suggest that butyrate functions as an oncometabolite, while a substantial of studies reported butyrate as tumor suppressive metabolite. Indeed, butyrate is known to exhibit differential effects toward normal and cancerous colonocytes. In colon cancer cells, butyrate is metabolized to a lesser extent compared to normal cells, thereby accumulating as HDAC inhibitor to inhibit cell proliferation and induce cell apoptosis.346 The difference observed in this study may lay between host genetic background, the age, and the presence of other bacterial metabolites. In addition, the study was performed in mice model, so the translation to human requires further investigations.
Sasaki et al. showed that transglucosidase, which generates prebiotics, can reduce high blood sugar levels in patients with T2DM and inhibit weight gain.347 Because probiotics and prebiotics are cheap and easy to handle, they are often used in the care of patients with AD. Long-term supplementation with milk enriched with Bifidobacteria and Lactobacillus fermentum improves learning and memory in AD patients.348 In CRC treatment, some probiotic strains could be beneficial as an adjuvant therapeutic agent, such as multigene and multistrain probiotics, including B. breve, B. infantis, B. longum, L. acidophilus.349,350 Recent studies in intestinal inflammatory disorders show that probiotics might have some efficacy in UC and pouchitis, but with insignificant effect in CD. Probiotic supplementation may significantly reduce rates of rotavirus diarrhea, although the curative effect of probiotics in NSAID enteropathy and IBS is controversial.351 This is because the study populations, types of probiotics and dosage and length of follow-up various greatly between the clinical studies. Similarly, the treatment with synbiotics and FMT demonstrated controversial results due to the limited data. In heart diseases, an in vivo study showed that rats treated with probiotics and prebiotics containing Lactobacillus plantarum 299 v could reduce infarct size and improve left ventricular function before coronary artery ligation.352 Gan et al. demonstrated similar cardioprotective results in a rat model of myocardial ischemia after supplementation with Lactobacillus rhamnosus GR-1.353 In addition, modulation of the gut microbiota through probiotics may present potential therapeutic strategies to protect against lung diseases.354 Moreover, probiotics is believed to have some positive effect on COVID-19 treatment. For example, Chen et al. suggested that probiotics could reduce hyperinflammation from COVID-19 through its anti-inflammatory effects.355 However, a systematic review by Bafeta et al., which investigated 384 RCTs, found that the report of side effects in published RCTs assessing probiotics, prebiotics, and symbiotic is often lacking or inadequate.356 Therefore, the safety of these interventions cannot be determined without enough safety data.
Antibiotic
Antibiotic administration is the most common approach to manipulate the composition of the gut microbiota. Researchers found that modulation of gut microbiota by antibiotics improves insulin signaling in high fat-fed mice.357 In a study of melanoma and lung cancer models, vancomycin can enhance the anti-tumor response induced by radiotherapy in mice by increasing CD8 + T cell infiltration and IFN-γ expression.358 Many studies have shown that antibiotics can prevent cancer development or attenuate tumor proliferation. For example, Bullman et al. showed that metronidazole treatment can eradicate the colonization of Fusobacterium and ameliorate the progression of CRC.359 The results showed that colonization of Fusobacterium with CRC tumor cells was maintained in distal metastasis, demonstrating a stable microbiome composition between primary and metastatic tumors. In addition, antibiotic treatment that reduced Fusobacterium load also inhibited cancer cell proliferation and tumor growth. Therefore, this finding suggests the potential of microbiota modulation, i.e., antibiotic intervention, for patients with microbial-associated cancer. However, we cannot exclude the possibility that broad spectrum antibiotics may have negative impact on the healthy intestinal microbiota, therefore the use of antimicrobial agent targeting to the specific bacteria is highly important. At the same time, antibiotics also showed potential as immunotherapy. In a metastatic mouse model, antibiotic consumption of the gut microbiota could inhibit tumor growth by triggering an anti-tumor immune response.360 The researchers studied the effect of antibiotics on tumor growth of pancreatic cancer, CRC, and melanoma. It is found that gut microbiota depletion by oral microbiota significantly reduced the tumor growth in all tumor models. Interestingly, the inhibition effect was not observed in Rag-1 KO mice, which lack mature B and T cells, suggesting the effect may be dependent on host immunity. Indeed, the mechanistic study showed that gut microbiota depletion resulted in a significant increase in IFγ producing T cells and the decrease in IL-17A/IL-10 producing T cells. In addition, gut microbiota depletion led to infiltration of effector-T cells into pancreatic tumors. Previous studies have shown that immune checkpoint inhibitors failed to antagonize pancreatic cancer due to low effector-T cell infiltration. Hence, the antibiotic treatment may be beneficial as an adjuvant therapy along with conventional immunotherapy. In patients with early gastric cancer, the eradication of H. pylori through the combination of amoxicillin and clarithromycin is associated with a lower incidence of metachronous gastric cancer and improvement of the degree of gastric gland atrophy.361
It remains controversial that, although antibiotics can effectively eradicate pathogens or harmful bacteria, their non-selective antibacterial effects may kill the symbiotic microbes, leading to another ecological disorder. It may also impair the efficacy of cancer immunotherapy and lead to treatment resistance. Vétizou et al. demonstrated that the efficacy of CTLA-4 blockade was associated with the T cell response for B. fragilis in mice and patients. Moreover, while GF mice were not responding to CTLA-4 blockade, introduction of B. fragilis was able to overcome this defect.362 Therefore, this study suggests a key role of microbiota in triggering the response to immunotherapy and it is meaningful to explore if other bacteria have the similar functions. Hernández et al. reported that compared with untreated individuals, subjects receiving antibiotics showed greater or unbalanced sugar anabolic capacity.363 Clinical studies have shown that antibiotics are closely related to the increased risk of CRC development,364,365 though this result may be affected by confounding indications.366 For example, compared with cancer patients, immunodeficient patients are more susceptible to bacterial infections that require antibiotic treatment.364 Despite the controversy surrounding the use of antibiotics, their potential for microbiota regulation should not be underestimated. With the development of modern sequencing methods, we can have a more comprehensive understanding of the impact of antibiotics on microbial communities, thereby bringing new vitality to the use of antibiotics.
Fecal microbiota transplantation
FMT refers to a method of introducing a solution of fecal matter from a donor into the intestinal tract of a recipient to cure disease. FMT treatment, which was first documented in China in the 4th century,367 will change the recipient’s microbial composition directly. The most prominent results in the use of fecal transplantation for disease treatment have been in the treatment of recurrent Clostridium difficile infection (rCDI), with reported cure rates near 90%.368 In 2013, FMT was approved by the FDA to treat rCDI. FMT methods include the use of a naso-intestinal tube, gastroscopy, and colonoscopy, all with different efficacies. A meta-analysis conducted by Laniro et al. showed that capsule FMT has an overall response rate of more than 90% and is minimally invasive.369
So far, more than 100 case reports and clinical trials of FMT for rCDI have been published; most reports have high resolution of diarrhea associated with rCDI. Several meta-analyses have confirmed that FMT is superior to standard antibiotic therapy. They also showed that FMT is a safe treatment method for patients with rCDI.370 Compared with the traditional therapy of vancomycin regimen which is only 31% effective, FMT therapy showed a cumulative effectiveness of 94%.371 The clinical remission rate of FMT therapy in the RCT study of UC is about 36–37%. FMT is also widely investigated in the treatment of cancer,372 diabetes,373 ASD,266 multiple sclerosis,374 atherosclerosis and hypertension,375 graft vs host disease,376 Parkinson’s disease,377 hepatic encephalopathy and NAFLD.378 Although these treatments showed promising results, they were investigated in preclinical models, or the sample sizes were too small. Therefore, extensive studies are required before drawing further conclusions.
Currently, there are several mechanisms proposed for FMT including the following: 1) FMT may stimulate decolonization of pathogenic microbes and enhance host resistance to pathogens by direct ecological competition;379 2) repopulating gut microbiota by FMT helps to restore immune function and reduce host damage induced by abnormal microbial colonization of the gastrointestinal tract; 3) FMT facilitates the restoration of essential metabolites used for host metabolism, including SCFAs, antimicrobial peptides, bacteriocins, and bile acids.380 FMT is safe to a large extent, and large studies report mainly minor, short-lived adverse reactions. The specific high-risk population is mainly immunocompromised patients.381 But this therapy still faces many challenges. For example, regarding the standardization of donor screening, eligible stool donors are often rare if considering the risk of infection. Starting from the selection of a donor to the route of administration and dynamic monitoring after FMT treatment, the entire FMT process requires the use of personalized methods to reach its full potential. Therefore, the future development direction of FMT may be in precision medicine.382
Engineering gut bacteria
Most bacteria that coexist with humans are nonpathogenic. Advances in modern DNA technology have made it possible to engineer bacteria for disease treatment. Based on traditional genetic engineering methods, engineered probiotics have been used in the treatment of colitis, diabetes, obesity, and a large number of pathogenic infections.383,384,385 Lactobacillus jannaschii (a conventional flora of the female vagina) has been modified to secrete HIV-resistant cyanovirin-N protein. This engineered bacteria has been proven to reduce HIV infection by 63% in rhesus monkeys.386 There are different types of engineered bacterial therapies for diseases, such as synthetic immune regulatory proteins, chemotactic response systems, and protein delivery systems.
“Smart probiotics” created using genetic engineering technology brings vitality to the application of probiotics. It has a better efficacy than natural probiotics. For example, Lactococcus lactis expressing human Trefoil Factor 1 (a cytopeptide involved in epithelial wound healing) has been formulated as a mouthwash for the treatment of oral mucositis.387 A combination therapy of engineered Lactococcus lactis has been used in a clinical trial of T1DM treatment.388 Moreover, an engineered Lactococcus lactis strain which secretes the anti-inflammatory cytokine IL-10 showed a clinical benefit in CD.389 Insulin production in epithelial cells can be induced by the gut hormone GLP.390 Duan et al. reported that an engineered GLP-1-secreting Lactobacillus gasseri strain can reprogram intestinal cells into insulin-secreting cells.391
Bacteria can bypass problems associated with poor selectivity and limited tumor penetrability of conventional cancer chemotherapies and can be finely engineered to sense and respond to the tumor microenvironment.392 One strategy is to utilize the native bacterial cytotoxicity to kill cancer cells. For example, Clostridium and Salmonella has exhibit anticancer effect in mice models. Accumulation of the bacteria in tumor tissues will induce neutrophil infiltration and antitumor immune response. Such response was also observed in phase 1 clinical trial that administrated a modified Salmonella strain to patients with metastatic melanoma.393 The second strategy is using engineered bacteria to directly express anticancer agents or transfer eukaryotic expression vectors into cancer cells.394 With these approaches, the bacteria can 1) generate cytotoxic agents such as Cytolysin A to induce cancer cell apoptosis, 2) deliver cytokines such as IL-2, TNFSF14 that activate immune cells to eradicate cancer cells, and 3) sensitizing immune system against cancer cells by expressing tumor antigen. The third strategy is using bacteria to transfer genetic material to cancer cells. Therefore, it stimulates competition with the mechanisms that foster tumor formation, through the in-situ delivery of polypeptides with pro-apoptotic activity, anti-angiogenic factors, and cytokines. Bacteria have also been engineered to silence the expression of important genes related to tumor development through RNA interference. For instance, Xiang et al. reported that E. coli can be engineered to transfect host cells with plasmids encoding short-hairpin RNAs (shRNAs) silencing catenin beta-1, whose overexpression is involved in several types of cancer.395 This therapy has been granted orphan drug status by the FDA for the treatment of familial adenomatous polyposis and is currently under clinical trial investigation to analyze the safety and tolerability.
Gene-editing technology such as CRISPR has broadened the application of engineered bacteria in microbiota modulation. CRISPR is being utilized in the development of novel antimicrobial strategies. Hwang et al. showed that the exonuclease CRISPR-associated protein 3 (Cas3) can be engineered into a probiotic, which has the capacity to efficiently kill pathogenic bacteria.396 A bacterial protein secretion system (T3SS) can transfer proteins into the cytoplasm of infected cells. With this system, engineered bacteria can carry polypeptide vaccines or proteins into host cells and carry transcription factors into the cell. In addition, dysregulation of the microbiome can lead to cytokine storms, which may be associated with a decrease in angiotensin 2 (ACE2).397 Based on this, Verma et al. developed an expression and delivery system (LP) using probiotic species Lactobacillus piracies as a live vector for oral delivery of human ACE2. It provides a new strategy for correcting the imbalance of the gut microbiota while increasing the serum ACE2 level.398 It is true that the way in which a given supplement or drug affects the microbiota-host interface is obviously not enough for the complex human environment. Awareness of the range of possible interactions between the intervention and the host’s diet, genome, immune system, and resident symbionts should be taken into consideration. Although the clinical application of new technologies, such as T3SS and CRISPR, still require more investigation, they provide more opportunities and possibilities for microbiota therapy in the future.
Gut microbiota and drug response
It is well established that drug response, mainly characterized by pharmacokinetic (PK) and pharmacodynamic (PD) properties, may differ among individuals due to factors such as gender, age, and genetic variations.399 However, it was only recently that researchers identified microbiota as a mediator of drug response, highlighting its role in medical therapy. We herein describe the role of gut microbiota on modulating drug effect as it is a current research focus.
It is known that the gut microbiota can metabolize a wide range of substances, which can have potential implications for affecting drug absorption. Particularly, the stability of orally administered drugs can be affected in the GI tract before entering the systemic circulation. Sousa et al. summarized that over thirty drugs are substrates of bacterial enzymes in the distal gut.400 Recently, it was suggested that small intestine microbiota may also have profound impact on host physiological functions.401 This finding highlighted the potential drug-microbiota interaction, since small intestine is a major site for drug absorption. Indeed, numerous studies have reported altered drug PK mediated by gut microbiota with clinical implications. For example, Sun et al. reported that a hypoxic environment can affect the composition of gut microbiota, which led to increased absorption of aspirin in rats.402 Matuskova et al. identified that concomitant orally administrated probiotic E. coli strain Nissle 1917 (EcN) affected the PK of the antiarrhythmic drug amiodarone in male rats.403 EcN increased the plasma level of amiodarone metabolites, probably due to increasing the drug absorption or the activity of CYP2C enzymes, which was not observed in the reference non-probiotic strain.
Wallace et al. suggested that β-glucuronidases from E. coli can metabolize irinotecan into the active metabolite SN-38 in intestinal lumen and damage the intestinal epithelium in a mouse model.404 Lindenbaum et al. reported that the most widely used cardiac glycoside digoxin can be converted to reduced derivatives produced by Eubacterium lentum, a common gut anaerobe.405 The same research group reported a follow-up study that showed that administration of antibiotics erythromycin or tetracycline was able to significantly reduce the levels of digoxin, reduced metabolites, and increase serum digoxin level to a maximum of 2-fold. Wu et al. established a pseudo-GF diabetic rat model to investigate the relationship between metformin and gut microbiota.406 They found that the antihyperglycemic effect of metformin was reduced by more than 40% in gut microbiota-depleted group. Moreover, the hepatoprotective effect of metformin was significantly reduced in the absence of gut microbiota. Recent studies in IBD revealed that gut microbiota may influence the metabolism of IBD drugs mesalazine, methotrexate, thioguanine, and glucocorticoids.407 In particular, thioguanine can be converted to its active form by gut bacteria without the requirement of host metabolism. Studies also suggested that microbiota can affect host metabolism by modulating cytochrome P450 enzymes and UDP-glucuronosyltransferase.408 Interestingly, a recent study by Klünemann et al. established multiple new bacteria-drug interactions, with more than half of them ascribed to bioaccumulation,409 phenomenon by which bacteria can store the drug without chemical modification, thereby altering host drug response. Particularly, the behavioral response of Caenorhabditis elegans to antidepressant duloxetine was attenuated by bioaccumulating bacteria such as S. salivarius.
A large number of data confirmed that the gut microbiota can have a major impact on drug PK and subsequently the drug response in clinical settings. A better understanding of such interaction is required to develop effective treatment strategies.
Conclusion
After decades of research, we have gradually established a new role of microbiota in health and disease. It is now confirmed that microbiota can affect almost all aspects of the host, while its dysbiosis is related to a wide spectrum of diseases. Thanks to advanced research technologies, we are able to closely examinate how microbiota maintain human health and contribute to pathogenesis. However, the study of microbiota is mainly focused on the bacterial component; the role of fungi, viruses, and other microbes in health and disease remain largely inconclusive. In addition, while microbiota dysbiosis is often observed in disease states, the causative role of microbiota is yet to be established. Hence, there are still a lot of questions to be answered in this field. The greater understanding of this host-microbiota relationship has allowed for the development of microbiota-based therapy such as FMT and bacteria modulation. These strategies are well on the way to achieving the optimal clinical effect in the treatment of C. difficile infection, diabetes, inflammatory bowel disease, etc. In summary, we are now in a better position to treat diseases and foster health via manipulation of the microbial symbionts.
References
Ursell, L. K. et al. The intestinal metabolome: an intersection between microbiota and host. Gastroenterology 146, 1470–1476 (2014).
Grice, E. A. & Segre, J. A. The human microbiome: our second genome. Annu. Rev. Genom. Hum. Genet. 13, 151–170 (2012).
Berg, G. et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome 8, 103 (2020).
Shreiner, A. B., Kao, J. Y. & Young, V. B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 31, 69–75 (2015).
Hillman, E. T., Lu, H., Yao, T. & Nakatsu, C. H. Microbial ecology along the gastrointestinal tract. Microbes Environ. 32, 300–313 (2017).
Laterza, L. et al. The gut microbiota and immune system relationship in human graft-versus-host disease. Mediterranean J. Hematol. Infect. Dis. 8, 2016025 (2016).
Auchtung, T. A. et al. Investigating colonization of the healthy adult gastrointestinal tract by fungi. mSphere. 3, e00092–18 (2018).
Lozupone, C. A. et al. Diversity, stability and resilience of the human gut microbiota. Nature 489, 220–230 (2012).
Deo, P. N. & Deshmukh, R. Oral microbiome: unveiling the fundamentals. J. Oral. Maxillofac. Pathol. 23, 122–128 (2019).
Segata, N. et al. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol. 13, R42 (2012).
Dickson, R. P. & Huffnagle, G. B. The lung microbiome: new principles for respiratory bacteriology in health and disease. PLoS Pathog. 11, e1004923 (2015).
Dickson, R. P., Martinez, F. J. & Huffnagle, G. B. The role of the microbiome in exacerbations of chronic lung diseases. Lancet 384, 691–702 (2014).
Tong, X. et al. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional Chinese herbal formula: a multicenter, randomized, open label clinical trial. mBio. 9, e02392–17 (2018).
Ley, R. E., Turnbaugh, P. J., Klein, S. & Gordon, J. I. Human gut microbes associated with obesity. Nature 444, 1022–1023 (2006).
Bouskra, D. et al. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature 456, 507–510 (2008).
Turnbaugh, P. J. et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027–1031 (2006).
Roberfroid, M. B., Bornet, F., Bouley, C. & Cummings, J. H. Colonic microflora: nutrition and health. Summary and conclusions of an International Life Sciences Institute (ILSI) [Europe] workshop held in Barcelona, Spain. Nutr. Rev. 53, 127–130 (1995).
Fan, Y. & Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 19, 55–71 (2021).
Flint, H. J., Scott, K. P., Louis, P. & Duncan, S. H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 9, 577–589 (2012).
Milani, C. et al. The first microbial colonizers of the human gut: composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 81, e00036–00017 (2017).
Rinninella, E. et al. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms. 7, 14 (2019).
Amabebe, E., Robert, F. O., Agbalalah, T. & Orubu, E. S. F. Microbial dysbiosis-induced obesity: role of gut microbiota in homoeostasis of energy metabolism. Br. J. Nutr. 123, 1127–1137 (2020).
Yatsunenko, T. et al. Human gut microbiome viewed across age and geography. Nature 486, 222–227 (2012).
Guigoz, Y., Doré, J. & Schiffrin, E. J. The inflammatory status of old age can be nurtured from the intestinal environment. Curr. Opin. Clin. Nutr. Metab. Care. 11, 13–20 (2008).
Hugenholtz, F. & de Vos, W. M. Mouse models for human intestinal microbiota research: a critical evaluation. Cell Mol. Life Sci. 75, 149–160 (2018).
Cheng, Y. et al. Principles of regulatory information conservation between mouse and human. Nature 515, 371–375 (2014).
Park, J. C. & Im, S.-H. Of men in mice: the development and application of a humanized gnotobiotic mouse model for microbiome therapeutics. Exp. Mol. Med. 52, 1383–1396 (2020).
Krych, L. et al. Quantitatively different, yet qualitatively alike: a meta-analysis of the mouse core gut microbiome with a view towards the human gut microbiome. PLoS One 8, e62578 (2013).
Bohnhoff, M., Drake, B. L. & Miller, C. P. The effect of an antibiotic on the susceptibility of the mouse’s intestinal tract to Salmonella infection. Antibiotics Annu. 3, 453–455 (1955).
Coyte, K. Z. & Rakoff-Nahoum, S. Understanding competition and cooperation within the mammalian gut microbiome. Curr. Biol. 29, R538–R544 (2019).
Martens, E. C., Chiang, H. C. & Gordon, J. I. Mucosal glycan foraging enhances fitness and transmission of a saccharolytic human gut bacterial symbiont. Cell Host Microbe 4, 447–457 (2008).
Ducarmon, Q. R. et al. Gut Microbiota and colonization resistance against bacterial enteric infection. Microbiol. Mol. Biol. Rev. 83, e00007–19 (2019).
Freter, R. et al. Mechanisms that control bacterial populations in continuous-flow culture models of mouse large intestinal flora. Infect. Immun. 39, 676–685 (1983).
Kashyap, P. C. et al. Genetically dictated change in host mucus carbohydrate landscape exerts a diet-dependent effect on the gut microbiota. Proc. Natl Acad. Sci. USA 110, 17059–17064 (2013).
Pukatzki, S. et al. Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl Acad. Sci. USA 103, 1528–1533 (2006).
Russell, A. B. et al. Type VI secretion delivers bacteriolytic effectors to target cells. Nature 475, 343–347 (2011).
Anderson, M. C. et al. Shigella sonnei encodes a functional T6SS used for interbacterial competition and niche occupancy. Cell Host Microbe 21, 769–776.e763 (2017).
Whitney, J. C. et al. A broadly distributed toxin family mediates contact-dependent antagonism between gram-positive bacteria. eLife 6, e26938 (2017).
Davies, E. V., Winstanley, C., Fothergill, J. L. & James, C. E. The role of temperate bacteriophages in bacterial infection. FEMS Microbiol. Lett. 363, fnw015 (2016).
Furuyama, N. & Sircili, M. P. Outer Membrane Vesicles (OMVs) produced by gram-negative bacteria: structure, functions, biogenesis, and vaccine application. BioMed. Res. Int. 2021, 1490732 (2021).
Loenen, W. A. M. & Raleigh, E. A. The other face of restriction: modification-dependent enzymes. Nucleic Acids Res. 42, 56–69 (2014).
Amitai, G. & Sorek, R. CRISPR-Cas adaptation: insights into the mechanism of action. Nat. Rev. Microbiol. 14, 67–76 (2016).
Goldfarb, T. et al. BREX is a novel phage resistance system widespread in microbial genomes. EMBO J. 34, 169–183 (2015).
Sobel, N. Is there a protective role for vaginal flora? Curr. Infect. Dis. Rep. 1, 379–383 (1999).
Fredricks, D. N., Fiedler, T. L. & Marrazzo, J. M. Molecular identification of bacteria associated with bacterial vaginosis. N. Engl. J. Med. 353, 1899–1911 (2005).
Ravel, J. et al. Vaginal microbiome of reproductive-age women. Proc. Natl Acad. Sci. USA 108, 4680–4687 (2011).
Gajer, P. et al. Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 4, 132ra152 (2012).
Boskey, E. R. et al. Acid production by vaginal flora in vitro is consistent with the rate and extent of vaginal acidification. Infect. Immun. 67, 5170–5175 (1999).
Watts, D. H. et al. Effects of bacterial vaginosis and other genital infections on the natural history of human papillomavirus infection in HIV-1-infected and high-risk HIV-1-uninfected women. J. Infect. Dis. 191, 1129–1139 (2005).
Peipert, J. F. et al. Bacterial vaginosis, race, and sexually transmitted infections: does race modify the association? Sexually Transmitted Dis. 35, 363–367 (2008).
Cherpes, T. L. et al. Association between acquisition of herpes simplex virus type 2 in women and bacterial vaginosis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 37, 319–325 (2003).
Schwebke, J. R. & Desmond, R. A randomized trial of metronidazole in asymptomatic bacterial vaginosis to prevent the acquisition of sexually transmitted diseases. Am. J. Obstet. Gynecol. 196, 517.e511–516 (2007).
Brotman, RebeccaM. et al. Bacterial vaginosis assessed by gram stain and diminished colonization resistance to incident gonococcal, chlamydial, and trichomonal genital infection. J. Infect. Dis. 202, 1907–1915 (2010).
Leyden, J. J., McGinley, K. J., Mills, O. H. & Kligman, A. M. Propionibacterium levels in patients with and without acne vulgaris. J. Investig. Dermatol. 65, 382–384 (1975).
Christensen, G. J. M. & Brüggemann, H. Bacterial skin commensals and their role as host guardians. Beneficial Microbes 5, 201–215 (2014).
Iwase, T. et al. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 465, 346–349 (2010).
Belkaid, Y. & Segre, J. A. Dialogue between skin microbiota and immunity. Science 346, 954–959 (2014).
Cryan, J. F. et al. The microbiota-gut-brain axis. Physiol. Rev. 99, 1877–2013 (2019).
Farmer, A. D. & Aziz, Q. Mechanisms and management of functional abdominal pain. J. R. Soc. Med. 107, 347–354 (2014).
Chen, K. et al. Drosophila histone demethylase KDM5 regulates social behavior through immune control and gut microbiota maintenance. Cell Host Microbe 25, 537–552.e538 (2019).
Bravo, J. A. et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl Acad. Sci. 108, 16050–16055 (2011).
Buffington, S. A. et al. Microbial reconstitution reverses maternal diet-induced social and synaptic deficits in offspring. Cell 165, 1762–1775 (2016).
Poutahidis, T. et al. Microbial symbionts accelerate wound healing via the neuropeptide hormone oxytocin. PLoS One. 8, e78898 (2013).
de Theije, C. G. M. et al. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain, Behav. Immun. 37, 197–206 (2014).
Gilbert, J. A. Social behavior and the microbiome. eLife 4, e07322 (2015).
Allen, A. P. et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 6, e939 (2016).
Sampson, T. R. et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 167, 1469–1480.e1412 (2016).
Tomova, A. et al. Gastrointestinal microbiota in children with autism in Slovakia. Physiol. Behav. 138, 179–187 (2015).
Comprehensive Physiology. 1 edn, (Wiley, 2011).
Maier, T. V. et al. Impact of dietary resistant starch on the human gut microbiome, metaproteome, and metabolome. mBio. 8, e01343–17 (2017).
Wall, R. et al. in Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease Vol. 817 (eds Mark Lyte & John F. Cryan) 221–239 (Springer New York, 2014).
Jaglin, M. et al. Indole, a signaling molecule produced by the gut microbiota, negatively impacts emotional behaviors in rats. Front. Neurosci. 12, 216 (2018).
Perez-Burgos, A. et al. Psychoactive bacteria Lactobacillus rhamnosus (JB-1) elicits rapid frequency facilitation in vagal afferents. Am. J. Physiol.-Gastrointest. Liver Physiol. 304, G211–G220 (2013).
Sgritta, M. et al. Mechanisms underlying microbial-mediated changes in social behavior in mouse models of autism spectrum disorder. Neuron 101, 246–259.e246 (2019).
De Vadder, F. et al. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl Acad. Sci. 115, 6458–6463 (2018).
Mayer, E. A. The neurobiology of stress and gastrointestinal disease. Gut 47, 861–869 (2000).
Sudo, N. et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice: commensal microbiota and stress response. J. Physiol. 558, 263–275 (2004).
Dinan, T. G. et al. Hypothalamic-pituitary-gut axis dysregulation in irritable bowel syndrome: plasma cytokines as a potential biomarker? Gastroenterology 130, 304–311 (2006).
Wang, B. et al. The human microbiota in health and disease. Engineering 3, 71–82 (2017).
Hoban, A. E. et al. Microbial regulation of microRNA expression in the amygdala and prefrontal cortex. Microbiome 5, 102 (2017).
Chen, J.-j et al. Effects of gut microbiota on the microRNA and mRNA expression in the hippocampus of mice. Behav. Brain Res. 322, 34–41 (2017).
Hildebrandt, M. A. et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology 137, 1716–1724.e1712 (2009).
Biagi, E. et al. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS One. 5, e10667 (2010).
Devkota, S. et al. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 487, 104–108 (2012).
Karstens, A. J. et al. Associations of the Mediterranean diet with cognitive and neuroimaging phenotypes of dementia in healthy older adults. Am. J. Clin. Nutr. 109, 361–368 (2019).
Sánchez-Villegas, A., Henríquez, P., Bes-Rastrollo, M. & Doreste, J. Mediterranean diet and depression. Public Health Nutr. 9, 1104–1109 (2006).
Cavaleri, F. & Bashar, E. Potential synergies of β -hydroxybutyrate and butyrate on the modulation of metabolism, inflammation, cognition, and general health. J. Nutr. Metab. 2018, 1–13 (2018).
Olson, C. A. et al. The gut microbiota mediates the anti-seizure effects of the ketogenic diet. Cell 173, 1728–1741.e1713 (2018).
Clarke, G. et al. Gut reactions: breaking down xenobiotic–microbiome interactions. Pharmacol. Rev. 71, 198–224 (2019).
Falony, G. et al. Population-level analysis of gut microbiome variation. Science 352, 560–564 (2016).
Zhernakova, A. et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 352, 565–569 (2016).
Jin, M. et al. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ. Int. 120, 421–430 (2018).
Alexander, K. L., Targan, S. R. & Elson, C. O. 3rd Microbiota activation and regulation of innate and adaptive immunity. Immunol. Rev. 260, 206–220 (2014).
Hapfelmeier, S. et al. Reversible microbial colonization of germ-free mice reveals the dynamics of IgA immune responses. Science 328, 1705–1709 (2010).
Zhao, Q. & Elson, C. O. Adaptive immune education by gut microbiota antigens. Immunology 154, 28–37 (2018).
Owaga, E. et al. Th17 cells as potential probiotic therapeutic targets in inflammatory bowel diseases. Int. J. Mol. Sci. 16, 20841–20858 (2015).
Sekirov, I., Russell, S. L., Antunes, L. C. M. & Finlay, B. B. Gut microbiota in health and disease. Physiol. Rev. 90, 859–904 (2010).
Salzman, N. H. et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nat. Immunol. 11, 76–82 (2010).
Maynard, C. L., Elson, C. O., Hatton, R. D. & Weaver, C. T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 489, 231–241 (2012).
Bager, P., Wohlfahrt, J. & Westergaard, T. Caesarean delivery and risk of atopy and allergic disease: meta-analyses. Clin. Exp. Allergy. J. Br. Soc. Allergy. Clin. Immunol. 38, 634–642 (2008).
Durko, T., Gryczyński, M. & Dabrowski, R. Evaluation of collagenous protein levels in inflamed middle ear mucosa. Otolaryngologia Pol. = Pol. Otolaryngol. 45, 208–212 (1991).
Thomson, A. W. & Knolle, P. A. Antigen-presenting cell function in the tolerogenic liver environment. Nat. Rev. Immunol. 10, 753–766 (2010).
Cullender, TylerC. et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 14, 571–581 (2013).
Macpherson, A. J. & Uhr, T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 303, 1662–1665 (2004).
Helgeland, L. et al. Microbial colonization induces oligoclonal expansions of intraepithelial CD8 T cells in the gut. Eur. J. Immunol. 34, 3389–3400 (2004).
Lamont, R. J., Koo, H. & Hajishengallis, G. The oral microbiota: dynamic communities and host interactions. Nat. Rev. Microbiol. 16, 745–759 (2018).
Marsh, P. D. Are dental diseases examples of ecological catastrophes? Microbiology 149, 279–294 (2003).
Duran-Pinedo, A. E. et al. Community-wide transcriptome of the oral microbiome in subjects with and without periodontitis. ISME J. 8, 1659–1672 (2014).
Maekawa, T. et al. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell Host Microbe 15, 768–778 (2014).
Hilty, M. et al. Disordered microbial communities in asthmatic airways. PLoS One 5, e8578 (2010).
Dickson, R. P., Erb-Downward, J. R., Martinez, F. J. & Huffnagle, G. B. The microbiome and the respiratory tract. Annu. Rev. Physiol. 78, 481–504 (2016).
Barcik, W., Boutin, R. C. T., Sokolowska, M. & Finlay, B. B. The role of lung and gut microbiota in the pathology of asthma. Immunity 52, 241–255 (2020).
Soroosh, P. et al. Lung-resident tissue macrophages generate Foxp3+ regulatory T cells and promote airway tolerance. J. Exp. Med. 210, 775–788 (2013).
Sellge, G. & Kufer, T. A. PRR-signaling pathways: learning from microbial tactics. Semin. Immunol. 27, 75–84 (2015).
Gollwitzer, E. S. et al. Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat. Med. 20, 642–647 (2014).
Lai, Y. et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 15, 1377–1382 (2009).
Lai, Y. et al. Activation of TLR2 by a small molecule produced by Staphylococcus epidermidis increases antimicrobial defense against bacterial skin infections. J. Investig. Dermatol. 130, 2211–2221 (2010).
Naik, S. et al. Compartmentalized control of skin immunity by resident commensals. Science 337, 1115–1119 (2012).
Aiyar, A. et al. Influence of the tryptophan-indole-IFNγ axis on human genital Chlamydia trachomatis infection: role of vaginal co-infections. Front. Cell. Infect. Microbiol. 4, 72 (2014).
Brotman, R. M. et al. Microbiome, sex hormones, and immune responses in the reproductive tract: challenges for vaccine development against sexually transmitted infections. Vaccine 32, 1543–1552 (2014).
Gilbert, N. M. et al. Gardnerella vaginalis and prevotella bivia trigger distinct and overlapping phenotypes in a mouse model of bacterial vaginosis. J. Infect. Dis. 220, 1099–1108 (2019).
Garcia, E. M., Kraskauskiene, V., Koblinski, J. E. & Jefferson, K. K. Interaction of Gardnerella vaginalis and vaginolysin with the apical versus basolateral face of a three-dimensional model of vaginal epithelium. Infect. Immun. 87, e00646–00618 (2019).
Doerflinger, S. Y., Throop, A. L. & Herbst-Kralovetz, M. M. Bacteria in the vaginal microbiome alter the innate immune response and barrier properties of the human vaginal epithelia in a species-specific manner. J. Infect. Dis. 209, 1989–1999 (2014).
Fichorova, R. N. et al. The villain team-up or how Trichomonas vaginalis and bacterial vaginosis alter innate immunity in concert. Sexually Transmitted Infect. 89, 460–466 (2013).
Lau, K. et al. Bridging the gap between gut microbial dysbiosis and cardiovascular diseases. Nutrients. 9, 859 (2017).
Sanchez-Rodriguez, E. et al. The gut microbiota and its implication in the development of atherosclerosis and related cardiovascular diseases. Nutrients. 12, 605 (2020).
DeStefano, F. et al. Dental disease and risk of coronary heart disease and mortality. BMJ 306, 688–691 (1993).
Schenkein, H. A., Papapanou, P. N., Genco, R. & Sanz, M. Mechanisms underlying the association between periodontitis and atherosclerotic disease. Periodontology 2000. 83, 90–106 (2020).
D’Aiuto, F. et al. Periodontal infections cause changes in traditional and novel cardiovascular risk factors: results from a randomized controlled clinical trial. Am. Heart J. 151, 977–984 (2006).
Ramírez, J. H. et al. Biomarkers of cardiovascular disease are increased in untreated chronic periodontitis: a case control study. Aust. Dent. J. 59, 29–36 (2014).
Roncal, C. et al. Trimethylamine-N-oxide (TMAO) predicts cardiovascular mortality in peripheral artery disease. Sci. Rep. 9, 15580 (2019).
Zeisel, S. H. & Warrier, M. Trimethylamine N-oxide, the microbiome, and heart and kidney disease. Annu. Rev. Nutr. 37, 157–181 (2017).
Liu, T., Zhang, L., Joo, D. & Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2, 17023 (2017).
Yoshida, N. et al. A possible beneficial effect of bacteroides on faecal lipopolysaccharide activity and cardiovascular diseases. Sci. Rep. 10, 13009 (2020).
Mathewson, N. D. et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 17, 505–513 (2016).
Bartolomaeus, H. et al. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 139, 1407–1421, (2019).
Kvakan, H. et al. Regulatory T cells ameliorate angiotensin II-induced cardiac damage. Circulation 119, 2904–2912 (2009).
Wang, L. et al. Sodium butyrate suppresses angiotensin II-induced hypertension by inhibition of renal (pro)renin receptor and intrarenal renin-angiotensin system. J. Hypertens. 35, 1899–1908 (2017).
Wu, Y., Xu, H., Tu, X. & Gao, Z. The role of short-chain fatty acids of gut microbiota origin in hypertension. 12, 730809 (2021).
Li, Q. et al. Gut microbiota: a novel regulator of cardiovascular disease and key factor in the therapeutic effects of flavonoids. Front. Pharmacol. 12, 651926 (2021).
Ferlay J. et al. Global Cancer Observatory: Cancer Today. Lyon: International Agency for Research on Cancer. (2020).
Golemis, E. A. et al. Molecular mechanisms of the preventable causes of cancer in the United States. Genes Dev. 32, 868–902 (2018).
Schistosomes, liver flukes and Helicobacter pylori. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Lyon, 7–14 June 1994. IARC Monogr. Eval. Carcinog. Risks Hum. 61, 1–241, (1994).
Farrell, J. J. et al. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer. Gut 61, 582–588 (2012).
Michaud, D. S. et al. Plasma antibodies to oral bacteria and risk of pancreatic cancer in a large European prospective cohort study. Gut 62, 1764–1770 (2013).
Fan, X. et al. Human oral microbiome and prospective risk for pancreatic cancer: a population-based nested case-control study. Gut 67, 120–127 (2018).
Peters, B. A. et al. Oral microbiome composition reflects prospective risk for esophageal cancers. Cancer Res. 77, 6777–6787 (2017).
Sami, A. et al. The role of the microbiome in oral squamous cell carcinoma with insight into the microbiome-treatment axis. Int. J. Mol. Sci. 21, 8061 (2020).
Cao, W. et al. Homoharringtonine induces apoptosis and inhibits STAT3 via IL-6/JAK1/STAT3 signal pathway in Gefitinib-resistant lung cancer cells. Sci. Rep. 5, 8477 (2015).
Kuboniwa, M. et al. P. gingivalis accelerates gingival epithelial cell progression through the cell cycle. Microbes Infect. 10, 122–128 (2008).
Donehower, L. A. Phosphatases reverse p53-mediated cell cycle checkpoints. Proc. Natl. Acad. Sci. USA. 111, 7172–7173, (2014).
Zhou, Y. et al. Noncanonical activation of β-catenin by Porphyromonas gingivalis. Infect. Immun. 83, 3195–3203 (2015).
Takayama, S. et al. Inhibition of TGF-beta1 suppresses motility and invasiveness of oral squamous cell carcinoma cell lines via modulation of integrins and down-regulation of matrix-metalloproteinases. Oncol. Rep. 21, 205–210 (2009).
Vyhnalova, T., Danek, Z., Gachova, D. & Linhartova, P. B. The role of the oral microbiota in the etiopathogenesis of oral squamous cell carcinoma. Microorganisms. 9, 1549 (2021).
Atarashi, K. et al. Ectopic colonization of oral bacteria in the intestine drives T(H)1 cell induction and inflammation. Science 358, 359–365 (2017).
Kostic, A. D. et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 14, 207–215 (2013).
Gomes, S. et al. Profiling of lung microbiota discloses differences in adenocarcinoma and squamous cell carcinoma. Sci. Rep. 9, 12838 (2019).
Peters, B. A. et al. The microbiome in lung cancer tissue and recurrence-free survival. Cancer Epidemiol. Biomark. Prev. 28, 731–740 (2019).
Jin, C. et al. Commensal microbiota promote lung cancer development via γδ T cells. Cell 176, 998–1013.e1016 (2019).
Tsay, J.-C. J. et al. Airway microbiota is associated with upregulation of the PI3K pathway in lung cancer. Am. J. Respir. Crit. Care Med. 198, 1188-1198, (2018).
Tsay, J. J. et al. Lower airway dysbiosis affects lung cancer progression. Cancer Discov. 11, 293–307 (2021).
Honda, K. & Littman, D. R. The microbiota in adaptive immune homeostasis and disease. Nature 535, 75–84 (2016).
Grivennikov, S. I. et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 491, 254–258 (2012).
Li, S. et al. Tumorigenic bacteria in colorectal cancer: mechanisms and treatments. Cancer Biol. Med. 19, 147–162 (2021).
Chen, J., Pitmon, E. & Wang, K. Microbiome, inflammation and colorectal cancer. Semin Immunol. 32, 43–53 (2017).
Dai, Z. et al. The role of microbiota in the development of colorectal cancer. Int. J. Cancer 145, 2032–2041 (2019).
Kostic, A. D. et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 22, 292–298 (2012).
de Groot, P. F. et al. Distinct fecal and oral microbiota composition in human type 1 diabetes, an observational study. PLOS One. 12, e0188475 (2017).
Mariño, E. et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 18, 552–562 (2017).
Leiva-Gea, I. et al. Gut microbiota differs in composition and functionality between children with type 1 diabetes and MODY2 and healthy control subjects: a case-control study. Diabetes Care. 41, 2385–2395 (2018).
Zhou, H. et al. Gut microbiota profile in patients with type 1 diabetes based on 16S rRNA gene sequencing: a systematic review. Dis. Mark. 2020, 3936247 (2020).
Vatanen, T. et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature 562, 589–594 (2018).
Suzuki, T. et al. Diabetogenic effects of lymphocyte transfusion on the NOD or NOD nude mouse. Karger (1985).
Acosta-Montaño, P. et al. Fatty acid and lipopolysaccharide effect on beta cells proteostasis and its impact on insulin secretion. Cells 8, 884 (2019).
Devaraj, S., Dasu, M. R., Park, S. H. & Jialal, I. Increased levels of ligands of Toll-like receptors 2 and 4 in type 1 diabetes. Diabetologia 52, 1665–1668 (2009).
Gülden, E. et al. Toll-like receptor 4 deficiency accelerates the development of insulin-deficient diabetes in non-obese diabetic mice. PLoS One 8, e75385 (2013).
Li, J., Wang, X., Zhang, F. & Yin, H. Toll-like receptors as therapeutic targets for autoimmune connective tissue diseases. Pharm. Ther. 138, 441–451 (2013).
Hou, K. et al. Reconstruction of intestinal microecology of Type 2 diabetes by fecal microbiota transplantation: why and how. Bosn. J. Basic Med. Sci. https://doi.org/10.17305/bjbms.2021.6323 (2021).
Larsen, N. et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLOS One. 5, e9085 (2010).
Almugadam, B. S. et al. Alterations of gut microbiota in type 2 diabetes individuals and the confounding effect of antidiabetic agents. J. Diabetes Res. 2020, 7253978 (2020).
Gomes, J. M. G., Costa, J. A. & Alfenas, R. C. G. Metabolic endotoxemia and diabetes mellitus: a systematic review. Metabolism 68, 133–144 (2017).
Shapiro, H., Kolodziejczyk, A. A., Halstuch, D. & Elinav, E. Bile acids in glucose metabolism in health and disease. J. Exp. Med. 215, 383–396 (2018).
Psichas, A. et al. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 39, 424–429 (2015).
Takahashi, D. et al. Microbiota-derived butyrate limits the autoimmune response by promoting the differentiation of follicular regulatory T cells. EBioMedicine 58, 102913 (2020).
He, J. et al. The oral microbiome diversity and its relation to human diseases. Folia Microbiol. 60, 69–80 (2015).
Xiao, E. et al. Diabetes enhances IL-17 expression and alters the oral microbiome to increase its pathogenicity. Cell Host Microbe 22, 120–128.e124 (2017).
Radzicka, S., Pietryga, M., Iciek, R. & Brązert, J. The role of visfatin in pathogenesis of gestational diabetes (GDM). Ginekol. Pol. 89, 518–521 (2018).
Gomez-Arango, L. F. et al. Connections between the gut microbiome and metabolic hormones in early pregnancy in overweight and obese women. Diabetes 65, 2214–2223 (2016).
Koren, O. et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 150, 470–480 (2012).
Cortez, R. V. et al. Microbiome and its relation to gestational diabetes. Endocrine 64, 254–264 (2019).
Crusell, M. K. W. et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 6, 89 (2018).
Ponzo, V. et al. The microbiota composition of the offspring of patients with gestational diabetes mellitus (GDM). PLoS One 14, e0226545 (2019).
Wang, J. et al. Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut 67, 1614–1625 (2018).
van Woerden, H. C. et al. Differences in fungi present in induced sputum samples from asthma patients and non-atopic controls: a community based case control study. BMC Infect. Dis. 13, 69 (2013).
Millares, L. et al. Bronchial microbiome of severe COPD patients colonised by Pseudomonas aeruginosa. Eur. J. Clin. Microbiol Infect. Dis. 33, 1101–1111 (2014).
Han, M. K. et al. Lung microbiome and disease progression in idiopathic pulmonary fibrosis: an analysis of the COMET study. Lancet Respir. Med. 2, 548–556 (2014).
Huang, Y. J. et al. Airway microbiota and bronchial hyperresponsiveness in patients with suboptimally controlled asthma. J. Allergy Clin. Immunol. 127, 372–381 (2011). e371–373.
Essilfie, A. T. et al. Combined Haemophilus influenzae respiratory infection and allergic airways disease drives chronic infection and features of neutrophilic asthma. Thorax 67, 588–599 (2012).
Alnahas, S. et al. IL-17 and TNF-α are key mediators of moraxella catarrhalis triggered exacerbation of allergic airway inflammation. Front. Immunol. 8, 1562 (2017).
Garcia-Nuñez, M. et al. Severity-related changes of bronchial microbiome in chronic obstructive pulmonary disease. J. Clin. Microbiol. 52, 4217–4223 (2014).
Sokolowska, M. et al. Microbiome and asthma. Asthma Res. Pract. 4, 1 (2018).
Roussos, A. et al. Increased prevalence of irritable bowel syndrome in patients with bronchial asthma. Respir. Med. 97, 75–79 (2003).
Rutten, E. P. A., Lenaerts, K., Buurman, W. A. & Wouters, E. F. M. Disturbed intestinal integrity in patients with COPD: effects of activities of daily living. Chest 145, 245–252 (2014).
Sprooten, R. T. M. et al. Increased small intestinal permeability during severe acute exacerbations of COPD. Respiration 95, 334–342 (2018).
Ottiger, M. et al. Gut, microbiota-dependent trimethylamine-N-oxide is associated with long-term all-cause mortality in patients with exacerbated chronic obstructive pulmonary disease. Nutrition 45, 135–141.e131 (2018).
Arrieta, M. C. et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 7, 307ra152 (2015).
Roduit, C. et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 74, 799–809 (2019).
Arpaia, N. et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 504, 451–455 (2013).
Cait, A. et al. Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal. Immunol. 11, 785–795 (2018).
Tulic, M. K., Piche, T. & Verhasselt, V. Lung-gut cross-talk: evidence, mechanisms and implications for the mucosal inflammatory diseases. Clin. Exp. Allergy 46, 519–528 (2016).
Huang, Y. et al. S1P-dependent interorgan trafficking of group 2 innate lymphoid cells supports host defense. Science 359, 114–119 (2018).
Perrone, E. E. et al. Mechanisms of methicillin-resistant Staphylococcus aureus pneumonia-induced intestinal epithelial apoptosis. Shock 38, 68–75 (2012).
Coopersmith, C. M. et al. Sepsis from Pseudomonas aeruginosa pneumonia decreases intestinal proliferation and induces gut epithelial cell cycle arrest. Crit. Care Med. 31, 1630–1637 (2003).
Gleeson, K., Eggli, D. F. & Maxwell, S. L. Quantitative aspiration during sleep in normal subjects. Chest 111, 1266–1272 (1997).
Bassis, C. M. et al. Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. mBio 6, e00037 (2015).
Segal, L. N. et al. Enrichment of lung microbiome with supraglottic taxa is associated with increased pulmonary inflammation. Microbiome 1, 19 (2013).
Durack, J. et al. Features of the bronchial bacterial microbiome associated with atopy, asthma, and responsiveness to inhaled corticosteroid treatment. J. Allergy Clin. Immunol. 140, 63–75 (2017).
Aaron, S. D. et al. Granulocyte inflammatory markers and airway infection during acute exacerbation of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 163, 349–355 (2001).
Thomas, P. S. Tumour necrosis factor-alpha: the role of this multifunctional cytokine in asthma. Immunol. Cell Biol. 79, 132–140 (2001).
Jousilahti, P. et al. The association of sensitive systemic inflammation markers with bronchial asthma. Ann. Allergy Asthma Immunol. 89, 381–385 (2002).
de Steenhuijsen Piters, W. A. et al. Dysbiosis of upper respiratory tract microbiota in elderly pneumonia patients. ISME J. 10, 97–108 (2016).
Vissing, N. H., Chawes, B. L. & Bisgaard, H. Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 188, 1246–1252 (2013).
Selwyn, P. A. et al. Increased risk of bacterial pneumonia in HIV-infected intravenous drug users without AIDS. AIDS 2, 267–272 (1988).
Segal, L. N. et al. Anaerobic bacterial fermentation products increase tuberculosis risk in antiretroviral-drug-treated HIV patients. Cell Host Microbe 21, 530–537.e534 (2017).
Gomez, J. C. et al. Mechanisms of interferon-γ production by neutrophils and its function during Streptococcus pneumoniae pneumonia. Am. J. Respir. Cell Mol. Biol. 52, 349–364 (2015).
Tsai, H. C., Velichko, S., Hung, L. Y. & Wu, R. IL-17A and Th17 cells in lung inflammation: an update on the role of Th17 cell differentiation and IL-17R signaling in host defense against infection. Clin. Dev. Immunol. 2013, 267971 (2013).
Salk, H. M. et al. Taxa of the nasal microbiome are associated with influenza-specific IgA response to live attenuated influenza vaccine. PLoS One 11, e0162803 (2016).
Fan, J. et al. The lung tissue microbiota features of 20 deceased patients with COVID-19. J. Infect. 81, e64–e67 (2020).
Han Y, Jia Z, Shi J, Wang W, He K. The active lung microbiota landscape of COVID-19 patients through the metatranscriptome data analysis. Bioimpacts 12,139–146 (2022).
Sulaiman, I. et al. Microbial signatures in the lower airways of mechanically ventilated COVID-19 patients associated with poor clinical outcome. Nat. Microbiol. 6, 1245–1258 (2021).
Schuijt, T. J. et al. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut 65, 575 (2016).
Yeoh, Y. K. et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 70, 698 (2021).
Kochar, B., Orkaby, A. R., Ananthakrishnan, A. N. & Ritchie, C. S. Frailty in inflammatory bowel diseases: an emerging concept. Ther. Adv. Gastroenterol. 14, 17562848211025474 (2021).
Lucas López, R., Grande Burgos, M. J., Gálvez, A. & Pérez Pulido, R. The human gastrointestinal tract and oral microbiota in inflammatory bowel disease: a state of the science review. APMIS 125, 3–10 (2017).
Leone, V., Chang, E. B. & Devkota, S. Diet, microbes, and host genetics: the perfect storm in inflammatory bowel diseases. J. Gastroenterol. 48, 315–321 (2013).
McGuckin, M. A. et al. Intestinal barrier dysfunction in inflammatory bowel diseases. Inflamm. Bowel Dis. 15, 100–113 (2009).
Kaistha, A. & Levine, J. Inflammatory bowel disease: the classic gastrointestinal autoimmune disease. Curr. Probl. Pediatr. Adolesc. Health Care. 44, 328–334 (2014).
Marcuzzi, A. et al. Genetic and functional profiling of Crohn’s disease: autophagy mechanism and susceptibility to infectious diseases. Biomed. Res. Int. 2013, 297501 (2013).
Kleessen, B., Kroesen, A. J., Buhr, H. J. & Blaut, M. Mucosal and invading bacteria in patients with inflammatory bowel disease compared with controls. Scand. J. Gastroenterol. 37, 1034–1041 (2002).
Ahmed, I. et al. Microbiome, metabolome and inflammatory bowel disease. Microorganisms. 4, 20 (2016).
Manichanh, C., Borruel, N., Casellas, F. & Guarner, F. The gut microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 9, 599–608 (2012).
Morgan, X. C. et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 13, R79 (2012).
Wang, Z. K. et al. Intestinal microbiota pathogenesis and fecal microbiota transplantation for inflammatory bowel disease. World J. Gastroenterol. 20, 14805–14820 (2014).
Yu, Q. et al. Enterohepatic helicobacter species as a potential causative factor in inflammatory bowel disease: a meta-analysis. Medicines94, e1773 (2015).
Ni, J., Wu, G. D., Albenberg, L. & Tomov, V. T. Gut microbiota and IBD: causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 14, 573–584 (2017).
Kitamoto, S. et al. The intermucosal connection between the mouth and gut in commensal pathobiont-driven colitis. Cell 182, 447–462.e414 (2020).
Derrien, M. et al. Mucin-bacterial interactions in the human oral cavity and digestive tract. Gut Microbes 1, 254–268 (2010).
She, Y. Y. et al. Periodontitis and inflammatory bowel disease: a meta-analysis. BMC Oral. Health 20, 67 (2020).
Said, H. S. et al. Dysbiosis of salivary microbiota in inflammatory bowel disease and its association with oral immunological biomarkers. DNA Res. 21, 15–25 (2014).
Vavricka, S. R. et al. Periodontitis and gingivitis in inflammatory bowel disease: a case-control study. Inflamm. Bowel Dis. 19, 2768–2777 (2013).
Walker, E. R., McGee, R. E. & Druss, B. G. Mortality in mental disorders and global disease burden implications: a systematic review and meta-analysis. JAMA Psychiatry 72, 334–341 (2015).
Varghese, F. P. & Brown, E. S. The hypothalamic-pituitary-adrenal axis in major depressive disorder: a brief primer for primary care physicians. Prim. Care Companion J. Clin. Psychiatry 3, 151 (2001).
Hao, W. Z., Li, X. J., Zhang, P. W. & Chen, J. X. A review of antibiotics, depression, and the gut microbiome. Psychiatry Res. 284, 112691 (2020).
Sherwin, E., Dinan, T. G. & Cryan, J. F. Recent developments in understanding the role of the gut microbiota in brain health and disease. Ann. N.Y. Acad. Sci. 1420, 5–25 (2018).
Desbonnet, L. et al. Microbiota is essential for social development in the mouse. Mol. Psychiatry 19, 146–148 (2014).
de Punder, K. & Pruimboom, L. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front. Immunol. 6, 223 (2015).
Peirce, J. M. & Alviña, K. The role of inflammation and the gut microbiome in depression and anxiety. J. Neurosci. Res. 97, 1223–1241 (2019).
Jiang, H. et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain, Behav. Immun. 48, 186–194 (2015).
Winter, G., Hart, R. A., Charlesworth, R. P. & Sharpley, C. F. Gut microbiome and depression: what we know and what we need to know. Rev. Neurosci. 29, 629–643 (2018).
Luo, Y. et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl. Psychiatry 8, 1–10 (2018).
Heijtz, R. D. et al. Normal gut microbiota modulates brain development and behavior. Proc. Natl Acad. Sci. 108, 3047–3052 (2011).
Crumeyrolle-Arias, M. et al. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology 42, 207–217 (2014).
Hsiao, ElaineY. et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 155, 1451–1463 (2013).
Finegold, S. M. et al. Gastrointestinal microflora studies in late-onset autism. Clin. Infect. Dis. 35, S6–S16 (2002).
Adams, J. B. et al. Gastrointestinal flora and gastrointestinal status in children with autism–comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 11, 22 (2011).
Kang, D.-W. et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: an open-label study. Microbiome 5, 10 (2017).
Sherwin, E., Rea, K., Dinan, T. G. & Cryan, J. F. A gut (microbiome) feeling about the brain. Curr. Opin. Gastroenterol. 32, 96–102 (2016).
Minter, M. R. et al. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer’s disease. Sci. Rep. 6, 30028 (2016).
Juárez, I., Gratton, A. & Flores, G. Ontogeny of altered dendritic morphology in the rat prefrontal cortex, hippocampus, and nucleus accumbens following Cesarean delivery and birth anoxia. J. Comp. Neurol. 507, 1734–1747 (2008).
Li, W. et al. Structural changes of gut microbiota in Parkinson’s disease and its correlation with clinical features. Sci. China Life Sci. 60, 1223–1233 (2017).
Li, C. et al. Gut microbiota differs between parkinson’s disease patients and healthy controls in northeast China. Front. Mol. Neurosci. 12, 171 (2019).
Desai, M. S. et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 167, 1339–1353.e1321 (2016).
Scheperjans, F. et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov. Disord. 30, 350–358 (2015).
Vogt, N. M. et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 7, 13537 (2017).
Cattaneo, A. et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 49, 60–68 (2017).
Hill, J. M. et al. Pathogenic microbes, the microbiome, and Alzheimer’s disease (AD). Front. Aging Neurosci. 6, 127 (2014).
Singh, V. et al. Microbiota dysbiosis controls the neuroinflammatory response after stroke. J. Neurosci. 36, 7428–7440 (2016).
Houlden, A. et al. Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav. Immun. 57, 10–20 (2016).
Caso, J. R. et al. Colonic bacterial translocation as a possible factor in stress-worsening experimental stroke outcome. Am. J. Physiol. Regul. Integr. Comp. Physiol. 296, R979–R985 (2009).
Lin, D. et al. Association between the oral microbiome and brain resting state connectivity in smokers. Neuroimage 200, 121–131 (2019).
Hicks, S. D. et al. Oral microbiome activity in children with autism spectrum disorder. Autism Res. 11, 1286–1299 (2018).
Wingfield, B. et al. Variations in the oral microbiome are associated with depression in young adults. Sci. Rep. 11, 15009 (2021).
Yussof, A. et al. A meta-analysis of the effect of binge drinking on the oral microbiome and its relation to Alzheimer’s disease. Sci. Rep. 10, 19872 (2020).
Mihaila, D. et al. The oral microbiome of early stage Parkinson’s disease and its relationship with functional measures of motor and non-motor function. PLoS One 14, e0218252 (2019).
Yang, I. et al. The oral microbiome and inflammation in mild cognitive impairment. Exp. Gerontol. 147, 111273 (2021).
Liu, X. X. et al. Analysis of salivary microbiome in patients with Alzheimer’s disease. J. Alzheimers Dis. 72, 633–640 (2019).
GBD Chronic Kidney Disease Collaboration. Global, regional, and national burden of chronic kidney disease, 1990-2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 395, 709–733, (2020).
Pluznick, J. L. The gut microbiota in kidney disease. Science 369, 1426–1427 (2020).
Barros, A. F. et al. Is there interaction between gut microbial profile and cardiovascular risk in chronic kidney disease patients? Future Microbiol. 10, 517–526 (2015).
Li, L. et al. Periodontitis exacerbates and promotes the progression of chronic kidney disease through oral flora, cytokines, and oxidative stress. Front. Microbiol. 12, 656372 (2021).
Kshirsagar, A. V. et al. Antibodies to periodontal organisms are associated with decreased kidney function. The dental atherosclerosis risk in communities study. Blood Purif. 25, 125–132 (2007).
Bastos, J. A. et al. Identification of periodontal pathogens and severity of periodontitis in patients with and without chronic kidney disease. Arch. Oral. Biol. 56, 804–811 (2011).
Sharma, P. et al. Association between periodontitis and mortality in stages 3–5 chronic kidney disease: NHANES III and linked mortality study. J. Clin. Periodontol. 43, 104–113 (2016).
Yang, T., Richards, E. M., Pepine, C. J. & Raizada, M. K. The gut microbiota and the brain–gut–kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 14, 442–456 (2018).
Kanbay, M. et al. The crosstalk of gut microbiota and chronic kidney disease: role of inflammation, proteinuria, hypertension, and diabetes mellitus. Int. Urol. Nephrol. 50, 1453–1466 (2018).
Wang, I.-K. et al. Real-time PCR analysis of the intestinal microbiotas in peritoneal dialysis patients. Appl. Environ. Microbiol. 78, 1107–1112 (2012).
Mafra, D. & Fouque, D. Gut microbiota and inflammation in chronic kidney disease patients. Clin Kidney J. 8, 332–334 (2015).
Pan, W. & Kang, Y. Gut microbiota and chronic kidney disease: implications for novel mechanistic insights and therapeutic strategies. Int. Urol. Nephrol. 50, 289–299 (2018).
Vaziri, N. D. et al. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 83, 308–315 (2013).
Li, F. et al. Alterations to the gut microbiota and their correlation with inflammatory factors in chronic kidney disease. Front. Cell. Infect. Microbiol. 9, 206 (2019).
Lun, H. et al. Altered gut microbiota and microbial biomarkers associated with chronic kidney disease. Microbiologyopen 8, e00678 (2019).
Mafra, D. et al. Role of altered intestinal microbiota in systemic inflammation and cardiovascular disease in chronic kidney disease. Future Microbiol. 9, 399–410 (2014).
Lobel, L. et al. Diet posttranslationally modifies the mouse gut microbial proteome to modulate renal function. Science 369, 1518–1524 (2020).
Krishnamurthy, V. M. R. et al. High dietary fiber intake is associated with decreased inflammation and all-cause mortality in patients with chronic kidney disease. Kidney Int. 81, 300–306 (2012).
Chung, S., Barnes, J. L. & Astroth, K. S. Gastrointestinal microbiota in patients with chronic kidney disease: a systematic review. Adv. Nutr. 10, 888–901 (2019).
Andersen, K. et al. Intestinal dysbiosis, barrier dysfunction, and bacterial translocation account for CKD-related systemic inflammation. J. Am. Soc. Nephrol. 28, 76–83 (2017).
Poesen, R. et al. Metabolism, protein binding, and renal clearance of microbiota–derived p-cresol in patients with CKD. Clin. J. Am. Soc. Nephrol. 11, 1136–1144 (2016).
Castillo-Rodriguez, E. et al. Impact of altered intestinal microbiota on chronic kidney disease progression. Toxins 10, 300 (2018).
Mutsaers, H. A. M. et al. Proximal tubular efflux transporters involved in renal excretion of p-cresyl sulfate and p-cresyl glucuronide: implications for chronic kidney disease pathophysiology. Toxicol. Vitr. 29, 1868–1877 (2015).
Watanabe, H. et al. p-Cresyl sulfate causes renal tubular cell damage by inducing oxidative stress by activation of NADPH oxidase. Kidney Int. 83, 582–592 (2013).
Bolati, D. et al. Indoxyl sulfate induces epithelial-to-mesenchymal transition in rat kidneys and human proximal tubular cells. Am. J. Nephrol. 34, 318–323 (2011).
Sun, C.-Y., Hsu, H.-H. & Wu, M.-S. p-Cresol sulfate and indoxyl sulfate induce similar cellular inflammatory gene expressions in cultured proximal renal tubular cells. Nephrol. Dial. Transplant. 28, 70–78 (2013).
Tang, W. W. et al. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res. 116, 448–455 (2015).
Kim, R. B. et al. Advanced chronic kidney disease populations have elevated trimethylamine N-oxide levels associated with increased cardiovascular events. Kidney Int. 89, 1144–1152 (2016).
Osna, N. A., Donohue, T. M. Jr. & Kharbanda, K. K. Alcoholic liver disease: pathogenesis and current management. Alcohol Res. 38, 147–161 (2017).
Brůha, R. et al. Alcoholic liver disease. Prague Med. Rep. 110, 181–190 (2009).
Tripathi, A. et al. The gut–liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 15, 397–411 (2018).
Mouzaki, M. et al. Intestinal microbiota in patients with nonalcoholic fatty liver disease. Hepatology 58, 120–127 (2013).
Raman, M. et al. Fecal microbiome and volatile organic compound metabolome in obese humans with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 11, 868–875 (2013). e861-863.
Wong, V. W. et al. Molecular characterization of the fecal microbiota in patients with nonalcoholic steatohepatitis–a longitudinal study. PLoS One 8, e62885 (2013).
Chen, Y. et al. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 54, 562–572 (2011).
Payne, C. M. et al. Deoxycholate induces mitochondrial oxidative stress and activates NF-kappaB through multiple mechanisms in HCT-116 colon epithelial cells. Carcinogenesis 28, 215–222 (2007).
Morgan, M. J. & Liu, Z. G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 21, 103–115 (2011).
Corbin, K. D. & Zeisel, S. H. Choline metabolism provides novel insights into nonalcoholic fatty liver disease and its progression. Curr. Opin. Gastroenterol. 28, 159–165 (2012).
Spencer, M. D. et al. Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology 140, 976–986 (2011).
Schwabe, R. F. & Luedde, T. Apoptosis and necroptosis in the liver: a matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 15, 738–752 (2018).
Seki, E. et al. TLR4 enhances TGF-beta signaling and hepatic fibrosis. Nat. Med. 13, 1324–1332 (2007).
Arrese, M., Cabrera, D., Kalergis, A. M. & Feldstein, A. E. Innate Immunity and Inflammation in NAFLD/NASH. Dig. Dis. Sci. 61, 1294–1303 (2016).
Han, P., Sun, D. & Yang, J. Interaction between periodontitis and liver diseases. Biomed. Rep. 5, 267–276 (2016).
Yoneda, M. et al. Involvement of a periodontal pathogen, Porphyromonas gingivalis on the pathogenesis of non-alcoholic fatty liver disease. BMC Gastroenterol. 12, 16 (2012).
Akinkugbe, A. A. et al. Periodontitis and non-alcoholic fatty liver disease, a population-based cohort investigation in the study of health in Pomerania. J. Clin. Periodontol. 44, 1077–1087 (2017).
Hajishengallis, G. et al. Complement involvement in periodontitis: molecular mechanisms and rational therapeutic approaches. Adv. Exp. Med. Biol. 865, 57–74 (2015).
Acharya, C., Sahingur, S. E. & Bajaj, J. S. Microbiota, cirrhosis, and the emerging oral-gut-liver axis. JCI Insight. 2, e94416 (2017).
Seki, E. & Schnabl, B. Role of innate immunity and the microbiota in liver fibrosis: crosstalk between the liver and gut. J. Physiol. 590, 447–458 (2012).
Silva Santos, P. S., Fernandes, K. S. & Gallottini, M. H. Assessment and management of oral health in liver transplant candidates. J. Appl Oral. Sci. 20, 241–245 (2012).
Ling, Z. et al. Decreased diversity of the oral microbiota of patients with hepatitis b virus-induced chronic liver disease: a pilot project. Sci. Rep. 5, 17098 (2015).
Quigley, E. M. M. & Gajula, P. Recent advances in modulating the microbiome. F1000Research. 9, 46 (2020).
Cunningham, M. et al. Applying probiotics and prebiotics in new delivery formats—is the clinical evidence transferable? Trends Food Sci. Technol. 112, 495–506 (2021).
Miller, L. E., Zimmermann, A. K. & Ouwehand, A. C. Contemporary meta-analysis of short-term probiotic consumption on gastrointestinal transit. World J. Gastroenterol. 22, 5122 (2016).
Anderson, A. D. G. Randomised clinical trial of synbiotic therapy in elective surgical patients. Gut 53, 241–245 (2004).
Doron, S. & Snydman, D. R. Risk and safety of probiotics. Clin. Infect. Dis. 60, S129–S134 (2015).
Adamberg, S. et al. Survival and synergistic growth of mixed cultures of bifidobacteria and lactobacilli combined with prebiotic oligosaccharides in a gastrointestinal tract simulator. Microb. Ecol. Health Dis. 25, 3402 (2014).
Forchielli, M. L. & Walker, W. A. The role of gut-associated lymphoid tissues and mucosal defence. Br. J. Nutr. 93, S41–S48 (2005).
Shoaf, K., Mulvey, G. L., Armstrong, G. D. & Hutkins, R. W. Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect. Immun. 74, 6920–6928 (2006).
Belcheva, A. et al. Gut microbial metabolism drives transformation of MSH2-deficient colon epithelial cells. Cell 158, 288–299 (2014).
Donohoe, D. R. et al. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol. Cell. 48, 612–626 (2012).
Sasaki, M. et al. Transglucosidase improves the gut microbiota profile of type 2 diabetes mellitus patients: a randomized double-blind, placebo-controlled study. BMC Gastroenterol. 13, 81 (2013).
Bonfili, L. et al. Microbiota modulation as preventative and therapeutic approach in Alzheimer’s disease. FEBS J. 288, 2836–2855 (2021).
Kahouli, I. et al. Design and validation of an orally administrated active L. fermentum-L. acidophilus probiotic formulation using colorectal cancer Apc Min/+ mouse model. Appl. Microbiol. Biotechnol. 101, 1999–2019 (2017).
Hu, J. et al. Anti-tumour immune effect of oral administration of Lactobacillus plantarum to CT26 tumour-bearing mice. J. Biosci. 40, 269–279 (2015).
Gallo, A. et al. Modulation of microbiota as treatment for intestinal inflammatory disorders: an uptodate. World J. Gastroenterol. 22, 7186 (2016).
Lam, V. et al. Intestinal microbiota determine severity of myocardial infarction in rats. FASEB J. 26, 1727–1735 (2012).
Gan, X. T. et al. Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ. Heart Fail. 7, 491–499 (2014).
Zhou, A. et al. Gut microbiota: the emerging link to lung homeostasis and disease. J. Bacteriol. 203, e00454–20 (2021).
Chen, J. & Vitetta, L. Modulation of gut microbiota for the prevention and treatment of COVID-19. J. Clin. Med. 10, 2903 (2021).
Bafeta, A., Koh, M., Riveros, C. & Ravaud, P. Harms reporting in randomized controlled trials of interventions aimed at modifying microbiota: a systematic review. Ann. Intern. Med. 169, 240 (2018).
Carvalho, B. M. et al. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 55, 2823–2834 (2012).
Uribe-Herranz, M. et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J. Clin. Investig. 130, 466–479 (2019).
Bullman, S. et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 358, 1443–1448 (2017).
Sethi, V. et al. Gut microbiota promotes tumor growth in mice by modulating immune response. Gastroenterology 155, 33–37.e36 (2018).
Choi, I. J. et al. Helicobacter pylori therapy for the prevention of metachronous gastric cancer. N. Engl. J. Med. 378, 1085–1095 (2018).
Vetizou, M. et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 350, 1079–1084 (2015).
Hernández, E. et al. Functional consequences of microbial shifts in the human gastrointestinal tract linked to antibiotic treatment and obesity. Gut Microbes 4, 306–315 (2013).
Kilkkinen, A. et al. Antibiotic use predicts an increased risk of cancer. Int. J. Cancer 123, 2152–2155 (2008).
Dik, V. K., van Oijen, M. G. H., Smeets, H. M. & Siersema, P. D. Frequent use of antibiotics is associated with colorectal cancer risk: results of a nested case–control study. Dig. Dis. Sci. 61, 255–264 (2016).
Kyriacou, D. N. & Lewis, R. J. Confounding by indication in clinical research. JAMA 316, 1818 (2016).
Stripling, J. & Rodriguez, M. Current evidence in delivery and therapeutic uses of fecal microbiota transplantation in human diseases-clostridium difficile disease and beyond. Am. J. Med Sci. 356, 424–432 (2018).
Kassam, Z., Lee, C. H., Yuan, Y. & Hunt, R. H. Fecal microbiota transplantation for clostridium difficile infection: systematic review and meta-analysis. Am. J. Gastroenterol. 108, 500–508 (2013).
Ianiro, G. et al. Efficacy of different faecal microbiota transplantation protocols for Clostridium difficile infection: a systematic review and meta-analysis. Urol. Eur. Gastroenterol. J. 6, 1232–1244 (2018).
Khan, M. Y., Dirweesh, A., Khurshid, T. & Siddiqui, W. J. Comparing fecal microbiota transplantation to standard-of-care treatment for recurrent Clostridium difficile infection: a systematic review and meta-analysis. Eur. J. Gastroenterol. Hepatol. 30, 1309–1317 (2018).
van Nood, E. et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl. J. Med. 368, 407–415 (2013).
Routy, B. et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359, 91–97 (2018).
Zhang et al. Impact of fecal microbiota transplantation on obesity and metabolic syndrome—a systematic review. Nutrients 11, 2291 (2019).
Makkawi, S., Camara-Lemarroy, C. & Metz, L. Fecal microbiota transplantation associated with 10 years of stability in a patient with SPMS. Neurol. - Neuroimmunol. Neuroinflammation. 5, e459 (2018).
Peng, J., Xiao, X., Hu, M. & Zhang, X. Interaction between gut microbiome and cardiovascular disease. Life Sci. 214, 153–157 (2018).
Qi, X. et al. Treating steroid refractory intestinal acute graft-vs.-host disease with fecal microbiota transplantation: a pilot study. Front. Immunol. 9, 2195 (2018).
Minato, T. et al. Progression of Parkinson’s disease is associated with gut dysbiosis: two-year follow-up study. PLOS One. 12, e0187307 (2017).
Zhou, D. et al. Total fecal microbiota transplantation alleviates high-fat diet-induced steatohepatitis in mice via beneficial regulation of gut microbiota. Sci. Rep. 7, 1529 (2017).
Wortelboer, K., Nieuwdorp, M. & Herrema, H. Fecal microbiota transplantation beyond Clostridioides difficile infections. EBioMedicine 44, 716–729 (2019).
Ademe, M. Benefits of fecal microbiota transplantation: a comprehensive review. J. Infect. Dev. Ctries. 14, 1074–1080 (2020).
Dailey, F. E., Turse, E. P., Daglilar, E. & Tahan, V. The dirty aspects of fecal microbiota transplantation: a review of its adverse effects and complications. Curr. Opin. Pharmacol. 49, 29–33 (2019).
Benech, N. & Sokol, H. Fecal microbiota transplantation in gastrointestinal disorders: time for precision medicine. Genome Med. 12, 58 (2020).
Rosberg-Cody, E. et al. Recombinant lactobacilli expressing linoleic acid isomerase can modulate the fatty acid composition of host adipose tissue in mice. Microbiology 157, 609–615 (2011).
Chen, H.-L. et al. Probiotic Lactobacillus casei expressing human lactoferrin elevates antibacterial activity in the gastrointestinal tract. BioMetals 23, 543–554 (2010).
Koo, O. K., Amalaradjou, M. A. R. & Bhunia, A. K. Recombinant probiotic expressing listeria adhesion protein attenuates listeria monocytogenes virulence in vitro. PLoS One. 7, e29277 (2012).
Lagenaur, L. A. et al. Prevention of vaginal SHIV transmission in macaques by a live recombinant Lactobacillus. Mucosal. Immunol. 4, 648–657 (2011).
Caluwaerts, S. et al. AG013, a mouth rinse formulation of Lactococcus lactis secreting human Trefoil Factor 1, provides a safe and efficacious therapeutic tool for treating oral mucositis. Oral. Oncol. 46, 564–570 (2010).
Takiishi, T. et al. Reversal of autoimmune diabetes by restoration of antigen-specific tolerance using genetically modified Lactococcus lactis in mice. J. Clin. Investig. 122, 1717–1725 (2012).
Braat, H. et al. A Phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin. Gastroenterol. Hepatol. 4, 754–759 (2006).
Suzuki, A., Nakauchi, H. & Taniguchi, H. Glucagon-like peptide 1 (1-37) converts intestinal epithelial cells into insulin-producing cells. Proc. Natl Acad. Sci. 100, 5034–5039 (2003).
Duan, F. F., Liu, J. H. & March, J. C. Engineered commensal bacteria reprogram intestinal cells into glucose-responsive insulin-secreting cells for the treatment of diabetes. Diabetes 64, 1794–1803 (2015).
Forbes, N. S. Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 10, 785–794 (2010).
Toso, J. F. et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 20, 142–152 (2002).
Brown, J. M. & Wilson, W. R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 4, 437–447 (2004).
Xiang, S., Fruehauf, J. & Li, C. J. Short hairpin RNA-expressing bacteria elicit RNA interference in mammals. Nat. Biotechnol. 24, 697–702 (2006).
Hwang, I. Y. et al. Reprogramming microbes to be pathogen-seeking killers. ACS Synth. Biol. 3, 228–237 (2014).
Penninger, J. M., Grant, M. B. & Sung, J. J. Y. The role of angiotensin converting enzyme 2 in modulating gut microbiota, intestinal inflammation, and coronavirus infection. Gastroenterology 160, 39–46 (2021).
Verma, A. et al. Expression of human ACE2 in lactobacillus and beneficial effects in diabetic retinopathy in mice. Mol. Ther. Methods Clin. Dev. 17, 400 (2020).
Haga, S. B. in Principles of Gender-Specific Medicine (Third Edition) (ed Marianne J. Legato) 717–732 (Academic Press, 2017).
Sousa, T. et al. The gastrointestinal microbiota as a site for the biotransformation of drugs. Int. J. Pharm. 363, 1–25 (2008).
El Aidy, S., van den Bogert, B. & Kleerebezem, M. The small intestine microbiota, nutritional modulation and relevance for health. Curr. Opin. Biotechnol. 32, 14–20 (2015).
Sun, Y. et al. Effects of intestinal flora on the pharmacokinetics and pharmacodynamics of aspirin in high-altitude hypoxia. PLOS One. 15, e0230197 (2020).
Matuskova, Z. et al. Administration of a probiotic can change drug pharmacokinetics: effect of E. coli Nissle 1917 on amidarone absorption in rats. PLoS One 9, e87150 (2014).
Roberts, A. B. et al. Molecular insights into microbial β-glucuronidase inhibition to abrogate CPT-11 toxicity. Mol. Pharmacol. 84, 208–217 (2013).
Saha, J. R., Butler, V. P. Jr., Neu, H. C. & Lindenbaum, J. Digoxin-inactivating bacteria: identification in human gut flora. Science 220, 325–327 (1983).
Wu, B. et al. In vivo pharmacodynamic and pharmacokinetic effects of metformin mediated by the gut microbiota in rats. Life Sci. 226, 185–192 (2019).
Crouwel, F., Buiter, H. J. C. & de Boer, N. K. Gut microbiota-driven drug metabolism in inflammatory bowel disease. J. Crohn’s Colitis 15, 307–315 (2020).
Barnes, K. J., Rowland, A., Polasek, T. M. & Miners, J. O. Inhibition of human drug-metabolising cytochrome P450 and UDP-glucuronosyltransferase enzyme activities in vitro by uremic toxins. Eur. J. Clin. Pharmacol. 70, 1097–1106 (2014).
Klünemann, M. et al. Bioaccumulation of therapeutic drugs by human gut bacteria. Nature 597, 533–538 (2021).
Al-Asmakh, M. & Zadjali, F. Use of germ-free animal models in microbiota-related research. J. Microbiol. Biotechnol. 25, 1583–1588 (2015).
Yi, P. & Li, L. The germfree murine animal: an important animal model for research on the relationship between gut microbiota and the host. Vet. Microbiol. 157, 1–7 (2012).
Kennedy, E. A., King, K. Y. & Baldridge, M. T. Mouse microbiota models: comparing germ-free mice and antibiotics treatment as tools for modifying gut bacteria. Front Physiol. 9, 1534 (2018).
Peloquin, J. M. & Nguyen, D. D. The microbiota and inflammatory bowel disease: insights from animal models. Anaerobe 24, 102–106 (2013).
Nguyen, T. L., Vieira-Silva, S., Liston, A. & Raes, J. How informative is the mouse for human gut microbiota research? Dis. Model Mech. 8, 1–16 (2015).
Schumann, S. et al. Dextran sodium sulfate-induced inflammation alters the expression of proteins by intestinal Escherichia coli strains in a gnotobiotic mouse model. Appl. Environ. Microbiol. 78, 1513–1522 (2012).
Acknowledgements
The authors would like to acknowledge Dr. Yangmin Chen (St. John’s University, New York) for editing the article.
Author information
Authors and Affiliations
Contributions
J.L. and Z.S.C. conceived of the paper. K.H., Z.X.W., X.Y.C., J.Q.W., J.K., C.X. wrote and edited the paper. D.Zhang., D.Zhu. generated the figures. K.H., Z.X.W., L.W. revised the paper. All authors have read and approved the final paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hou, K., Wu, ZX., Chen, XY. et al. Microbiota in health and diseases. Sig Transduct Target Ther 7, 135 (2022). https://doi.org/10.1038/s41392-022-00974-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41392-022-00974-4
This article is cited by
-
Restoring the epigenetic landscape of lung microbiome: potential therapeutic approach for chronic respiratory diseases
BMC Pulmonary Medicine (2024)
-
Integrative metagenomic analysis reveals distinct gut microbial signatures related to obesity
BMC Microbiology (2024)
-
High-throughput fabrication of antimicrobial phage microgels and example applications in food decontamination
Nature Protocols (2024)
-
Altered gut microbiota in Taiwanese A97S predominant transthyretin amyloidosis with polyneuropathy
Scientific Reports (2024)
-
Bifidobacterium longum 1714 improves sleep quality and aspects of well-being in healthy adults: a randomized, double-blind, placebo-controlled clinical trial
Scientific Reports (2024)
