
Abstract
During the last 20 years, a new field of research delineating the importance of the microbiota in health and diseases has emerged. Inappropriate host-microbiota interactions have been shown to trigger a wide range of chronic inflammatory diseases, and defining the exact mechanisms behind perturbations of such relationship, as well as ways by which these disturbances can lead to disease states, both remain to be fully elucidated. The mucosa-associated microbiota constitutes a recently studied microbial population closely linked with the promotion of chronic intestinal inflammation and associated disease states. This review will highlight seminal works that have brought into light the importance of the mucosa-associated microbiota in health and diseases, emphasizing the challenges and promises of expending the mucosal microbiology field of research.
Similar content being viewed by others


Introduction
The intestinal microbiota is a vast and complex community of microorganisms inhabiting the gastrointestinal tract, encompassing 1013 bacteria per intestine and about 100–500 different species per individual.1,2 Among its functions in numerous aspects of host physiology, the gut microbiota is essential to promote maturation of the intestinal immune system as well as digestion by providing extraction of calories and nutrients that would otherwise be excreted in feces. This beneficial equilibrium between the host and its microbiota, or symbiosis, can also turn detrimental and participate in the development and/or the worsening of chronic inflammatory diseases.3,4 Hence, efforts have been made toward characterization of gut microbiota composition and function in health and diseases. Numerous studies have reported microbiota alterations in both preclinical and clinical models of chronic inflammatory diseases. Although those alterations may, in part, be driven by the disease state, recent works have highlighted that an altered microbiota can also play a central role in driving the disease itself, with the observation that disease can be transferred to germ-free mice through microbiota transplantation.5
Altogether, these findings suggest that the intestinal microbiota holds the potential for innovative therapeutic approaches, where microbiota modulations through fecal microbiota transplantation, diet, prebiotic or probiotic can be used to treat and/or prevent diseases. However, except for recurring Clostridium difficile infections, none of these approaches have demonstrated efficiency to restore host-microbiota homeostasis in chronic inflammatory disease patients. First, a great resilience is observed regarding human microbiota composition, so that its modulation is difficult to achieve and often only transient.6 Second, high interindividual variations, driven by genetic and environmental factors, are observed in human microbiota composition, suggesting the need for microbiota-based patient stratifications.7 Third, more work is still required to identify select microbiota alterations that are playing a role in disease progression, instead of simply being associated with it. Finally, the vast majority of the research performed on the intestinal microbiota is focusing on stool samples, while accumulating data demonstrate the importance of mucosa-associated microbiota in health and diseases. While some reviews previously elegantly described the role of host/microbiota interactions, they highlighted that these interactions are far from being characterized at the mucosal interface.8,9,10,11 Hence, we will highlight here several works that demonstrate the importance of the mucosa-associated microbiota, and we will discuss the promises of expanding the mucosal microbiology field of research.
Intestinal microbiota composition—3 dimensions matter
Longitudinal axis
As previously summarized by Donaldson et al., the lower gastrointestinal tract (i.e., the small intestine, the cecum, and the colon) harbors different habitats populated by specific bacterial communities, in a way that several microbial communities can be identified within the same individual and along a longitudinal axis.12 Each of these intestinal compartments is characterized by specific physiological, chemical, nutritional and immune conditions, which altogether shape region-specific communities. For example, and following the discovery of Helicobacter pylori, a limited microbiota comprising few genera (Propionibacterium, Lactobacillus, Streptococcus and Staphylococcus) was found in the stomach, while this organ was previously regarded as sterile.13 The small intestine harbors Lactobacillaceae (Firmicutes) and Enterobacteriaceae (Proteobacteria) facultative anaerobe families able to tolerate the acidic environment, as well as high levels of oxygen and antimicrobial molecules (e.g., bile acids).14,15 Finally, the cecum and the colon are mainly intended for complex carbohydrate fermentation and contained a denser and more diversified population, composed of the following dominant bacterial phyla: Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria, and Verrucomicrobia. This final part of the gastrointestinal tract is harboring what is considered the densest bacterial community on earth.
Transversal axis
The impact of the transversal axis on microbiota composition has been recently questioned. Work from several research teams revealed that several microenvironments exist and influence radial spatial distribution of bacterial communities.
From the lumen to the epithelium
It was for example demonstrated, in several mammal models (murine, swine, macaque or human), that the luminal and the mucosa-associated microbiota are significantly different in terms of composition (Table 1).16,17,18,19,20,21 From 1965 to 1972, a series of histological studies reported that bacteria embedded in the mucous layers of the epithelium had a distinct fusiform shape compared to those present in the lumen.16,17,18 Novel approaches, such as laser capture microdissection (LCM), further attempted to understand the impact of the transversal axis on microbiota composition. In the mouse ascending colon, Nava et al. notably demonstrated that the mucosa constitutes a specialized niche enriched in Firmicutes phylum (Lachnospiraceae, Ruminococcaceae families), while the luminal region is colonized by Bacteroidaceae, Enterococcaceae, and Lactobacillaceae families.22 In another interesting study, Liu et al. used the in vitro Twin Simulator of the Human Intestinal Microbial Ecosystem (TWIN-SHIME) model to compare luminal and mucosal communities tropism. With such an approach, unique mucosa-associated and lumen-associated microbial communities were observed in each of the colonic regions tested (ascending, transverse, and descending).23 While multiple studies demonstrated the existence of a transversal axis influencing microbiota composition, crypts- and inner mucus-associated microbiota have recently gained a lot of attention.
Crypt-associated microbiota
While the distribution of the gut microbiota along a radial axis was unraveled, pioneering work from Sansonetti’s team,24 combining Whartin—Starry (silver/nitrate) staining and FISH with 16S rRNA probes, established the existence of a “crypt-specific core microbiota” (CSCM).25 This CSCM was composed of limited bacterial members, which exclusively colonized the cecum and the proximal part of the colon, while being absent in the duodenum, jejunum and distal colon. Using microdissection and sequencing approaches, Firmicutes and Proteobacteria phyla were observed to be predominant in luminal crypts, while Bacteroidetes phylum was poorly represented.25 Interestingly, aerobic bacteria belonging to the Burkholderiales or Xanthomonadales groups dominated the CSCM, suggesting an oxygen-enrich environment close to the crypt. Follow-up studies demonstrated that Acinetobacter genera (Proteobacteria phylum, Moraxellaceae family) is more abundant in the crypts compared to the luminal environment, in both mice and humans.25,26 Besides Acinetobacter, the same group demonstrated that Delftia tsuruhatensis and Stenotrophomonas maltophilia are also important members of the CSCM.27 Following monocolonization of germ-free mice, these bacteria were detected in the colonic crypts, while the small intestinal crypt remained sterile.27 Highlighting their impact on intestinal homeostasis, the study of these CSCM members revealed their ability to decrease proliferation of epithelial cells, hence suggesting a central role for this community in intestinal homeostasis.27 Using an organoids approach, it was reported that, compared to a sonicated sample of Gram-positive bacteria grown in vitro, purified lipopolysaccharide (LPS) from CSCM-associated species led to organoid hypotrophy and stimulation of goblet cell differentiation that associated with IL-33 and Muc2 upregulation.27 Hence, this research elegantly demonstrated that the intestinal crypt environment harbors a unique microbial community of central importance for the mutualistic relationship between the host and its microbiota.
Inner mucus layer-associated microbiota
Besides crypt-associated microbiota, the role of the inner-mucus layer-associated microbiota in health and diseases has recently gained attention. While studies focusing on mucosa-associated microbiota are using crude extract from colonic biopsies, encompassing outer mucus-, inner mucus-, epithelium- and crypt-associated bacteria,17,19,22,23,25,26 more targeted approaches on microbiota members specifically colonizing the inner-mucus layer have demonstrated their association with detrimental outcomes in preclinical and clinical models. The importance of this mucus-associated microbial community in health and diseases will be described below (Chapter “Why is mucosa-associated microbiota important?”)
Time also matters
From birth to old age
Intestinal colonization by microorganisms starts at birth and undergoes significant changes during the first years of life.28,29 Fecal analyses have for example demonstrated that the Bacilli class is dominant after birth and subsequently decreases during the first 2 months, with concomitant increases in Clostridia and Gammaproteobacteria.30 Moreover, various perinatal parameters, encompassing health status of the mother, mode of delivery, antibiotic usage and type of feeding, are influencing bacterial colonization of the infant gut.31,32,33 Bergström et al. reported that the intestinal microbiota is subjected to significant changes between 9 and 18 months of age, with cessation of breastfeeding and introduction of solid food being essential factors impacting its composition.34 Interestingly, such microbiota shifts around the weaning period are associated with profound impacts on the host immune system maturation and function.35,36 Al Nabhani et al. elegantly demonstrated that the intestinal immune system undergoes a strong “weaning reaction” in young mice during which the intestinal microbiota is a key actor for the development of a balanced immune system.37,38 Importantly, such weaning reaction is central to protect against numerous inflammatory diseases later in life, in part through the induction of RORγt + regulatory T cells.37,38
In adults, a core microbiota of around 40 species that account for more than 75% of the community can be detected per subject over a 1-year period, suggesting that some microbiota members may be residents for decades.39,40 In the elderly population, fecal microbiota appears enriched in Enterobacteriaceae and depleted in Clostridium cluster IV and XIVa as well as Bifidobacterium.41 While some links have been established between alterations in the intestinal microbiota composition and type 2 diabetes (T2D), cancer (see below) and Alzheimer’s disease,42 the exact impact of microbiota evolution on senescence and age-associated diseases remains to be fully elucidated.43,44
Circadian oscillations
The intestinal microbiota is also subjected to circadian oscillations. In humans, Kaczmarek et al. demonstrated that the intestinal microbiota composition fluctuates during the day.45 Moreover, elegant mouse studies bring into light the close relationship between microbiota and the light/dark cycle, as well reviewed by Parkar et al.46 Importantly, perturbations of these circadian oscillations can trigger microbiota perturbations and intestinal barrier dysfunction.47,48 Moreover, circadian oscillations of serum metabolites are a microbiota-dependent process, and the intestinal microbiota has been observed as a central actor in modulating the circadian liver transcriptome and detoxification ability.49 Altogether, these data demonstrate that numerous spatial and temporal factors act in concert to finely regulate the intestinal microbiota composition and function.
Why is the mucosa-associated microbiota important?
Metabolic disorders
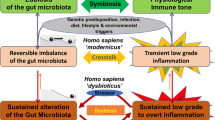
The potential role played by mucosa-associated microbiota in chronic inflammatory diseases was highlighted by multiple models of inflammation describing an “aggressive” microbiota able to penetrate the normally sterile inner mucus layer (Fig. 1). For example, consumption of synthetic dietary emulsifiers, carboxymethylcellulose (CMC) and polysorbate 80 (P80), altered mucosa-associated microbiota composition and function, leading to low-grade intestinal inflammation and metabolic disorders in mice50 (cf. details below). Importantly, it was also previously reported that microbiota encroachment is a feature of metabolic deregulations in humans, with the observation that microbiota-epithelium distance is inversely correlated with body mass index, fasting glucose levels, and hemoglobin A1C.51 Moreover, pioneering work by Cani et al. elegantly demonstrated that endotoxemia, referring to the elevation in circulating LPS, is associated with the promotion of metabolic disorders such as T2D and obesity.52 Such endotoxemia can occur following microbiota disturbance and/or increase in intestinal permeability, for example induced by a high-fat diet (HFD) regimen.53 Hence, these studies further illustrated the need for a well-controlled host/microbiota interaction at the mucosal surface.

At steady state (left part), the mucus layer keeps the bacterial community at a safe distance, while select symbionts favor maturation of the mucosal immune system by interacting with the host epithelium. In response to various stressors (right part), the mucus layer is altered in a way that leads to microbiota encroachment and chronic inflammatory diseases. Such altered host/microbiota relationship can be reversed by fiber-rich diet consumption, Akkermansia muciniphila administration or targeted mucosal immunization.
Inflammatory bowel diseases (IBD) and irritable bowel syndrome (IBS)
IBD include Crohn’s disease (CD) and ulcerative colitis (UC) that are chronic idiopathic disorders causing inflammation in the gastrointestinal tract.54 The etiology of these diseases has been linked to genetic factors and aberrant immune response to the gut microbiota.55,56,57,58 Decreased microbiota diversity was repeatedly found in fecal59,60 and mucosal61,62 samples from IBD patients, with depletion in anaerobic bacteria such as Bacteroides, Eubacterium or Lactobacillus.61 For example, lower abundance of Faecalibacterium prausnitzii (Firmicutes phylum) was observed in IBD patients60,62 in a way that associates with a higher risk of relapse.62,63 Furthermore, investigating microbiota organization along the transversal intestinal axis appears of central importance in IBD, with the presence of adherent and invasive microbiota members. Members of the Enterobacteriaceae family, such as the adherent invasive Escherichia coli (AIEC) pathovar,64,65 have an increased prevalence in the mucosa of IBD patients compared with healthy controls.66,67,68,69 AIEC pathobionts are flagellated and express multiple virulence factors with unique regulation of their expression, such as the Vat mucinase that allows mucins degradation.70 Altogether, these factors enhance AIEC’s ability to penetrate the mucus layer and to adhere to and invade intestinal epithelial cells in a way that promotes chronic intestinal inflammation.71 Ongoing studies suggest that fecal screening of AIEC bacteria fails to properly identify people carrying such bacteria in their intestinal mucosa, highlighting the importance to characterize this specific microbial population in IBD patients. Supporting this concept, a recent study demonstrated that CD relapse following partial bowel resection can be predicted by mucosa-associated microbiota composition in the ileum.72
IBS is a gastrointestinal disorder syndrome whose etiology remains unclear. The rectal mucosa-associated microbiota has been proposed as a potential predictor of small intestinal overgrowth, a common feature of IBS.73 Jabbar et al. recently analyzed mucus from sigmoid colon biopsies through metaproteomic approaches. They identified Brachyspira in 30–40% of IBS patients, suggesting a role for this bacterium in IBS, and further highlighting the importance of the mucosal microbiota in gastrointestinal disorders.74
Colorectal cancer
Colorectal cancer (CRC) is the third most prevalent cancer worldwide and is associated with a high lethality rate.75 Alterations of fecal and tumor-associated microbiota composition have been described.76,77 Some bacterial strains have been associated with susceptibility to CRC and have focused particular attention during the past decade, such as Fusobacterium nucleatum, enterotoxigenic Bacteroides fragilis (ETBF) and Escherichia coli expressing the polyketide synthase genomic island that enables colibactin expression.55,78,79,80,81,82 Efforts toward identification of CRC-associated microbiota have also highlighted the importance of mucosa-associated microbiota.26,81,82,83,84,85,86,87 For example, Saffarian et al. characterized crypt-associated and mucosal communities in CRC. They identified Bacteroidetes, Firmicutes and Proteobacteria in murine and human colonic crypts, as well as differential microbial signatures according to the colonic area. For example, Fusobacterium and Bacteroides fragilis were enriched in right-sided tumors, while Parvimonas micro abundance was elevated in left-sided tumors.26 However, the exact role played by such alterations in tumorigenesis remains to be fully characterized.
Are mucosa-associated bacteria always bad?
While, as highlighted above, mucosa-associated microbiota is involved in various disease states, it also appears as a critical factor influencing maturation of the intestinal immune system.
Segmented filamentous bacteria
The best example illustrating this concept is the work on Segmented Filamentous Bacteria (SFB), which is able to stimulate both innate and adaptive immune responses, notably Th17 (Fig. 1). This bacterium is adhering to the epithelium in the terminal region of the small intestine. It harbors a nipple-like appendage inserted into the epithelium, especially in the follicle-associated epithelium of Peyer’s patches,88 which forms an attachment site with pronounced actin rearrangements.89,90,91 While adherence to the epithelium is a hallmark of enteropathogens, SFB are autochthonous bacteria found to play an essential role for the host immune system. SFB colonization is observed after weaning and for 2 months before declining in mice, while in humans, its presence is detected in the first 2 years of life.92,93,94,95 Studies using SFB-monocolonized mice showed that SFB stimulate mouse intestinal and peripheral IgA responses with a potent activation of Peyer’s patches germinal centers, gut-associated lymphoid structures required for SFB-mediated intestinal IgA responses.96,97,98 Gaboriau-Routhiau et al. demonstrated that SFB monocolonisation is sufficient to recapitulate both innate and adaptive immune priming with strong Th1, Th2, Th17 and Treg responses compared to germ-free mice,99 and Ivanov et al. also demonstrated the central role played by SFB in Th17 responses. Associated with its strong immunostimulatory capacities, SFB has also been described to contribute to colonization resistance against several enteric pathogens such as Citrobacter rodentium and Salmonella enteritidis.100,101 More recently, Shi et al. reported that SFB colonization is sufficient to protect mice against rotavirus infection.102 Hence, while mucosa-associated bacteria can promote chronic inflammatory diseases, data on SFB demonstrate that this microbial community is also involved in central aspects of symbiosis. Modulation/inhibition of mucosa-associated bacteria should therefore be performed with extreme caution, and recent success in in vitro culture of SFB should bring important knowledge on such duality.103
Bacteroides fragilis
Works from Mazmanian et al. bring into light the central role of Bacteroides fragilis at the mucosal surface. After observing that this bacterium can bind to the mucus layer,104 they demonstrated that B. fragilis possesses a unique genetic locus of commensal colonization factors, referred to as the CCF system, enabling this bacterium to reside within the mucosal surface.105 It was also observed that, at steady state, the immune system tolerates B. fragilis despite its mucosal localization, with a mechanism that involves intestinal IgA.106 Moreover, B. fragilis is an obligate anaerobe but can nonetheless tolerate the oxidative stress caused by the epithelium through the alkyl hydroperoxide reductase, suggesting that this bacterium is well equipped for a mucosal niche colonization.107 Importantly, through the expression of its polysaccharide (PSA), B. fragilis can modulate T cell responses and cytokine production.108 Mechanistically, B. fragilis delivers its PSA to dendritic cells through outer membrane vesicles recognized by TLR2, which subsequently trigger immunomodulatory effects.109 In several models, including colitis, CRC and viral encephalitis, this bacterium was observed to be highly protective, further demonstrating the ability of select mucus-associated microbiota members to promote health.110,111,112
Regulation of the mucosa-associated microbiota
The oxygen hypothesis
The gut microbiota is influenced by several environmental factors, including oxygen concentration. As mentioned previously, the CSCM niche is mainly dominated by aerobic genera (e.g., Acinetobacter, strictly aerobic), suggesting an oxygen-enriched environment in the proximity of the epithelium.113 Combining an intraluminal probe with a phosphorescent quenching method, Albenberg et al. assessed oxygen distribution along the radial axis of the mouse intestine and demonstrated that it diffuses from epithelial cells to the lumen.114 In humans, the analysis of mucosal biopsies showed that adherent bacteria (Proteobacteria, Actinobacteria) were more aerotolerant than luminal bacteria and preferentially metabolize proteins instead of carbohydrates as substrates. Moreover, fecal microbiota composition is altered following hyperbaric oxygen therapy in mice, suggesting the impact of oxygen on the transversal microbiota compartmentalization.114
Several observations indicate that alteration in this radial oxygen distribution may lead to opportunistic pathogens invasion and disease development.113,115,116 Expansion of the aerobic zone is notably suspected of playing a role in inflammatory bowel diseases (IBDs). Indeed, IBD-associated dysbiosis is characterized by a decreased proportion of strict anaerobes (Faecalibacterium prausnitzii) in combination with the overgrowth of facultative aerobes, particularly the Enterobacteriaceae family (AIEC).60,61,62,66,67,69,117,118 Rigottier-Gois assimilated this dysbiosis to dysanaerobiosis, creating a favorable environment for the growth and invasion of aerotolerant bacteria that can enhance inflammation.113
The oxygen hypothesis was further consolidated by works from Baümler et al. exploring mechanisms lowering colonization resistance against Enterobacteriaceae, which demonstrated that antibiotic treatment promotes the shift from anaerobe to aerobe bacteria through various mechanisms.116 Streptomycin was reported to disrupt gut microbiota composition, depleting Clostridia class.116,119,120 Yet, Clostridia are important producers of short-chain fatty acids, such as butyrate, which is used as an energy source by mature colonocytes.121,122 Butyrate metabolization requires substantial quantities of oxygen, promoting a hypoxic environment close to the epithelium and avoiding aerobic bacteria colonization.123,124 In the absence of butyrate, colonocytes switch to glucose fermentation to obtain ATP. Hence, streptomycin, by reducing Clostridia, indirectly favors Enterobacteriaceae bacteria by generating an aerobic niche.116,120 In addition, streptomycin was reported to promote synthesis of the inducible nitric oxide synthase and subsequent production of nitric oxide, a reactive nitrogen species suggested to catalyze monosaccharide oxidation, leading to an increase in resources critical for pathogens (e.g., glucarate and galactarate that constitute a nutrient niche for species like E. coli and S. typhimurium).125 These studies elegantly indicate that controlling oxygen concentration at the epithelium surface may represent a possible mechanism to shape bacterial colonization of this specific intestinal niche. Interestingly, recent work from Bäumler’s team demonstrated that Citrobacter rodentium pathogen is able to intimately attach to the intestinal epithelium and grow through hydrogen peroxide (H2O2) respiration in the non-inflamed gut, with a central role played by the NAPDH oxidase NOX1. These data suggest that H2O2 can also be an important player in the regulation of the mucosa-associated microbiota.126
Dietary factors
Among factors modulating bacterial colonization at the mucosal side, diet is certainly one of the most important. As highlighted below, several dietary factors may alter mucosa-associated microbiota in a way that promotes chronic intestinal inflammation.
Western diet
Western Diet (WD) is characterized by an increased fat intake, refined sugars/sweeteners and animal proteins, and a decreased consumption of fruits, vegetables, and whole grains.127 WD consumption is strongly associated with metabolic disorders, such as obesity, T2D, and non-alcoholic liver disease. Over the last decades, the role of the host/microbiota relationship was demonstrated in several WD-induced pathologies (reviewed in128,129). It was for example shown that HFD shifts commensal bacterial composition, increasing Proteobacteria, while decreasing Bacteroidetes relative abundance.130 Interestingly, Schroeder et al. reported that WD affects the growth rate and penetrability of the colonic mucus layer compared to a control fiber-rich diet.131 Moreover, WD-fed mice displayed reduced inner mucus layer thickness and slower mucus growth.131 They also observed increased mucus penetrability of pathogens, as well as goblet cells hypertrophy and Muc2 and Dmbt1 overproduction, likely reflecting host compensatory mechanisms.131 Fructose is another nutrient part or the WD, and Montrose et al. recently demonstrated, in both Citrobacter rodentium-induced colitis and IL10−/− models, that a high fructose diet is sufficient to worsen intestinal inflammation and damage the integrity of the gut barrier. A reduction of the mucus layer thickness associated with bacterial colonization was observed, demonstrating that numerous dietary factors are acting in combination to regulate the host/microbiota interactions at the mucosal surface.132
Fiber-free diet
Studies indeed demonstrated that a fiber-depleted diet participates in chronic inflammatory diseases through modulation of microbiota composition and function.133,134 For example, mice fed a low-fat low-fiber diet exhibit increased body weight and altered metabolism compared to mice fed a fiber-rich diet.135 Other studies demonstrated that supplementation with oligofructose, a chicory inulin-type fructan, is sufficient to improve metabolic parameters altered by a HFD.136,137,138,139
Desai et al. elegantly demonstrated that fiber deprivation altered the intestinal microbiota in a way that promoted degradation of the colonic mucus barrier and enhanced pathogen expansion, exacerbating susceptibility to colitis.140 Using a synthetic minimal microbiota, they demonstrated that, in the absence of fiber, the gut microbiota shifted towards mucin degraders to fulfill its nutritional needs. Consequently, bacteria such as A. muciniphila or B. thetaiotaomicron, erode the colonic mucus barrier, hence enabling epithelium colonization by pathogenic bacteria, as demonstrated using the Citrobacter rodentium infection model. In 2016, Sonnenburg et al. demonstrated, in a mouse model, that dysbiosis and associated disorders induced by a fiber-deprived diet are reversible within a generation, but became non-reversible after multiple generations.141 Hence, a fiber-rich diet appears as an effective way to reinforce the intestinal barrier via its beneficial impact on mucosa-associated microbiota.
Emulsifiers
Select food additives appear to play an important role in regulating mucus-associated microbiota composition and function. It has been hypothesized that emulsifiers, which are added to most processed foods to improve texture and extend shelf life, might have contributed to the rapid post–mid-20th century increase in the incidence of chronic inflammatory diseases.142,143 Investigation of this hypothesis demonstrated that dietary emulsifiers can indeed detrimentally impact the intestinal microbiota in a way that drives chronic inflammatory diseases. In wild-type mice, bacteria were only rarely observed within 10 µm of the epithelium, and the average closest bacteria detected over multiple high-powered fields was about 25 µm from the epithelium.50 In contrast, in mice fed with dietary emulsifying agents CMC and P80, bacteria could be found in direct contact with the epithelium, and the average distance of the closest bacteria per field was <10 µm.50 Such effects of emulsifier exposure on the microbiota were associated with the development of chronic colitis in susceptible mice, while wild-type mice developed chronic low-grade intestinal inflammation and metabolic deregulations. Importantly, microbiota/epithelium distance inversely correlated with the extent of intestinal inflammation, supporting the central and direct role played by mucus penetrating bacteria in emulsifier-induced promotion of chronic intestinal inflammation.50 It was also reported that in gnotobiotic mice colonized with a highly restricted microbiota comprised of only eight bacteria (namely “Altered Schaedler Flora”, ASF), emulsifier consumption was not sufficient to induce microbiota encroachment, intestinal inflammation, nor metabolism alteration. This thus suggested that a complex microbiota containing specific species infiltrating the mucus layer is required for the detrimental effect of emulsifiers.144 In more recent work, pathobiont colonization of ASF mice was observed to be sufficient to make the animals susceptible to microbiota encroachment and chronic intestinal inflammation induced by emulsifier consumption.145 Hence, this demonstrates that select bacteria are needed to mediate the detrimental effect of emulsifier exposure through their encroachment within the mucus layer and subsequent promotion of chronic intestinal inflammation. Further research is now warranted to identify these mucus invaders in both animal and human models.
Mucosa-associated Microbiota—Opening of therapeutic avenues?
As detailed above, there is now evidence that mucosa-associated microbiota is crucial for host-bacteria interactions, which may open the door for therapeutic strategies in the coming years. On one hand, some commensals, such as Akkermansia muciniphila, may help to maintain mucosal integrity through a probiotic-type mechanism. On another hand, microbiota encroachment was reported to associate with an array of poor health outcomes, with the observation that a reduced epithelium-microbiota distance correlates with the severity of intestinal inflammation in mice and dysglycemia in human.50,51 While any causal link between microbiota encroachment and chronic inflammatory diseases remains to be studied, this underscores the need for novel approaches to target and inhibit encroachment of deleterious bacteria (Fig. 1).
Probiotic approach: the example of Akkermansia muciniphila
Isolated in 2004 by Derrien et al.,146 A. muciniphila is a mucin-degrading bacterium and one of the most described commensal bacterium residing in the intestinal mucus layer.147,148 Its abundance is positively associated with metabolic health, as elegantly demonstrated by the work of De Vos’ and Cani’s teams.149,150,151 Everard et al. for example reported that mucus thickness is reduced by half in diet-induced obese mice, a phenomenon that can be prevented by A. muciniphila treatment.149 It was also reported that this bacterium can increase the number of regulatory T cells and goblet cells, as well as the expression of the antimicrobial Reg3 γ peptide, in the intestine.152,153,154 In vitro, A. muciniphila adheres to the intestinal epithelium and strengthens enterocyte monolayer integrity.155 More recently, a single protein, called Amuc_1100, was sufficient to improve gut barrier and partly recapitulate the previously observed beneficial effects in vivo.156,157 Moreover, it was reported that treatment with A. muciniphila-derived extracellular vesicles could enhance intestinal tight junction function, reduce body weight gain and improve glucose homeostasis in obese mice.158 Plovier et al. demonstrated that pasteurization of this bacterium did not affect its beneficial effects, but even potentiated it.157,159 Finally, in a human pilot study, pasteurized A. muciniphila tended to decrease fat-mass gain and hip circumference, as well as improved insulin sensitivity, total cholesterol and blood markers of liver dysfunction, inflammation and endotoxemia in a cohort of overweight human volunteers.120 Altogether, these data elegantly support the therapeutic interest of this bacterium able to colonize the mucosa.149,160,161
Interleukine-33 (IL-33)
IL-33 belongs to the IL-1 cytokine family, binds the ST2 receptor, and is involved in various cellular pathways, including cytokine secretion, epithelial repair process and cell replication and survival.162,163,164,165 IL-33 appears as a crucial amplifier of the mucosal and systemic innate and acquired immune response.166,167 Interestingly, IL-33−/− mice displayed a dysbiosis and a decrease in Paneth and goblet cells that associated with an increased susceptibility to colitis.168,169,170 In a mouse model of chronic colitis, IL-33 administration led to disease improvement and increased mucin production.163,168,171,172,173,174 Several publications demonstrated that IL-33 plays a role in preventing encroachment by pathogens such as helminths,175,176,177,178 Clostridium difficile,174 Salmonella Typhimurium179 and H. pylori.180 Thus, IL-33 emerges as a central actor in regulating the host/microbiota interactions at the mucosal surface. However, caution appears warranted, as some studies reported a colitogenic impact of IL-33. In the DSS-induced colitis model, IL-33 administration was found to exacerbate intestinal inflammation, while IL-33KO mice were protected against colitis.181,182,183 Hence, while this cytokine could be a therapeutic tool to prevent microbiota encroachment, its exact role in modulating the host/microbiota relationship remains to be fully elucidated.
Mucosal immunization
It was recently hypothesized that the mucosal adaptive immune system, in close contact with the microbiota at the mucosal side, can be used to prevent microbiota encroachment by excluding motile bacteria from the mucus layer through targeted anti-flagellin response. This idea stemmed from a study revealing that flagellum appendage is central to the ability of bacteria to penetrate the colonic mucus layer,71 and from recent observations indicating that purified anti-flagellin antibody can rapidly shut down flagellin expression, and thereby bacterial motility.184 Based on these findings, it was speculated that flagellin immunization might result in lower levels of bioactive flagellin in a way that will prevent microbiota encroachment. Mice immunized with purified flagellin displayed strong fecal anti-flagellin IgG and IgA responses that associate with reduced expression of flagellin by the intestinal microbiota. Moreover, flagellin immunization was sufficient to increase the distance separating the microbiota from the epithelium in a way that correlated with protection against colitis and diet-induced obesity.184 Overall, these data support the concept that vaccination strategies aiming to prevent microbiota encroachment might protect against, or perhaps even treat, chronic inflammatory diseases with a microbiota component.
Conclusion
Multiple lines of evidence point to the importance of mucosa-associated microbiota in health and diseases. Additional research is warranted to characterize this “hidden” ecosystem, and uncover mechanisms by which it can promote health as well as inflammatory and metabolic disorders. To date, most microbiota studies have relied on analyzing microbiota composition via 16S rRNA gene sequencing of fecal material. While the mucosa-associated microbiota is of central importance, its study requires access to intestinal biopsies combined with molecular/culturomic approaches. Moreover, the causal link between microbiota encroachment and the promotion of chronic inflammatory diseases remains to be fully elucidated. Recently developed approaches allowing to specifically study mucus-associated microbiota should help in this endeavor.185,186 Besides its composition, identification of gene expression by this specific community also appears warranted in order to understand mechanisms by which select microbiota members colonize this unique niche in a way that induce chronic intestinal inflammation. Hence, while this mucosal microbiology field of research is just emerging, it holds exciting promises for the prevention and/or treatment of chronic inflammatory diseases!
References
Qin, J. et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464, 59–65 (2010).
Lloyd-Price, J., Abu-Ali, G. & Huttenhower, C. The healthy human microbiome. Genome Med. 8, 51 (2016).
Chassaing, B., Aitken, J. D., Gewirtz, A. T. & Vijay-Kumar, M. Chapter Three - Gut Microbiota Drives Metabolic Disease in Immunologically Altered Mice. in Advances in Immunology (Alt, F. W.) 116, 93–112 (Academic Press, 2012).
Belkaid, Y. & Hand, T. W. Role of the Microbiota in Immunity and Inflammation. Cell 157, 121–141 (2014).
Nagao-Kitamoto, H. et al. Functional Characterization of Inflammatory Bowel Disease–Associated Gut Dysbiosis in Gnotobiotic Mice. Cell Mol. Gastroenterol. Hepatol. 2, 468–481 (2016).
Palleja, A. et al. Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 3, 1255 (2018).
Rothschild, D. et al. Environment dominates over host genetics in shaping human gut microbiota. Nature https://doi.org/10.1038/nature25973 (2018).
Bunker, J. J. & Bendelac, A. IgA Responses to Microbiota. Immunity 49, 211–224 (2018).
Stange, E. F. & Schroeder, B. O. Microbiota and mucosal defense in IBD: an update. Expert Rev. Gastroenterol. Hepatol. 13, 963–976 (2019).
Etienne-Mesmin, L. et al. Experimental models to study intestinal microbes–mucus interactions in health and disease. FEMS Microbiol. Rev. 43, 457–489 (2019).
Ahern, P. P. & Maloy, K. J. Understanding immune–microbiota interactions in the intestine. Immunology 159, 4–14 (2020).
Donaldson, G. P., Lee, S. M. & Mazmanian, S. K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 14, 20–32 (2016).
Delgado, S., Cabrera-Rubio, R., Mira, A., Suárez, A. & Mayo, B. Microbiological Survey of the Human Gastric Ecosystem Using Culturing and Pyrosequencing Methods. Micro. Ecol. 65, 763–772 (2013).
Zoetendal, E. G. et al. The human small intestinal microbiota is driven by rapid uptake and conversion of simple carbohydrates. ISME J. 6, 1415–1426 (2012).
Gu, S. et al. Bacterial Community Mapping of the Mouse Gastrointestinal Tract. PLoS ONE 8, e74957 (2013).
Dubos, R., Schaedler, R. W., Costello, R. & Hoet, P. Indigenous, normal, and autochthonous flora of the gastrointestinal tract. J. Exp. Med. 122, 67–76 (1965).
Savage, D. C., Dubos, R. & Schaedler, R. W. The gastrointestinal epithelium and its autochthonous bacterial flora. J. Exp. Med. 127, 67–76 (1968).
Davis, C. P., Mulcahy, D., Takeuchi, A. & Savage, D. C. Location and Description of Spiral-Shaped Microorganisms in the Normal Rat Cecum. Infect. Immun. 6, 184–192 (1972).
Galley, J. D. et al. The structures of the colonic mucosa-associated and luminal microbial communities are distinct and differentially affected by a prolonged murine stressor. Gut Microbes 5, 748–760 (2014).
Yasuda, K. et al. Biogeography of the Intestinal Mucosal and Lumenal Microbiome in the Rhesus Macaque. Cell Host Microbe 17, 385–391 (2015).
Adhikari, B., Kim, S. W. & Kwon, Y. M. Characterization of Microbiota Associated with Digesta and Mucosa in Different Regions of Gastrointestinal Tract of Nursery Pigs. Int. J. Mol. Sci. 20, 1630 (2019).
Nava, G. M., Friedrichsen, H. J. & Stappenbeck, T. S. Spatial organization of intestinal microbiota in the mouse ascending colon. ISME J. 5, 627–638 (2011).
Liu, L. et al. Establishing a mucosal gut microbial community in vitro using an artificial simulator. PLOS ONE 13, e0197692 (2018).
Stedman, A., Brunner, K. & Nigro, G. Decrypting the communication between microbes and the intestinal mucosa—A brief review on Pathogénie Microbienne Moléculaire’s latest research. Cell. Microbiol. 0, e13118 (2019).
Pédron, T. et al. A Crypt-Specific Core Microbiota Resides in the Mouse Colon. mBio 3, e00116–12 (2012).
Saffarian, A. et al. Crypt- and Mucosa-Associated Core Microbiotas in Humans and Their Alteration in Colon Cancer Patients. mBio 10, e01315–e01319 (2019).
Naito, T. et al. Lipopolysaccharide from Crypt-Specific Core Microbiota Modulates the Colonic Epithelial Proliferation-to-Differentiation Balance. mBio 8, e01680–17 (2017).
Milani, C. et al. The First Microbial Colonizers of the Human Gut: composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 81, e00036–17 (2017).
Perez-Muñoz, M. E., Arrieta, M.-C., Ramer-Tait, A. E. & Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: implications for research on the pioneer infant microbiome. Microbiome 5, 48 (2017).
La Rosa, P. S. et al. Patterned progression of bacterial populations in the premature infant gut. Proc. Natl Acad. Sci. USA 111, 12522–12527 (2014).
Palmer, C., Bik, E. M., DiGiulio, D. B., Relman, D. A. & Brown, P. O. Development of the Human Infant Intestinal Microbiota. PLOS Biol. 5, e177 (2007).
Dominguez-Bello, M. G. et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. PNAS 107, 11971–11975 (2010).
Selma-Royo, M. et al. Perinatal environment shapes microbiota colonization and infant growth: impact on host response and intestinal function. Microbiome 8, 167 (2020).
Bergström, A. et al. Establishment of Intestinal Microbiota during Early Life: a Longitudinal, Explorative Study of a Large Cohort of Danish Infants. Appl Environ. Microbiol 80, 2889–2900 (2014).
Tanaka, M. & Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 66, 515–522 (2017).
Laursen, M. F., Bahl, M. I., Michaelsen, K. F. & Licht, T. R. First Foods and Gut Microbes. Front. Microbiol. 8, 356 (2017).
Al Nabhani, Z. et al. A Weaning Reaction to Microbiota Is Required for Resistance to Immunopathologies in the Adult. Immunity 50, 1276–1288.e5 (2019).
Al Nabhani, Z. & Eberl, G. Imprinting of the immune system by the microbiota early in life. Mucosal. Immunol. 13, 183–189 (2020).
Martínez, I., Muller, C. E. & Walter, J. Long-Term Temporal Analysis of the Human Fecal Microbiota Revealed a Stable Core of Dominant Bacterial Species. PLoS ONE 8, e69621 (2013).
Faith, J. J. et al. The long-term stability of the human gut microbiota. Science 341, 1237439 (2013).
Quercia, S. et al. Gut microbiome response to short-term dietary interventions in reactive hypoglycemia subjects. Diabetes/Metab. Res. Rev. 33, e2927 (2017).
Jiang, C., Li, G., Huang, P., Liu, Z. & Zhao, B. The Gut Microbiota and Alzheimer’s Disease. J. Alzheimer’s Dis. 58, 1–15 (2017).
Aleman, F. D. D. & Valenzano, D. R. Microbiome evolution during host aging. PLoS Pathog 15, e1007727 (2019).
Biver, E. et al. Gut microbiota and osteoarthritis management: An expert consensus of the European society for clinical and economic aspects of osteoporosis, osteoarthritis and musculoskeletal diseases (ESCEO). Ageing Res. Rev. 55, 100946 (2019).
Kaczmarek, J. L., Musaad, S. M. & Holscher, H. D. Time of day and eating behaviors are associated with the composition and function of the human gastrointestinal microbiota. Am. J. Clin. Nutr. ajcn156380 https://doi.org/10.3945/ajcn.117.156380 (2017).
Parkar, S. G., Kalsbeek, A. & Cheeseman, J. F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 7, 41 (2019).
Voigt, R. M. et al. Circadian Disorganization Alters Intestinal Microbiota. PLoS ONE 9, e97500 (2014).
Deaver, J. A., Eum, S. Y. & Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front. Microbiol. 9, 737 (2018).
Thaiss, C. A. et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 167, 1495–1510.e12 (2016).
Chassaing, B. et al. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 519, 92–96 (2015).
Chassaing, B., Raja, S. M., Lewis, J. D., Srinivasan, S. & Gewirtz, A. T. Colonic Microbiota Encroachment Correlates With Dysglycemia in Humans. Cell. Mol. Gastroenterol. Hepatol. 4, 205–221 (2017).
Cani, P. D. et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 56, 1761–1772 (2007).
Cani, P. D. et al. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet–Induced Obesity and Diabetes in Mice. Diabetes 57, 1470–1481 (2008).
Ng, S. C. et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet 390, 2769–2778 (2017).
Kostic, A. D., Xavier, R. J. & Gevers, D. The Microbiome in Inflammatory Bowel Diseases: current Status and the Future Ahead. Gastroenterology 146, 1489–1499 (2014).
Libertucci, J. et al. Inflammation-related differences in mucosa-associated microbiota and intestinal barrier function in colonic Crohn’s disease. Am. J. Physiol-Gastrointest. Liver Physiol. https://doi.org/10.1152/ajpgi.00411.2017 (2018).
Britton, G. J. et al. Microbiotas from Humans with Inflammatory Bowel Disease Alter the Balance of Gut Th17 and RORγt+ Regulatory T Cells and Exacerbate Colitis in Mice. Immunity 50, 212–224.e4 (2019).
Zhong, W. et al. Distinct Microbial Populations Exist in the Mucosa-associated Microbiota of Diarrhea Predominant Irritable Bowel Syndrome and Ulcerative Colitis. J. Clin. Gastroenterol. 53, 660–672 (2019).
Manichanh, C. et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 55, 205–211 (2006).
Sokol, H. et al. Low Counts of Faecalibacterium prausnitzii in Colitis Microbiota. Inflamm. Bowel Dis. 15, 1183–1189 (2009).
Ott, S. J. et al. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 53, 685–693 (2004).
Sokol, H. et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl Acad. Sci. 105, 16731–16736 (2008).
Zhao, H. et al. Systematic Review and Meta-Analysis of the Role of Faecalibacterium prausnitzii Alteration in Inflammatory Bowel Disease. J. Gastroenterol. Hepatol. Online ahead of print.
Swidsinski, A., Loening–Baucke, V., Verstraelen, H., Osowska, S. & Doerffel, Y. Biostructure of Fecal Microbiota in Healthy Subjects and Patients With Chronic Idiopathic Diarrhea. Gastroenterology 135, 568–579.e2 (2008).
Shaler, C. R., Elhenawy, W. & Coombes, B. K. The Unique Lifestyle of Crohn’s Disease-Associated Adherent-Invasive Escherichia coli. J. Mol. Biol. 431, 2970–2981 (2019).
Darfeuille-Michaud, A. et al. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 127, 412–421 (2004).
Baumgart, M. et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 1, 403–418 (2007).
Kotlowski, R., Bernstein, C. N., Sepehri, S. & Krause, D. O. High prevalence of Escherichia coli belonging to the B2+D phylogenetic group in inflammatory bowel disease. Gut 56, 669–675 (2007).
Lupp, C. et al. Host-Mediated Inflammation Disrupts the Intestinal Microbiota and Promotes the Overgrowth of Enterobacteriaceae. Cell Host Microbe 2, 119–129 (2007).
Gibold, L. et al. The Vat-AIEC protease promotes crossing of the intestinal mucus layer by Crohn’s disease-associated Escherichia coli. Cell. Microbiol. 18, 617–631 (2016).
Sevrin, G. et al. Adaptation of adherent-invasive E. coli to gut environment: Impact on flagellum expression and bacterial colonization ability. Gut Microbes 11, 364–380 (2020).
Sokol, H. et al. Prominence of ileal mucosa-associated microbiota to predict postoperative endoscopic recurrence in Crohn’s disease. Gut 69, 462–472 (2020).
Yang, M. et al. Duodenal and rectal mucosal microbiota related to small intestinal bacterial overgrowth in diarrhea-predominant irritable bowel syndrome. J. Gastroenterol. Hepatol. 35, 795–805 (2020).
Jabbar, K. S. et al. Association between Brachyspira and irritable bowel syndrome with diarrhoea. Gut https://doi.org/10.1136/gutjnl-2020-321466 (2020).
World Health Organization, International Agency for Research on Cancer Cancer today. at http://gco.iarc.fr/today/home.
Gagnaire, A., Nadel, B., Raoult, D., Neefjes, J. & Gorvel, J.-P. Collateral damage: insights into bacterial mechanisms that predispose host cells to cancer. Nat. Rev. Microbiol. 15, 109–128 (2017).
Allen, J. & Sears, C. L. Impact of the gut microbiome on the genome and epigenome of colon epithelial cells: contributions to colorectal cancer development. Genome Med. 11, 1–18 (2019).
Cuevas-Ramos, G. et al. Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc. Natl Acad. Sci. USA 107, 11537–11542 (2010).
Arthur, J. C. et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 338, 120–123 (2012).
Castellarin, M. et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 22, 299–306 (2012).
Dejea, C. M. et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl Acad. Sci. USA 111, 18321–18326 (2014).
Pleguezuelos-Manzano, C. et al. Mutational signature in colorectal cancer caused by genotoxic pks + E. coli. Nature 580, 269–273 (2020).
Chen, W., Liu, F., Ling, Z., Tong, X. & Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLOS ONE 7, e39743 (2012).
Gao, R. et al. Mucosa-associated microbiota signature in colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 36, 2073–2083 (2017).
Tomkovich, S. et al. Locoregional effects of microbiota in a preclinical model of colon carcinogenesis. Cancer Res. 77, 2620–2632 (2017).
Tomkovich, S. et al. Human colon mucosal biofilms from healthy or colon cancer hosts are carcinogenic. J. Clin. Investig. 129, 1699–1712 (2019).
Tomkovich, S. et al. Human Colon Mucosal Biofilms and Murine Host Communicate via Altered mRNA and microRNA Expression during Cancer. mSystems 5, e00451–19 (2020).
Caselli, M., Holton, J., Boldrini, P., Vaira, D. & Calò, G. Morphology of segmented filamentous bacteria and their patterns of contact with the follicle-associated epithelium of the mouse terminal ileum: implications for the relationship with the immune system. Gut Microbes 1, 367–372 (2010).
Davis, C. P. & Savage, D. C. Habitat, Succession, Attachment, and Morphology of Segmented, Filamentous Microbes Indigenous to the Murine Gastrointestinal Tract. Infect. Immun. 10, 948–956 (1974).
Chase, D. G. & Erlandsen, S. L. Evidence for a complex life cycle and endospore formation in the attached, filamentous, segmented bacterium from murine ileum. J. Bacteriol. 127, 572–583 (1976).
Jepson, M. A., Clark, M. A., Simmons, N. L. & Hirst, B. H. Actin accumulation at sites of attachment of indigenous apathogenic segmented filamentous bacteria to mouse ileal epithelial cells. Infect. Immun. 61, 4001–4004 (1993).
Yin, Y. et al. Comparative analysis of the distribution of segmented filamentous bacteria in humans, mice and chickens. ISME J. 7, 615–621 (2013).
Klaasen, H. L. B. M., Koopman, J. P., Scholten, P. M., Brink, M. E. V. D. & Theeuwes, A. G. M. Effect of Preventing Coprophagy on Colonisation by Segmented Filamentous Bacteria in the Small Bowel of Mice. Microb. Ecol. Health Dis. 3, 99–103 (1990).
Snel, J. et al. Interactions between gut-associated lymphoid tissue and colonization levels of indigenous, segmented, filamentous bacteria in the small intestine of mice. Can. J. Microbiol. 44, 1177–1182 (1998).
Schnupf, P., Gaboriau-Routhiau, V. & Cerf-Bensussan, N. Host interactions with Segmented Filamentous Bacteria: An unusual trade-off that drives the post-natal maturation of the gut immune system. Semin. Immunol. 25, 342–351 (2013).
Klaasen, H. L. et al. Apathogenic, intestinal, segmented, filamentous bacteria stimulate the mucosal immune system of mice. Infect. Immun. 61, 303–306 (1993).
Talham, G. L., Jiang, H.-Q., Bos, N. A. & Cebra, J. J. Segmented Filamentous Bacteria Are Potent Stimuli of a Physiologically Normal State of the Murine Gut Mucosal Immune System. Infect. Immun. 67, 1992–2000 (1999).
Lécuyer, E. et al. Segmented Filamentous Bacterium Uses Secondary and Tertiary Lymphoid Tissues to Induce Gut IgA and Specific T Helper 17 Cell Responses. Immunity 40, 608–620 (2014).
Gaboriau-Routhiau, V. et al. The Key Role of Segmented Filamentous Bacteria in the Coordinated Maturation of Gut Helper T Cell Responses. Immunity 31, 677–689 (2009).
Ivanov, I. I. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139, 485–498 (2009).
Garland, C. D., Lee, A. & Dickson, M. R. Segmented filamentous bacteria in the rodent small intestine: Their colonization of growing animals and possible role in host resistance toSalmonella. Micro. Ecol. 8, 181–190 (1982).
Shi, Z. et al. Segmented Filamentous Bacteria Prevent and Cure Rotavirus Infection. Cell 179, 644–658.e13 (2019).
Schnupf, P. et al. Growth and host interaction of mouse Segmented Filamentous Bacteria in vitro. Nature 520, 99–103 (2015).
Huang, J. Y., Lee, S. M. & Mazmanian, S. K. The human commensal Bacteroides fragilis binds intestinal mucin. Anaerobe 17, 137–141 (2011).
Lee, S. M. et al. Bacterial colonization factors control specificity and stability of the gut microbiota. Nature 501, 426–429 (2013).
Donaldson, G. P. et al. Gut microbiota utilize immunoglobulin A for mucosal colonization. Science 360, 795–800 (2018).
Donaldson, G. P. et al. Spatially distinct physiology of Bacteroides fragilis within the proximal colon of gnotobiotic mice. Nat. Microbiol. 5, 746–756 (2020).
Mazmanian, S. K., Liu, C. H., Tzianabos, A. O. & Kasper, D. L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 122, 107–118 (2005).
Shen, Y. et al. Outer Membrane Vesicles of a Human Commensal Mediate Immune Regulation and Disease Protection. Cell Host Microbe 12, 509–520 (2012).
Mazmanian, S. K., Round, J. L. & Kasper, D. L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 453, 620–625 (2008).
Lee, Y. K. et al. The Protective Role of Bacteroides fragilis in a Murine Model of Colitis-Associated Colorectal Cancer. mSphere 3, e00587–18 (2018).
Ramakrishna, C. et al. Bacteroides fragilis polysaccharide A induces IL-10 secreting B and T cells that prevent viral encephalitis. Nat. Commun. 10, 2153 (2019).
Rigottier-Gois, L. Dysbiosis in inflammatory bowel diseases: the oxygen hypothesis. ISME J. 7, 1256–1261 (2013).
Albenberg, L. et al. Correlation Between Intraluminal Oxygen Gradient and Radial Partitioning of Intestinal Microbiota. Gastroenterology 147, 1055–1063.e8 (2014).
Marteyn, B. et al. Modulation of Shigella virulence in response to available oxygen in vivo. Nature 465, 355–358 (2010).
Rivera-Chávez, F., Lopez, C. A. & Bäumler, A. J. Oxygen as a driver of gut dysbiosis. Free Radic. Biol. Med. 105, 93–101 (2017).
Manichanh, C. et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 55, 205–211 (2006).
Frank, D. N. et al. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl Acad. Sci. USA 104, 13780–13785 (2007).
Godinez, I. et al. T Cells Help To Amplify Inflammatory Responses Induced by Salmonella enterica Serotype Typhimurium in the Intestinal Mucosa. Infect. Immun. 76, 2008–2017 (2008).
Rivera-Chávez, F. et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe 19, 443–454 (2016).
Atarashi, K. et al. T reg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 500, 232–236 (2013).
Donohoe, D. R. et al. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 13, 517–526 (2011).
Velázquez, O. C., Lederer, H. M. & Rombeau, J. L. Butyrate and the Colonocyte. in Dietary Fiber in Health and Disease (Kritchevsky, D. & Bonfield, C.) 123–134 (Springer US, Boston, MA, 1997). https://doi.org/10.1007/978-1-4615-5967-2_14.
Furuta, G. T. et al. Hypoxia-Inducible Factor 1–Dependent Induction of Intestinal Trefoil Factor Protects Barrier Function during Hypoxia. J. Exp. Med. 193, 1027–1034 (2001).
Faber, F. et al. Host-mediated sugar oxidation promotes post-antibiotic pathogen expansion. Nature 534, 697–699 (2016).
Miller, B. M. et al. Anaerobic Respiration of NOX1-Derived Hydrogen Peroxide Licenses Bacterial Growth at the Colonic Surface. Cell Host Microbe 28, 789–797.e5 (2020).
Popkin, B. M. Nutrition Transition and the Global Diabetes Epidemic. Curr. Diab. Rep. 15, 64 (2015).
Zhao, L. The gut microbiota and obesity: from correlation to causality. Nat. Rev. Microbiol. 11, 639–647 (2013).
Sonnenburg, J. L. & Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 535, 56–64 (2016).
Tomas, J. et al. High-fat diet modifies the PPAR-γ pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. Proc. Natl Acad. Sci. 113, E5934–E5943 (2016).
Schroeder, B. O. et al. Bifidobacteria or Fiber Protects against Diet-Induced Microbiota-Mediated Colonic Mucus Deterioration. Cell Host Microbe 23, 27–40.e7 (2018).
Montrose, D. C. et al. Dietary Fructose Alters the Composition, Localization, and Metabolism of Gut Microbiota in Association With Worsening Colitis. Cellular Mol. Gastroenterol. Hepatol. 11, 525–550 (2020).
Veronese, N. et al. Dietary fiber and health outcomes: an umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 107, 436–444 (2018).
Sonnenburg, E. D. & Sonnenburg, J. L. Starving our Microbial Self: The Deleterious Consequences of a Diet Deficient in Microbiota-Accessible Carbohydrates. Cell Metab. 20, 779–786 (2014).
Chassaing, B. et al. Lack of soluble fiber drives diet-induced adiposity in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 309, G528–G541 (2015).
Cani, P. D. et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 50, 2374–2383 (2007).
Delzenne, N. M., Cani, P. D. & Neyrinck, A. M. Modulation of Glucagon-like Peptide 1 and Energy Metabolism by Inulin and Oligofructose: Experimental Data. J. Nutr. 137, 2547S–2551S (2007).
Everard, A. et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and Diet-Induced Leptin-Resistant Mice. Diabetes 60, 2775–2786 (2011).
Everard, A. et al. Microbiome of prebiotic-treated mice reveals novel targets involved in host response during obesity. ISME J. 8, 2116–2130 (2014).
Desai, M. S. et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 167, 1339–1353.e21 (2016).
Sonnenburg, E. D. et al. Diet-induced extinctions in the gut microbiota compound over generations. Nature 529, 212–215 (2016).
Swidsinski, A. et al. Bacterial Overgrowth and Inflammation of Small Intestine After Carboxymethylcellulose Ingestion in Genetically Susceptible Mice. Inflamm. Bowel Dis. 15, 359–364 (2009).
Roberts, C. L., Rushworth, S. L., Richman, E. & Rhodes, J. M. Hypothesis: increased consumption of emulsifiers as an explanation for the rising incidence of Crohn’s disease. J. Crohn’s Colitis 7, 338–341 (2013).
Chassaing, B., Van de Wiele, T., De Bodt, J., Marzorati, M. & Gewirtz, A. T. Dietary emulsifiers directly alter human microbiota composition and gene expression ex vivo potentiating intestinal inflammation. Gut 66, 1414–1427 (2017).
Viennois, E. et al. Dietary Emulsifiers Directly Impact Adherent-Invasive E. coli Gene Expression to Drive Chronic Intestinal Inflammation. Cell Rep. 33, 108229 (2020).
Derrien, M., Vaughan, E. E., Plugge, C. M. & Vos, W. M. de Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evolut. Microbiol. 54, 1469–1476 (2004).
Berry, D. et al. Host-compound foraging by intestinal microbiota revealed by single-cell stable isotope probing. PNAS 110, 4720–4725 (2013).
Geerlings, S. Y., Kostopoulos, I., Vos, W. Mde & Belzer, C. Akkermansia muciniphila In the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 6, 75 (2018).
Everard, A. et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl Acad. Sci. U. S. A. 110, 9066–9071 (2013).
Dao, M. C. et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 65, 426–436 (2016).
Dao, M. C. et al. Akkermansia muciniphila abundance is lower in severe obesity but its increased level after bariatric surgery is not associated with metabolic health improvement. Am. J. Physiol-Endocrinol. Metab. (2019). https://doi.org/10.1152/ajpendo.00140.2019
Shin, N.-R. et al. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 63, 727–735 (2014).
Shin, J. et al. Elucidation of Akkermansia muciniphila Probiotic Traits Driven by Mucin Depletion. Front. Microbiol. 10, 1137 (2019).
Hänninen, A. et al. Akkermansia muciniphila induces gut microbiota remodelling and controls islet autoimmunity in NOD mice. Gut 67, 1445–1453 (2018).
Reunanen, J. et al. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 81, 3655–3662 (2015).
Ottman, N. et al. Genome-Scale Model and Omics Analysis of Metabolic Capacities of Akkermansia muciniphila Reveal a Preferential Mucin-Degrading Lifestyle. Appl. Environ. Microbiol. 83, e01014–e01017 (2017).
Plovier, H. et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 23, 107–113 (2017).
Chelakkot, C. et al. Akkermansia muciniphila -derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med 50, e450–e450 (2018).
Depommier, C. et al. Pasteurized Akkermansia muciniphila increases whole-body energy expenditure and fecal energy excretion in diet-induced obese mice. Gut Microbes 11, 1231–1245 (2020).
Druart, C. et al. Toxicological safety evaluation of pasteurized Akkermansia muciniphila. J. Appl. Toxicol. https://doi.org/10.1002/jat.4044 (2020).
Depommier, C. et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 25, 1096 (2019).
Liew, F. Y., Pitman, N. I. & McInnes, I. B. Disease-associated functions of IL-33: the new kid in the IL-1 family. Nat. Rev. Immunol. 10, 103–110 (2010).
Monticelli, L. A. et al. IL-33 promotes an innate immune pathway of intestinal tissue protection dependent on amphiregulin–EGFR interactions. PNAS 112, 10762–10767 (2015).
Hodzic, Z., Schill, E. M., Bolock, A. M. & Good, M. IL-33 and the intestine: the good, the bad, and the inflammatory. Cytokine 100, 1–10 (2017).
Lopetuso, L. R. et al. IL-33 promotes recovery from acute colitis by inducing miR-320 to stimulate epithelial restitution and repair. Proc. Natl Acad. Sci. USA 115, E9362–E9370 (2018).
Moussion, C., Ortega, N. & Girard, J.-P. The IL-1-Like Cytokine IL-33 Is Constitutively Expressed in the Nucleus of Endothelial Cells and Epithelial Cells In Vivo: a Novel ‘Alarmin’? PLoS ONE 3, e3331 (2008).
Oboki, K. et al. IL-33 is a crucial amplifier of innate rather than acquired immunity. PNAS 107, 18581–18586 (2010).
Waddell, A. et al. IL-33 signaling protects from murine oxazolone colitis by supporting intestinal epithelial function. Inflamm. Bowel Dis. 21, 2737–2746 (2015).
Malik, A. et al. IL-33 regulates the IgA-microbiota axis to restrain IL-1α–dependent colitis and tumorigenesis. J. Clin. Investig. 126, 4469–4481 (2016).
Hodzic, Z., Schill, E. M., Bolock, A. M. & Good, M. IL-33 and the intestine: the good, the bad, and the inflammatory. Cytokine 100, 1–10 (2017).
Schmitz, J. et al. IL-33, an Interleukin-1-like Cytokine that Signals via the IL-1 Receptor-Related Protein ST2 and Induces T Helper Type 2-Associated Cytokines. Immunity 23, 479–490 (2005).
Groβ, P., Doser, K., Falk, W., Obermeier, F. & Hofmann, C. IL-33 Attenuates Development and Perpetuation of Chronic Intestinal Inflammation. Inflamm. Bowel Dis. 18, 1900–1909 (2012).
Waddell, A., Vallance, J. E., Hummel, A., Alenghat, T. & Rosen, M. J. IL-33 Induces Murine Intestinal Goblet Cell Differentiation Indirectly via Innate Lymphoid Cell IL-13 Secretion. J. Immunol. 202, 598–607 (2019).
Frisbee, A. L. et al. IL-33 drives group 2 innate lymphoid cell-mediated protection during Clostridium difficile infection. Nat. Commun. 10, 2712 (2019).
Neill, D. R. et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 464, 1367–1370 (2010).
Moro, K. et al. Innate production of T H 2 cytokines by adipose tissue-associated c-Kit + Sca-1 + lymphoid cells. Nature 463, 540–544 (2010).
Hoyler, T. et al. The transcription factor GATA3 controls cell fate and maintenance of type 2 innate lymphoid cells. Immunity 37, 634–648 (2012).
Shimokawa, C. et al. Mast Cells Are Crucial for Induction of Group 2 Innate Lymphoid Cells and Clearance of Helminth Infections. Immunity 46, 863–874.e4 (2017).
Mahapatro, M. et al. Programming of Intestinal Epithelial Differentiation by IL-33 Derived from Pericryptal Fibroblasts in Response to Systemic Infection. Cell Rep. 15, 1743–1756 (2016).
Satoh-Takayama, N. et al. Bacteria-Induced Group 2 Innate Lymphoid Cells in the Stomach Provide Immune Protection through Induction of IgA. Immunity 52, 635–649.e4 (2020).
Pushparaj, P. N. et al. Interleukin-33 exacerbates acute colitis via interleukin-4 in mice. Immunology 140, 70–77 (2013).
Qiu, X., Qi, C., Li, X., Fang, D. & Fang, M. IL-33 deficiency protects mice from DSS-induced experimental colitis by suppressing ILC2 and Th17 cell responses. Inflamm. Res. 69, 1111–1122 (2020).
Mishima, Y. et al. Interleukin-33 delays recovery of mucosal inflammation via downregulation of homeostatic ABCG5/8 in the colon. Lab. Investig. 100, 491–502 (2020).
Cullender, T. C. et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 14, 571–581 (2013).
Chassaing, B. & Gewirtz, A. T. Identification of Inner Mucus-Associated Bacteria by Laser Capture Microdissection. Cell Mol. Gastroenterol. Hepatol. 7, 157–160 (2018).
Wang, Y. et al. Laser capture microdissection and metagenomic analysis of intact mucosa-associated microbial communities of human colon. Appl. Microbiol. Biotechnol. 88, 1333–1342 (2010).
Nava, G. M. & Stappenbeck, T. S. Diversity of the autochthonous colonic microbiota. Gut Microbes 2, 99–104 (2011).
Wrzosek, L. et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 11, 61 (2013).
Rossi, O. et al. Faecalibacterium prausnitzii Strain HTF-F and Its Extracellular Polymeric Matrix Attenuate Clinical Parameters in DSS-Induced Colitis. PLoS ONE 10, e0123013 (2015).
Martín, R., Bermúdez-Humarán, L. G. & Langella, P. Searching for the Bacterial Effector: the Example of the Multi-Skilled Commensal Bacterium Faecalibacterium prausnitzii. Front. Microbiol. 9, 346 (2018).
Gibold, L. et al. The Vat-AIEC protease promotes crossing of the intestinal mucus layer by Crohn’s disease-associated Escherichia coli. Cell Microbiol. 18, 617–631 (2016).
Acknowledgements
B.C.’s laboratory is supported by a Starting Grant from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (Grant agreement No. ERC-2018-StG- 804135), an Innovator Award from the Kenneth Rainin Foundation, and a Chaire d’Excellence from IdEx Université de Paris - ANR-18-IDEX-0001.
Author information
Authors and Affiliations
Contributions
N.D., E.L. and B.C. wrote the paper. N.D. and B.C. designed the figure.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Daniel, N., Lécuyer, E. & Chassaing, B. Host/microbiota interactions in health and diseases—Time for mucosal microbiology!. Mucosal Immunol 14, 1006–1016 (2021). https://doi.org/10.1038/s41385-021-00383-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41385-021-00383-w
This article is cited by
-
Chronic intestinal pseudo-obstruction: associations with gut microbiota and genes expression of intestinal serotonergic pathway
BMC Microbiology (2024)
-
Ultra-processed foods and food additives in gut health and disease
Nature Reviews Gastroenterology & Hepatology (2024)
-
Sialidases and fucosidases of Akkermansia muciniphila are crucial for growth on mucin and nutrient sharing with mucus-associated gut bacteria
Nature Communications (2023)
-
Alterations of mucosa-attached microbiome and epithelial cell numbers in the cystic fibrosis small intestine with implications for intestinal disease
Scientific Reports (2022)
-
Role of mucus-bacteria interactions in Enterotoxigenic Escherichia coli (ETEC) H10407 virulence and interplay with human microbiome
npj Biofilms and Microbiomes (2022)
