
Abstract
Transforming growth factor-β (TGF-β) plays a central role in hepatic fibrogenesis. This study investigated the function and mechanism of bone morphogenetic protein-2 (BMP-2) in regulation of hepatic fibrogenesis. BMP-2 expression in fibrotic liver was measured in human tissue microarray and mouse models of liver fibrosis induced by bile duct ligation surgery or carbon tetrachloride administration. Adenovirus-mediated BMP-2 gene delivery was used to test the prophylactic effect on liver fibrosis. Primary hepatic stellate cells (HSC), HSC-T6 and clone-9 cell lines were used to study the interplay between BMP-2 and TGF-β1. Hepatic BMP-2 was localized in parenchymal hepatocytes and activated HSCs and significantly decreased in human and mouse fibrotic livers, showing an opposite pattern of hepatic TGF-β1 contents. BMP-2 gene delivery alleviated the elevations of serum hepatic enzymes, cholangiocyte marker CK19, HSC activation markers, and liver fibrosis in both models. Mechanistically, exogenous TGF-β1 dose dependently reduced BMP-2 expression, whereas BMP-2 significantly suppressed expression of TGF-β and its cognate type I and II receptor peptides, as well as the induced Smad3 phosphorylation levels in primary mouse HSCs. Aside from its suppressive effects on cell proliferation and migration, BMP-2 treatment prominently attenuated the TGF-β1-stimulated α-SMA and fibronectin expression, and reversed the TGF-β1-modulated epithelial-to-mesenchymal transition marker expression in mouse HSCs. The mutual regulation between BMP-2 and TGF-β1 signaling axes may constitute the anti-fibrogenic mechanism of BMP-2 in the pathogenesis of liver fibrosis. BMP-2 may potentially serve as a novel therapeutic target for treatment of liver fibrosis.
Similar content being viewed by others
Introduction
Hepatic fibrosis is a dynamic process mediated by chronic liver injury of various etiologies. The main feature of fibrosis is increased deposition of extracellular matrix (ECM) by activated hepatic stellate cells (HSCs) [1,2,3]. During chronic liver injury, HSCs lose their intracellular lipid and retinoid droplets and transdifferentiate from quiescent cells to myofibroblast-like cells that express α-smooth muscle actin (α-SMA). Simultaneously, activated HSCs increase proliferation rate and produce excessive ECM molecules, including fibrillar type I collagen and fibronectin [1, 3,4,5]. In the context of hepatic fibrogenesis, transforming growth factor-β (TGF-β) is known to induce myofibroblastic transition and ECM overproduction of HSCs [3, 6], and has been proved to be a vital mediator of fibrogenesis in many chronic liver diseases [7]. The high expression of TGF-β1 not only contributes to recruitment of inflammatory cells and fibroblasts into the injured area and subsequent activation of HSCs, but also leads to epithelial-to-mesenchymal transition (EMT), which is a form of epithelial plasticity important in hepatic fibrogenesis [8,9,10,11]. Mechanistically, TGF-β1 induces activation of numerous signaling pathways that result in the loss of expression of epithelial markers such as E-cadherin and the increased de novo expression of mesenchymal markers such as vimentin and fibronectin [8, 11, 12]. Conversely, targeting TGF-β1 signal has been claimed to have therapeutic benefits in experimental and clinical liver fibrosis diseases, aiming for not only preventing fibrosis progression, but also preserving liver function [10, 13].
Bone morphogenetic proteins (BMPs) are a group of growth factors classified into TGF-β superfamily, and initially identified as morphogenetic factors in bone development [14, 15]. All members of the TGF-β superfamily signal through highly conserved transmembrane receptors and Smads [16]. TGF-β ligand is typically coupled and forms a complex with cell surface type I and type II receptors [17]. The ligand–receptor complex recruits and causes phosphorylation of Smad2 and Smad3 proteins, followed by formation of a heterotrimeric complex with Smad4 and its subsequent nuclear translocation [18]. Distinct from the TGF-β-driven signaling machinery, BMPs convey signals toward nucleus through a different set of Smad proteins, including Smad1, Smad5, and Smad8 [16]. The similarities of ligand structure and receptor-binding modality as well as the co-utilization of Smad4 protein between BMP and TGF-β signaling cascades strongly suggest that BMPs are very likely to participate in not only liver development but also liver fibrogenesis through dynamic competition with TGF-β. Indeed, earlier studies of liver development have demonstrated that bone morphogenetic protein (BMP)-9 plays a hepatogenic role by stimulating hepatocyte proliferation [19] and that BMP-4 signaling is morphogenetically essential for the growth of hepatic endoderm into a liver bud [20], and vital for the three-dimensional assembly of hepatic cords from hepatoblasts and mesenchymal cells [21]. By contrast, BMP-2 is found to be a negative regulator of hepatocyte proliferation and downregulated in the regenerating liver [22]. Recent studies using a rat unilateral urethral obstruction model demonstrate that BMP-2 exerts therapeutic effect on renal interstitial fibrosis via antagonizing TGF-β1-induced profibrogenic signals and EMT in interstitial fibroblasts [23, 24], implicating a negative regulatory role of BMP-2 in organ fibrogenesis. Other lines of evidence also indicates that BMP-2 may suppress the TGF-β1-induced myofibroblastic differentiation of lung fibroblasts [25] and pancreatic stellate cells [26]. However, the regulatory role of BMP-2 in hepatic fibrogenesis remains elusive. Therefore, this study examined the expression profiles of BMP-2 and TGF-β1 in human and mouse fibrotic livers, and evaluated the prophylactic efficacy of BMP-2 gene delivery in two mouse models of liver fibrosis induced by bile duct ligation (BDL) surgery and carbon tetrachloride (CCl4) intoxication, which emulate acute and chronic hepatic damages, respectively [27]. The regulatory mechanism of BMP-2 in TGF-β1/Smad cascade was further elucidated in a HSC-T6 cell line and primary mouse HSCs.
Materials and methods
Human liver tissue microarray (TMA)
The human liver TMA consisting of normal livers (n = 30), chronic hepatitis (n = 10), and cirrhosis livers (n = 40) was purchased from US Biomax, Inc. (Kit no. LV805a, Rockville, MD). The TMA slides were subjected immunohistochemistry (IHC) staining of TGF-β1 and BMP-2, and quantified by using the HistoQuest immunohistochemistry analysis software (TissueGnostics) [28].
Animal models and study ethics
The animal study protocol was reviewed and approved by the Institutional Animal Care and Use Committee of the Chang Gung Memorial Hospital (Approval no. 2016092604). FVB male mice (age 7–8 weeks) weighing 25–35 g were purchased from BioLASCO Co., Ltd. (Taipei, Taiwan). All animals were housed in an animal facility (set at 22 °C, with a relative humidity of 55%, and in a 12h-light/12h-dark cycle) with sterile tap water and food available ad libitum. Mouse liver fibrosis was induced by conducting common BDL surgery or subcutaneous administration of CCl4 as previously described [27]. Liver sera and tissues were collected at 1 and 2 weeks after BDL, or at 2 and 4 weeks after initiation of CCl4 injection. Sera were subjected to biochemical parameter measurements by a standard clinical automatic analyzer, including alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), direct, and total bilirubin. Liver tissues were dissected, snap frozen, and stored at −80 ℃ for extracting total RNA, tissue lysates, and further molecular detection.
Cloning, expression, and purification of adenoviruses for gene delivery
To evaluate the anti-fibrotic effect of BMP-2, gene delivery administration was performed at 24 h prior to the day of CCl4 injection or BDL surgery. The replication-deficient, recombinant adenoviruses containing green fluorescent protein (Ad-GFP) and BMP-2 genes (Ad-BMP-2) were generated as previously described [29]. In brief, 30 μl of 1 × 109 plaque-forming units (pfu)/ml of Ad-GFP or Ad-BMP-2, which was stored in 10% glycerol and diluted in saline immediately before use, were injected into the tail veins using a 27 G needle under anesthesia.
Reverse transcription and quantitative polymerase chain reaction (RT-qPCR)
To quantify the mRNA levels of BMP-2 and TGF-β1 in the liver, we isolated total RNA performed RT reaction and qPCR as previously described [27]. The primer sequences were as follows: β-actin, 5’-TCACCCACACTGTGCCCATCTACGA-3’ (forward) and 5’-CAGCGGAACCGCTCATTGCCAATGG-3’ (reverse); TGF-β1, 5’-CGTCAGACATTCGGGAAGC-3’ (forward) and 5’-CAGCCACTCAGGCGTATCA-3’ (reverse); BMP-2, 5’-GGGTGGAATGACTGGATCG-3’ (forward) and 5’-ATGGCATGGTTAGTGGAGTT-3’ (reverse).
Western blot analysis
Protein lysates were prepared from mouse liver tissue samples or cells in 1 × radio immunoprecipitation assay lysis buffer (Cell Signaling Technology, Billerica, MA). Protein concentrations were measured using a protein assay dye (Bio-Rad Laboratories, Hercules, CA). SDS-PAGE and immunoblotting analysis were performed as described previously [30]. The detecting antibodies were raised against BMP-2 (ab6285, abcam, UK), TGF-β1 (ab92486, abcam), TGF-β1 receptor I (ab31013, abcam), TGF-β1 receptor II (ab186838, abcam), GAPDH (GTX627408, GeneTex, SA), phospho-Smad3 (pSmad3; ab51451, abcam), Smad3 (ab40854, abcam), fibronectin (ab2413, abcam), E-cadherin (ab76055, abcam), vimentin (2707-1 epitomics), and β-actin (sc-1615, Santa Cruz).
Histochemistry and IHC stainings
Formalin fixed, paraffin-embedded mouse liver sections were subjected to hematoxylin and eosin (H&E), Sirius red, IHC stainings using standard procedures as previously described [27]. For IHC, sections were incubated with anti-TGF-β1 or BMP-2 monoclonal antibodies (1:500 dilution) at 4 °C overnight. The color reaction was developed using HRP-linked polymer detection system (Biogenex, San Ramon, CA), followed by counterstaining with hematoxylin. Pre-immune rabbit serum was applied at equimolar concentration in normal liver sections as blank controls. Microphotographs were documented with a light microscope (BX50, Olympus Optical Company, Japan) equipped with a CCD digital camera (DP-70, Olympus).
Liver fibrosis score
The degree of hepatic fibrosis in mice was assessed by examining the H&E-stained sections and morphometrical analysis on Sirius red-stained sections was performed to METAVIR Scoring System. The liver fibrosis degree was classified into the following four grades on the basis of inflammation, necrosis, fibrosis and structural changes; F0: normal hepatic structure; F1: portal (without septa) fibrosis; F2: portal (with few septa) fibrosis; F3: septal fibrosis; F4: cirrhosis.
Immunofluorescence staining
The HSCs were cultured in six-well glass slide chambers for 24 h and further treated BMP-2 and TGF-β1 for 48 h. The cells were then fixed with 4% paraformaldehyde, permeabilized with 0.25% Triton X-100, and blocked with 3% bovine serum albumin for 30 min at room temperature. The fixed cells were then incubated with the primary antibodies against α-SMA (ab5694, abcam, UK), fibronectin, E-cadherin, vimentin, glial fibrillary acidic protein (GFAP; ab10062, abcam), albumin (GTX102419, GeneTex), and BMP-2 at 4 °C overnight followed by visualization with Alexa Fluor 488- (green) or Alexa Fluor 595 (red)-conjugated secondary antibodies at room temperature for 1 h. Nuclei were counterstained with 4',6-diamidino-2-phenylindole. The stained cells were mounted with a fluorescent mounting medium (Dako Cytomation) and observed by fluorescence microscopy (Olympus). The exposure gains and rates were consistent between samples. Fluorescent intensities were quantified on independent color channels by using Image J software (NIH, USA).
Cell lines, primary HSC isolation, and in vitro treatments
A normal rat hepatocyte cell line (clone-9), a rat liver stellate cell line (HSC-T6), and primary mouse HSCs were used for cytokine stimulation experiments. The clone-9 cell line (BCRC no. 60201) were purchased from Bioresource Collection and Research Center (Hsin-Chu, Taiwan) and maintained in F-12K medium (Sigma-Aldrich, St. Louis, MO) supplemented with 10 % heat-inactivated fetal calf serum at 37 °C in 5 % CO2-enriched air. The HSC-T6 cell line was a gift from Dr. Bruno Jawan (Kaohsiung Chang Gung Memorial Hospital, Taiwan) and maintained in Waymouth’s medium (Gibco BRL, Grand Island, NY) as previously described [31]. Both lines of cells (1 × 106/well) were stimulated with recombinant human TGF-β1 or BMP-2 (R&D Systems, Minneapolis, MN) at indicated doses.
Primary HSCs were isolated from mouse livers by sequential digestion with pronase and collagenase, followed by density gradient centrifugation in 8.5% Nycodenz (Sigma-Aldrich) as described previously [30, 32]. Cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 5% newborn calf serum. After 1 day in culture, the HSCs had a quiescent phenotype and they developed an activated phenotype after 7–14 days. All experiments were carried out using the cells between passages 2 and 6. The primary activated HSCs were pretreated with BMP-2 (250 ng/ml) for 1 h, followed by TGF-β1 exposure (10 ng/ml) for 48 h, and then subjected to cell proliferation and migration assays as well as monitoring the expression of fibrosis and EMT markers. Four independent in vitro experiments were performed.
Proliferation and migration assays
Cell proliferation and migration assays were performed as described previously [30]. For proliferation assay, a cell concentration of 5 × 103 cells/well was adopted for 48 h incubation. After the incubation period, 10 μl premixed WST-1 reagents (Roche Applied Science, Mannheim, Germany) was added into each well, and the cells were incubated for 4 h in a humidified atmosphere (e.g., 37℃ 5% CO2). Afterwards, the absorbance of the samples was measured at 450 nm against a background control with a reference wavelength higher than 600 nm on a microtiter plate (ELISA) reader.
For migration assay, images were taken at 0, 6, 12, and 15 h, and superimposed using PhotoImpact (Adobe). The number of cells that migrated into the wounded space were manually counted in three fields per well under a light microscope at × 40 magnifications. Areas were quantified by using Image J analysis software.
Statistical analysis
All values in the figures were expressed as mean ± standard error of the mean and analyzed by unpaired t test. P value < 0.05 was considered to be statistically significant.
Results
Downregulation of BMP-2 in human fibrotic livers
To investigate the expression of BMP-2 and its relationship with TGF-β1 in liver fibrosis, immunohistochemical analysis of BMP-2 and TGF-β1 protein expression was performed on human liver TMA slides containing normal, chronic hepatitis, and cirrhotic liver samples. It was found that BMP-2 immunostaining was prominent in all normal liver samples with strong immunoreactivity, whereas the signals became weaker in the hepatitis group and much weaker in the cirrhosis liver tissues (Fig. 1a). The intensity scanning data indicated that BMP-2 expression is downregulated in both human livers with inflammation hepatitis and cirrhosis (Fig. 1b). In contrast, the increased TGF-β1 expression was only noted in livers with inflammation hepatitis and cirrhosis (Fig. 1c). These results suggest that hepatic BMP-2 expression is downregulated in human fibrotic livers.

Downregulation of BMP-2 in human hepatitis and cirrhotic livers. a Immunohistochemistry (IHC) analysis of BMP-2 and TGF-β1 expression in human fibrotic liver tissue microarray (TMA). TMA slides containing normal, hepatitis, and cirrhosis livers were subjected to IHC staining with BMP-2 and TGF-β1. Arrows indicate hepatocytes; arrowheads indicate HSCs. b, c Quantification of BMP-2 and TGF-β1 expression levels from IHC staining results. Data represent the mean ± SEM. *P < 0.05 compared between the indicated groups
Downregulation of BMP-2 in murine fibrotic livers
Thereafter, we used the fibrotic animal models to validate the association of BMP-2 expression during liver fibrosis. It was found that the BMP-2 expression was significantly downregulated in the BDL- and CCl4-treated mouse livers at both mRNA (Fig. 2a) and protein levels (Fig. 2b). Consistently, IHC analysis also indicated that BMP-2 level was significantly decreased at 1 and 2 weeks in BDL group or at 2 and 4 weeks in CCl4 group (Fig. 2c). This was correlated with a significantly increased number of TGF-β1-positive cells in both models (Fig. 2d). Moreover, TGF-β1- and BMP-2-positive signals were found in the fibrotic areas with parenchymal and stellate cell morphology. These results suggest that a mechanism of mutual regulation might exist between BMP-2 and TGF-β1 during hepatic fibrogenesis.

Downregulation of BMP-2 in murine fibrotic livers induced by BDL surgery and CCl4 administration. a Gene expression of BMP-2 in normal and fibrotic mouse livers. BMP-2 mRNA levels were measured by RT-qPCR. b Protein expression of BMP-2 in mouse normal and fibrotic livers. Representative images of western blotting (upper) and densitometry data (lower) are shown. c, d Localization and quantification of hepatic BMP-2 and TGF-β1 expression in normal and fibrotic mouse livers. Representative images of immunohistochemical staining are shown (upper). Arrows indicate hepatocytes; arrowheads indicate HSCs. Signal mean intensities in the fibrotic areas were quantified based on 6–10 different fields/slide (lower). Data represent the mean ± SEM of four or five samples per group. *P < 0.05 compared between the indicated groups. Original magnification: × 200
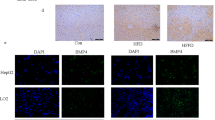
To delineate the types of cells with BMP-2 expression in the fibrotic livers, dual immunofluorescence staining showed remarkably increased expression of GFAP, which is known as an early HSC activation marker, in the BDL-treated mouse livers compared with normal control (Fig. 3a, b). The BMP-2 immunoreactivity was significantly decreased in mouse fibrotic livers (Fig. 3b). In addition, the colocalization of BMP-2 with GFAP was only seen in fibrotic liver at 7 days post BDL surgery, indicating that HSCs also expressed BMP-2 protein during fibrogenesis. By contrast, TGF-β1 immunoreactivity was significantly increased along with the HSC activation (Fig. 3c, d). Similar to the IHC-staining pattern (Fig. 2d), the immunofluorescence staining by using albumin as a hepatocyte marker further demonstrated the colocalization of BMP-2 with albumin proteins in both normal and BDL-treated livers (Supplementary Figure S1). Taken together, these results strongly suggest that the downregulated BMP-2 in mouse fibrotic livers post BDL treatment was localized in both parenchymal hepatocytes and activated HSCs.

Colocalization of BMP-2 or TGF-β1 with GFAP in BDL-induced mouse fibrotic livers. a, c Dual staining of BMP-2 or TGF-β1 with GFAP, an early marker of activated HSCs. Representative images stained with GFAP and BMP-2 or TGF-β1 demonstrated the colocalization of BMP-2 and TGF-β1 in the activated HSCs. b Quantification of fluorescence intensities of GFAP and BMP-2. d Quantification of fluorescence intensities of GFAP and TGF-β1. The intensity data were measured from 5–10 different fields per sample. Data represent the mean ± SEM. * P < 0.05 compared between the indicated groups. Original magnification: × 400
Prophylactic BMP-2 gene delivery suppressed hepatic injury and collagen deposition in experimental liver fibrosis
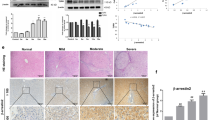
To evaluate the prophylactic effect of BMP-2 on hepatic fibrogenesis, mice were given adenoviral vectors carrying BMP-2 or GFP genes 1 day before BDL surgery or CCl4 injection, and sera and liver tissues were harvested after 2 weeks and 4 weeks, respectively. Serum biochemical measurements showed that BMP-2 gene delivery significantly reduced AST, ALT, and ALP enzyme activities in sera of BDL-treated mice (Fig. 4a), but only alleviated ALP levels in the mice with CCl4-mediated hepatic injury (Fig. 4b). Moreover, IHC staining with CK19, a cholangiocyte marker, demonstrated that BMP-2 gene delivery significantly ameliorated ductular reaction in the settings of both BDL- (Fig. 4c, d) and CCl4-induced biliary damage (Fig. 4e, f). Further histological examinations with H&E stain indicated that Ad-BMP-2 infection markedly decreased the number and magnitude of bile infarcts in livers of BDL-treated mice and alleviated the degree of hepatic fibrosis and pseudolobular formation in CCl4-treated mice (Fig. 5a). METAVIR scoring analysis indicated that BMP-2 gene delivery significantly reduced the degree of liver fibrosis compared with Ad-GFP groups (Fig. 5b). In addition, Sirius red staining was used to visualize collagen bundles distributed in fibrous bridging and nodular patterns in fibrotic mouse livers (Fig. 5c). Subsequent morphometrical analysis with METAVIR scoring analysis showed that BMP-2 gene delivery prominently reduced the increased areas of collagen deposition in both models (Fig. 5d). Although BMP-2 gene delivery did not affect serum TGF-β1 levels in BDL-treated mice (data not shown), IHC staining results indicated that BMP-2 gene delivery increased hepatic CD68-positive cells (Supplementary Figure S2) and decreased MMP-9 expression only in the mice with BDL treatment (Supplementary Figure S3). In addition, BMP-2 prominently reduced α-SMA expression in BDL model (Supplementary Figure S4) and suppressed fibronectin expression in CCl4 model (Supplementary Figure S5). These findings collectively support that BMP-2 gene therapy exerts prophylactic effect on liver fibrogenesis by alleviating liver injury and abating collagen deposition and fibrosis severity in the livers.

Effects of adenovirus-mediated BMP-2 gene delivery on BDL- and CCl4-induced mouse hepatic and biliary injuries. Mice received prophylactic treatment with adenovirus carrying BMP-2 (Ad-BMP-2) or GFP (Ad-GFP) as vehicle control at 1 day before undergoing BDL surgery or consecutive CCl4 administration. Sera and liver tissues were collected after 2 and 4 weeks, respectively. a Effects of BMP-2 gene delivery on BDL-induced liver injury. Serum biochemical measurements included ALT, AST, ALP, direct, and total bilirubin. b Effects of BMP-2 gene delivery on serum ALP levels in CCl4-treated mice. c, e IHC staining of cytokeratin 19 (CK19) in fibrotic livers with or without BMP-2 gene delivery in both models. Original magnification: × 200. d, f Quantification of CK19-positive areas, showing suppressive effect of BMP-2 gene therapy on biliary damage in both models. Data are expressed as mean ± SEM. *indicates a P < 0.05 between the indicated groups

Effects of adenovirus-mediated BMP-2 gene delivery on BDL- and CCl4-induced liver fibrosis of mice. Mice received prophylactic treatment with adenovirus carrying BMP-2 (Ad-BMP-2) or GFP (Ad-GFP) as vehicle control at 1 day before undergoing BDL surgery or consecutive CCl4 administration. Liver tissues were collected after 2 and 4 weeks, respectively. a H&E staining of mouse fibrotic livers with Ad-GFP or Ad-BMP-2 treatment. Arrows indicate hepatic infarct areas. b Metavir fibrosis in both models. c Sirius red (SR) staining of mouse fibrotic livers with Ad-GFP or Ad-BMP-2 treatment. Arrows indicate collagen fibril bundles. d Quantification of SR-positive area. Data represent the mean ± SEM of six samples per group. *P < 0.05 compared between the indicated groups
Mutual regulation of BMP-2 and TGF-β1 expression in cultured HSCs/hepatocytes
To determine the role of TGF-β1 in regulation of BMP-2 expression in HSCs/hepatocytes and vice versa, HSC-T6 and Clone-9 cells were treated with recombinant BMP-2 or TGF-β1 in vitro. Quantitative RT-PCR analysis showed that TGF-β1 significantly suppressed BMP-2 mRNA in HSC-T6 cells (Fig. 6a) and clone-9 hepatocytes (Fig. 6b). On the contrary, BMP-2 significantly suppressed TGF-β1 mRNA levels in HSC-T6 cells (Fig. 6a) and Clone-9 hepatocytes (Fig. 6b). Consistently, immunoblot analysis indicated that TGF-β1 significantly suppressed BMP-2 protein, whereas BMP-2 significantly suppressed TGF-β1 protein contents in HSC-T6 (Fig. 6c) and Clone-9 cells (Fig. 6d). These results suggest that a mechanism of mutual regulation exists between BMP-2 and TGF-β1 during hepatic fibrogenesis.

Mutual regulation between BMP-2 and TGF-β1 in cultured HSCs and hepatocyte cell lines. HSC-T6 or Clone-9 cells were treated with recombinant BMP-2 or TGF-β1 at indicated doses. After 6 and 24 h, total RNA and protein lysates, respectively, were extracted and subjected to RT-qPCR and western blotting detections. a, b RT-qPCR detection of BMP-2 and TGF-β1 mRNA levels in HSC-T6 cells and clone-9 hepatocytes. c, d Western blotting and densitometry of BMP-2 and TGF-β1 protein levels in HSC-T6 cells and clone-9 hepatocytes. β-actin was used as internal loading control. All data are expressed as mean ± SEM (n = 3). *P < 0.05 compared between the indicated groups
BMP-2 pretreatment mitigated TGF-β1-induced HSC proliferation and migration
Given that the cells undergoing the EMT process have higher mobility, we next examined the modulatory effects of BMP-2 on the cell proliferating and migrating activities in TGF-β1-stimulated HSCs. WST-1 cell proliferation assay demonstrated that BMP-2 pretreatment significantly inhibited the TGF-β1-induced cell proliferation (Fig. 7a). Wound-healing assay demonstrated that BMP-2 alone and pretreatment significantly blocked the TGF-β1-stimulated motility of primary mouse HSCs (Fig. 7b, c). These results demonstrated that BMP-2 may be a key mediator that negatively regulates the TGF-β1-altered expression of EMT markers involved in hepatic fibrogenesis.

Effects of BMP-2 on TGF-β1-stimulated proliferation and migration of primary mouse HSCs. Primary HSCs were pretreated with BMP-2 (250 ng/ml) or PBS (vehicle control) for 1 h, followed by TGF-β1 treatment (10 ng/ml) for 48 h. a Effect of BMP-2 pretreatment on TGF-β1-increased HSC proliferation. A WST-1 assay was performed after 48 h of TGF-β1- stimulation. b Effect of BMP-2 pretreatment on TGF-β1-increased HSC migration. The representative images (original magnification × 40) of a wound-healing migration assay were photographed at the indicated time points. c Image analysis data of migration assay at 15 h. All data are expressed as mean ± SEM (n = 4). *P < 0.05 compared between the indicated groups
BMP-2 inhibited TGF-β1-induced HSC activation, myofibroblastic transformation, and EMT marker expressions
To examine whether BMP-2 affects TGF-β1-induced HSC activation, myofibroblastic transformation, and ECM formation, primary mouse HSCs were isolated and pretreated with BMP-2 for 1 h followed by TGF-β1 stimulation for 48 h. Immunofluorescence staining with α-SMA and fibronectin antibodies revealed that, compared with the vehicle control, TGF-β1-increased α-SMA and fibronectin expression; BMP-2 alone had a slight effect on α-SMA and fibronectin expression. However, BMP-2 pretreatment significantly inhibited TGF-β1-induced α-SMA and fibronectin expression compared with TGF-β1 alone (Fig. 8a, b). In parallel, fluorescence visualization with EMT markers, E-cadherin and vimentin, showed that BMP-2 significantly reversed the TGF-β1-induced downregulation of E-cadherin as well as the upregulation of vimentin in cultured HSCs (Fig. 8c, d). These findings suggest that BMP-2 inhibits TGF-β1-induced HSC activation, myofibroblastic transformation, and EMT marker expression.

Effects of BMP-2 on TGF-β1-induced activation and EMT marker expression in primary mouse HSCs. Primary HSCs were pretreated with BMP-2 (250 ng/ml) or PBS (vehicle control) for 1 h, followed by TGF-β1 treatment (10 ng/ml) for 48 h. a Immunofluorescent staining of α-SMA (red), fibronectin (green), and nuclei (blue) in HSCs (original magnification × 200). b Fluorescence intensity analysis of α-SMA and fibronectin. c Immunofluorescent staining of E-cadherin (red), vimentin (green), and nuclei (blue) in HSCs (original magnification × 200). d Fluorescence intensity analysis of EMT marker expression, E-cadherin and vimentin. The fluorescence intensities were quantified by counting 5–10 different fields per sample. All data are expressed as mean ± SEM (n = 3). *P < 0.05 compared between the indicated groups
Suppressive effect of BMP-2 on TGF-β1/Smad signal axis in HSCs
To further elucidate the underlying mechanism by which BMP-2 antagonizes EMT and TGF-β1/Smad signaling activity in the progression of hepatic fibrosis, we next measured protein levels of TGF-β1 signaling-associated mediators and EMT markers in the BMP-2-pretreated HSCs. Western blot detection clearly showed that BMP-2 pretreatment significantly suppressed TGF-β1-induced TGF-β1 RI, RII, and total Smad3 expression, as well as Smad3 hyperphosphorylation (Fig. 9a, b). In consistence with the fluorescence staining results, BMP-2 significantly reversed the TGF-β1-triggered EMT process by increasing E-cadherin and decreasing fibronectin expression (Fig. 9c, d). These results demonstrated that BMP-2 significantly blocked the TGF-β1/Smad signal transduction, thereby inhibiting the TGF-β1-induced EMT response during liver fibrogenesis.

Blockade of TGF-β1-induced Smad signaling and HSC activation by BMP-2 in primary mouse HSCs. Primary HSCs were pretreated with BMP-2 (250 ng/ml) or PBS (vehicle control) for 1 h, followed by TGF-β1 treatment (10 ng/ml) for 48 h. The protein lysates of treated HSCs were subjected to western blotting detection. a, c Representative images of immunoblotting results. b Densitometrical quantification of TGF-β1 RI, RII, total, and phosphorylated Smad3 expression levels. d Densitometrical quantification of fibronectin and vimentin expression levels. GAPDH was used as an internal loading control. Data are expressed as mean ± SEM (n = 3). *P < 0.05 compared between the indicated groups. e Prophylactic mechanism of BMP-2 in liver fibrosis treatment. The present study demonstrates that concurrent BMP-2 downregulation and TGF-β1 upregulation were noted in human and mouse fibrotic livers. Mechanistically, BMP-2 may inhibit myofibroblastic activation of HSCs through counteracting TGF-β1/Smad signaling and suppress TGF-β1-induced EMT process via increasing E-cadherin and decreasing fibronectin and vimentin expression during hepatic fibrogenesis
Discussion
Despite that hepatic BMP expression has been previously reported in acute liver injury [33], the pathogenetic significance of BMP-2 in liver fibrosis remains a mystery. The current study investigated the role of BMP-2 and its prophylactic effect in hepatic fibrogenesis. The results delineated downregulation of BMP-2 in human hepatitis and cirrhosis TMA tissues and the mouse fibrotic livers from two models with different induction methods. Prophylactic experiment further confirmed that Ad-BMP-2 infection significantly attenuated BDL and CCl4-iduced liver injuries as revealed by reduction of plasma AST and ALT levels as well as hepatic CK19 contents. Histopathological staining analyses showed consistent results that BMP-2 gene delivery remarkably mitigated HSC activation and intrahepatic collagen deposition in both BDL- and CCl4-treated mouse livers. Furthermore, in vitro studies identified a mutual regulation mechanism between BMP-2 and TGF-β1 in cultured hepatocytes and HSC cell lines. Most importantly, BMP-2 was able to antagonize the TGF-β1-induced Smad signaling, cellular activation, ECM overproduction as well as EMT marker expression in primary mouse HSCs. The anti-fibrogenic role of BMP-2 in the livers is proposed and depicted in Fig. 9e.
This study is the first to report the downregulation of BMP-2 in human hepatitis and cirrhotic livers as well as in the fibrotic livers of mice receiving BDL or CCl4 treatment. It is worth to mention that a transient BMP-2 upregulation has been previously evidenced in the high-dose CCl4-induced acute liver injury in rats [33]. The upregulated BMP-2 was claimed to not only to participate in the process of hepatogenesis in early embryos, but also contribute to liver regeneration by eliciting wound-healing signal that governs maturation of hepatic stem-like oval cells. This notion could be supported by the fact that BMP-2 recombinant protein is frequently added into the culture medium for induction of hepatic differentiation and maturation in embryonic stem cells [34] and adult mesenchymal stem cells [35]. By contrast, BMP-2 down regulation is previously demonstrated in regenerative livers of the rats underwent partial hepatectomy [28], and BMP-2 is thus suggested a negative regulator in hepatogenesis, because it suppresses proliferation of hepatoma cells. It is conceivable that the major function of BMP-2 is to induce maturation, but not proliferation, of hepatic progenitor cells. It is therefore reasonable to propose that the downregulation of BMP-2 in injured livers may provide a regenerative niche for tissue repair. In the aspect of liver fibrogenesis, the BMP-2-suppressed motility of HSCs suggests that the reduced BMP-2 contents in fibrotic and regenerative livers may facilitate the mobilization HSCs and/or myofibroblasts to the site of injury.
In the mechanistic context of BMP-2 and TGF-β1 expression in livers, HSCs activation and trans-differentiation are important features in liver fibrosis and TGF-β1 has a pivotal role in fibrogenesis because of its potency on myofibroblast transition and ECM overproduction [30]. Given that BMPs and TGF-β belong to the same superfamily shared with a similar ligand structure and downstream signaling cascades, they have been known to play important roles in the regulation of many cellular functions [26]. Previous studies have demonstrated that BMP-2 can antagonize TGF-β by suppressing TGF-β1 receptors and Smad2/3 phosphorylation in renal interstitial fibrosis [23, 24, 26]. Consistent to those findings, this study identified a mutual regulation between BMP-2 and TGF-β1 in primary mouse HSCs and cultured hepatocyte cell line. Histological observation of hepatic BMP-2 and TGF-β1 in human and mouse fibrotic livers revealed both cytokines were expressed in parenchymal hepatocytes and HSCs, strongly suggesting that the equilibrium of both cytokines in pro-inflammatory and profibrogenic milieu might orchestrate HSC activation in the injured livers. Mechanistically, exogenous BMP-2 pretreatment significantly reduced the TGF-β1-induced increases in TGF-β1 receptor expression and Smad3 hyperphosphorylation. Interestingly, we noted that BMP-2 gene delivery markedly decreased hepatic MMP-9 expression in BDL-treated mice. In addition to serving as the key enzyme responsible for ECM degradation, MMP-9 has been known to activate latent TGF-β1 peptides [36, 37] and induce HSC trans-differentiation [38]. Other line of evidence also shows that BMP-2 suppress MMP-9 activation during chondrocyte commitment [39]. Taken together, these findings support that the BMP-2 gene delivery might attenuate liver fibrosis at least in part via the modulation of MMP-9 and TGF-β activities and the disruption of the TGF-β1/Smad3-signaling pathway.
The occurrence of EMT in liver fibrosis is now proved to be a viable therapeutic target, which is perhaps the most intriguing and controversial of recent hypotheses on the mechanism of fibrosis that contribute directly to matrix deposition and repair [8, 24, 26]. Also, TGF-β1, the classical EMT inducer will adopt a fibroblast-like morphology, with decreased membrane E-cadherin and increased expression of vimentin and fibronectin [8, 24]. The present study detected the EMT parameters in primary mouse HSCs and demonstrated that exogenous BMP-2 significantly suppressed the TGF-β1-induced fibronectin and vimentin expression in vitro. In contrast, the E-cadherin expression was increased after BMP-2 cotreatment. Moreover, BMP-2 significantly attenuated the activated HSC migration and proliferation. These results are consistent with previous mechanistic studies using cultured rat HSCs [40] and mouse pancreatic stellate cells [41]. The former study identified that the spontaneous activation of primary rat HSCs grown on plastic culture ware is accompanied with altered expression of EMT markers [40], the latter corroborated the suppressive effect of BMP-2 pretreatment on the TGF-β1-modulated EMT marker expression in mouse pancreatic stellate cells [41]. The findings of this study demonstrated that exogenous BMP-2 treatment antagonized the TGF-β1-induced Smad signaling activity, extracellular matrix biosynthesis, and myofibroblastic transformation of HSCs, supporting that BMP-2 may exhibit therapeutic effects via enhancing liver fibrosis resolution.
In conclusion, the current study demonstrated that the inhibitory effects of BMP-2 on hepatic fibrosis were via inhibiting the TGF-β1/Smad3 pathways, HSCs activation, as well as EMT suppression. All together, these results support that BMP-2 plays a negative role in regulation of TGF-β1 receptor expression, signal transduction, and subsequent HSC activation. It is suggested that BMP-2 might have both preventive and therapeutic effects on hepatic fibrogenesis. BMP-2 restoration might constitute a novel approach for treatment of liver fibrosis.
References
Lee UE, Friedman SL. Mechanisms of hepatic fibrogenesis. Best Pract Res Clin Gastroenterol. 2011;25:195–206.
Friedman SL. Evolving challenges in hepatic fibrosis. Nat Rev Gastroenterol Hepatol. 2010;7:425–36.
Bataller R, Brenner DA. Liver fibrosis. J Clin Invest. 2005;115:209–18.
Modol T, Brice N, Ruiz de Galarreta M, et al. Fibronectin peptides as potential regulators of hepatic fibrosis through apoptosis of hepatic stellate cells. J Cell Physiol. 2015;230:546–53.
Liu XY, Liu RX, Hou F, et al. Fibronectin expression is critical for liver fibrogenesis in vivo and in vitro. Mol Med Rep. 2016;14:3669–75.
Bissell DM, Roulot D, George J. Transforming growth factor beta and the liver. Hepatology. 2001;34:859–67.
Jonsson JR, Clouston AD, Ando Y, et al. Angiotensin-converting enzyme inhibition attenuates the progression of rat hepatic fibrosis. Gastroenterology. 2001;121:148–55.
O’Connor JW, Gomez EW. Biomechanics of TGFbeta-induced epithelial-mesenchymal transition: implications for fibrosis and cancer. Clin Transl Med. 2014;3:23.
Bottinger EP. TGF-beta in renal injury and disease. Semin Nephrol. 2007;27:309–20.
Liu X, Hu H, Yin JQ. Therapeutic strategies against TGF-beta signaling pathway in hepatic fibrosis. Liver Int. 2006;26:8–22.
Zeisberg M, Hanai J, Sugimoto H, et al. BMP-7 counteracts TGF-beta1-induced epithelial-to-mesenchymal transition and reverses chronic renal injury. Nat Med. 2003;9:964–8.
Park J, Schwarzbauer JE. Mammary epithelial cell interactions with fibronectin stimulate epithelial-mesenchymal transition. Oncogene. 2014;33:1649–57.
Border WA, Noble NA. Transforming growth factor beta in tissue fibrosis. N Engl J Med. 1994;331:1286–92.
Wozney JM, Rosen V, Celeste AJ, et al. Novel regulators of bone formation: molecular clones and activities. Science. 1988;242:1528–34.
Massague J. The transforming growth factor-beta family. Annu Rev Cell Biol. 1990;6:597–641.
Itoh S, Itoh F, Goumans MJ, et al. Signaling of transforming growth factor-beta family members through Smad proteins. Eur J Biochem. 2000;267:6954–67.
Feng XH, Derynck R. Specificity and versatility in TGF-beta signaling through Smads. Annu Rev Cell Dev Biol. 2005;21:659–93.
Eickelberg O, Morty RE. Transforming growth factor beta/bone morphogenic protein signaling in pulmonary arterial hypertension: remodeling revisited. Trends Cardiovasc Med. 2007;17:263–9.
Song JJ, Celeste AJ, Kong FM, et al. Bone morphogenetic protein-9 binds to liver cells and stimulates proliferation. Endocrinology. 1995;136:4293–7.
Duncan SA, Watt AJ. BMPs on the road to hepatogenesis. Genes Dev. 2001;15:1879–84.
Tsai MS, Suksaweang S, Jiang TX, et al. Proper BMP signaling levels are essential for 3D assembly of hepatic cords from hepatoblasts and mesenchymal cells. Dig Dis Sci. 2015;60:3669–80.
Xu CP, Ji WM, van den Brink GR, et al. Bone morphogenetic protein-2 is a negative regulator of hepatocyte proliferation downregulated in the regenerating liver. World J Gastroenterol. 2006;12:7621–5.
Yang YL, Liu YS, Chuang LY, et al. Bone morphogenetic protein-2 antagonizes renal interstitial fibrosis by promoting catabolism of type I transforming growth factor-beta receptors. Endocrinology. 2009;150:727–40.
Yang YL, Ju HZ, Liu SF, et al. BMP-2 suppresses renal interstitial fibrosis by regulating epithelial-mesenchymal transition. J Cell Biochem. 2011;112:2558–65.
Shlyonsky V, Soussia IB, Naeije R, et al. Opposing effects of bone morphogenetic protein-2 and endothelin-1 on lung fibroblast chloride currents. Am J Respir Cell Mol Biol. 2011;45:1154–60.
Gao X, Cao Y, Staloch DA, et al. Bone morphogenetic protein signaling protects against cerulein-induced pancreatic fibrosis. PLoS ONE. 2014;9:e89114.
Kao YH, Chen CL, Jawan B, et al. Upregulation of hepatoma-derived growth factor is involved in murine hepatic fibrogenesis. J Hepatol. 2010;52:96–105.
Massoner P, Kugler KG, Unterberger K, et al. Characterization of transcriptional changes in ERG rearrangement-positive prostate cancer identifies the regulation of metabolic sensors such as neuropeptide Y. PLoS ONE. 2013;8:e55207.
Tai MH, Cheng H, Wu JP, et al. Gene transfer of glial cell line-derived neurotrophic factor promotes functional recovery following spinal cord contusion. Exp Neurol. 2003;183:508–15.
Huang YH, Tiao MM, Huang LT, et al. Activation of miR-29a in activated hepatic stellate cells modulates its profibrogenic phenotype through inhibition of histone deacetylases 4. PLoS ONE. 2015;10:e0136453.
Kao YH, Chen PH, Wu TY, et al. Lipopolysaccharides induce Smad2 phosphorylation through PI3K/Akt and MAPK cascades in HSC-T6 hepatic stellate cells. Life Sci. 2017;184:37–46.
Yang YL, Wang FS, Li SC, et al. MicroRNA-29a alleviates bile duct ligation exacerbation of hepatic fibrosis in mice through epigenetic control of methyltransferases. Int J Mol Sci. 2017;18:192.
Nakatsuka R, Taniguchi M, Hirata M, et al. Transient expression of bone morphogenic protein-2 in acute liver injury by carbon tetrachloride. J Biochem. 2007;141:113–9.
Cai J, Zhao Y, Liu Y, et al. Directed differentiation of human embryonic stem cells into functional hepatic cells. Hepatology. 2007;45:1229–39.
Li X, Yuan J, Li W, et al. Direct differentiation of homogeneous human adipose stem cells into functional hepatocytes by mimicking liver embryogenesis. J Cell Physiol. 2014;229:801–12.
Yu Q, Stamenkovic I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-beta and promotes tumor invasion and angiogenesis. Genes Dev. 2000;14:163–76.
Kao YH, Jawan B, Goto S, et al. Serum factors potentiate hypoxia-induced hepatotoxicity in vitro through increasing transforming growth factor-beta1 activation and release. Cytokine. 2009;47:11–22.
Han YP, Yan C, Zhou L, et al. A matrix metalloproteinase-9 activation cascade by hepatic stellate cells in trans-differentiation in the three-dimensional extracellular matrix. J Biol Chem. 2007;282:12928–39.
Choi YA, Kang SS, Jin EJ. BMP-2 treatment of C3H10T1/2 mesenchymal cells blocks MMP-9 activity during chondrocyte commitment. Cell Biol Int. 2009;33:887–92.
Zhang QD, Xu MY, Cai XB, et al. Myofibroblastic transformation of rat hepatic stellate cells: the role of Notch signaling and epithelial-mesenchymal transition regulation. Eur Rev Med Pharmacol Sci. 2015;19:4130–8.
Gao X, Cao Y, Yang W, et al. BMP-2 inhibits TGF-beta-induced pancreatic stellate cell activation and extracellular matrix formation. Am J Physiol Gastrointest Liver Physiol. 2013;304:G804–813.
Acknowledgements
This study was supported by the grants in part from Kaohsiung Chang Gung Memorial Hospital (Nos. CMRPG8F1561, CMRPG8F1562) and Kaohsiung Armed Forces General Hospital (No. 103-6), Taiwan.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
These authors contributed equally: Yueh-Hua Chung, Ying-Hsien Huang.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Chung, YH., Huang, YH., Chu, TH. et al. BMP-2 restoration aids in recovery from liver fibrosis by attenuating TGF-β1 signaling. Lab Invest 98, 999–1013 (2018). https://doi.org/10.1038/s41374-018-0069-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41374-018-0069-9
This article is cited by
-
Growth differentiation factor 7 autocrine signaling promotes hepatic progenitor cell expansion in liver fibrosis
Stem Cell Research & Therapy (2023)
-
Dynamic alterations in metabolomics and transcriptomics associated with intestinal fibrosis in a 2,4,6-trinitrobenzene sulfonic acid-induced murine model
Journal of Translational Medicine (2023)
-
MiR-26a-5p as a useful therapeutic target for upper tract urothelial carcinoma by regulating WNT5A/β-catenin signaling
Scientific Reports (2022)
-
Bone morphogenetic protein 2 is a new molecular target linked to non-alcoholic fatty liver disease with potential value as non-invasive screening tool
Biomarker Research (2022)
