
Abstract
Histone acetylation and other modifications of the chromatin are important regulators of gene expression and, consequently, may contribute to drug-induced behaviors and neuroplasticity. Earlier studies have shown that a reduction in histone deacetylase (HDAC) activity results in the enhancement of some psychostimulant-induced behaviors. In this study, we extend those seminal findings by showing that the administration of the HDAC inhibitor sodium butyrate enhances morphine-induced locomotor sensitization and conditioned place preference. In contrast, this compound has no effects on the development of morphine tolerance and dependence. Similar effects were observed for cocaine and ethanol-induced behaviors. These behavioral changes were accompanied by a selective boosting of a component of the transcriptional program activated by chronic morphine administration that included circadian clock genes and other genes relevant to addictive behavior. Our results support a specific function for histone acetylation and the epigenetic modulation of transcription at a reduced number of biologically relevant loci on non-homeostatic, long-lasting, drug-induced behavioral plasticity.
Similar content being viewed by others


INTRODUCTION
Behavioral changes observed after chronic exposure to drugs of abuse, such as tolerance, dependence, and addictive behavior, appear or are maintained long after the drugs have been cleared from the organism and cannot be accounted by acute effects of the interaction of drugs with their primary molecular targets. Although these clinically relevant phenomena notably differ in their temporal persistence, all of them arise from the ability of drugs to promote persistent structural and functional changes in the central nervous system (Chao and Nestler, 2004). These phenomena are usually referred as ‘drug-induced neuroplasticity’ and depend on changes in gene expression (McClung and Nestler, 2008).
There is a growing interest in the possible functional consequences of covalent modifications of the chromatin in the appearance and maintenance of behavioral changes (Sweatt, 2009), including the development and manifestation of addictive behavior (Renthal and Nestler, 2008b; Tsankova et al, 2007). Changes in the structure of the chromatin could underlay long-lasting changes on neuronal gene expression and ultimately contribute to explain the persistence of addictive behavior. Recent studies have provided initial support to this hypothesis. For example, the administration of psychostimulants, such as cocaine, at dosages that promote conditioned place preference (CPP) or locomotor sensitization results in histone hyperacetylation at specific loci relevant to the development of addictive behavior (Kumar et al, 2005; Levine et al, 2005; Renthal et al, 2008a, 2007) and perhaps also at the bulk chromatin level (Kalda et al, 2007). Furthermore, genetic studies have established a functional role for histone acetyltransferase (HAT) (Levine et al, 2005) and histone deacetylase (HDAC) (Renthal et al, 2007) activities in the mechanisms of action of psychostimulants, a view also supported by pharmacological experiments with HDAC inhibitors (HDACi) (Kalda et al, 2007; Kumar et al, 2005; Renthal et al, 2007).
As most of the research conducted in this area has focused on psychostimulants such as cocaine or amphetamine, here we explore the effect of HDAC inhibition on drugs of abuse belonging to three different pharmacological families: cocaine, ethanol, and morphine. We found that the HDACi sodium butyrate selectively enhanced some, but not all, behavioral responses to chronic administration of these drugs. We also extended earlier studies by performing a detailed biochemical and gene profiling analysis of striatal tissue, which showed that morphine, as cocaine, induces striatal histone H3 phosphorylation, and indicated that the effects on behavior are not associated to global changes in gene expression or chromatin acetylation, but to the specific modulation of relevant loci and genetic programs.
METHODS AND MATERIALS
Subjects
Male Swiss-Albino mice, 6–8 weeks old, were purchased from Janvier España, S.A. (Madrid, Spain). Subjects housing, care, and experimental manipulation followed the national guidelines and approved by the Institutional Animal Care and Use Committees.
Behavioral Procedures
Sensitization
Locomotor sensitization induced by ethanol, cocaine, and morphine was evaluated using a protocol divided into two phases: induction and challenge. The induction phase involved six trials on alternate days, one trial per day. On each one of these trials, mice received an injection of saline or sodium butyrate (100, 150, or 300 mg/kg) followed by a second injection of ethanol (2.5 g/kg), morphine (20 mg/kg), or cocaine (10 mg/kg). On the basis of the results of earlier studies (Kumar et al, 2005), both treatments were separated by a 20 min delay in the case of ethanol or cocaine experiments. However, attending to the need of using a longer period in morphine-related experiments (see below), sodium butyrate was simultaneously administered with morphine. Immediately after this second injection, mice were placed in open-field chambers, consisting of glass cylinders of 25 cm in diameter, and locomotion was registered by a computerized video-tracking system (SMART, Panlab SL, Spain). The duration of these testing sessions was restricted to 20 min in the case of ethanol and cocaine, whereas it was prolonged to 60 min in the case of morphine. On the other hand, the challenge phase consisted of a single trial conducted 7 days after the last test of the induction phase. In this case, all animals received a single injection of ethanol (2.5 g/kg), cocaine (10 mg/kg), or morphine (20 mg/kg), and locomotion was assessed as in the treatment phase.
Conditioned place preference
Morphine-induced CPP was assessed using four black acrylic chambers (30 × 15 × 20 cm). Tactile cues (interchangeable grid and hole floors) were used as conditioned stimuli. The behavioral procedure was divided into three consecutive phases. First, initial preference was assessed in three successive daily tests by placing each animal in the CPP chambers (floor divided with half grid, half holes) for 15 min. The individual scores of the third test were used to match two groups that did not differ according to their initial preference for any of the floors. The second phase of this procedure consisted of six trials (20 min duration, one trial per day), corresponding to three morphine/CS+ pairings and three saline/CS− pairings. In each one of the six conditioning sessions, mice received an injection of saline or sodium butyrate (150 mg/kg) 20 min before receiving the corresponding injection of morphine (20 mg/kg) or saline. Immediately after this second injection, mice were confined into the conditioning chamber prepared with the corresponding CS+ or CS− floor for 20 min. The third phase consisted of a single test (duration: 15 min) conducted on the day after the last conditioning session. In this case the floor of the conditioning chambers was divided (half grid, half holes) and the time spent in each floor was assessed by a video-tracking system (SMART, Panlab SL, Spain).
Tolerance
The development of tolerance to the analgesic effects of morphine was assessed using the tail-flick procedure using an automated analgesiometer (Ugo Basile, Italy). The procedure had three main phases. First, drug-naive mice were first tested to establish their individual pain thresholds in a single assay (infrared intensity: 50). According to these initial values, mice were matched in two pretreatment (saline vs sodium butyrate) groups. In the second phase, that started 24 h later, mice received an acute saline or sodium butyrate (300 mg/kg, i.p.) injection and, 20 min later, all mice received a saline injection. The latency to withdraw the tail from the heat focus was evaluated in a single assay performed 10 min after the saline injection. Finally, the third phase of this experiment addressed the analgesic effects of morphine and the development of tolerance to this effect. This phase consisted of six trials (one trial per day on consecutive days). Each trial was identical to that described for phase 2, but mice were treated with morphine (5 mg/kg, i.p.) instead of saline 10 min before assessing the latency to flick the tail. The development of tolerance to the motor impairing effects of ethanol (2.5 g/kg) was assessed using a commercial rota-rod for mice (rota-rod 9756; Ugo Basile, Italy). First, ethanol-naive mice were trained in the rota-rod under a constant acceleration schedule (3.75–37.5 rpm over 5 min) till they were able to stay in the rod for 120 s for two of three consecutive trials. All mice learned this ability after two training sessions (five trials per session) conducted in consecutive days. Twenty-four hours after this training phase, mice were randomly assigned to a pretreatment group and received an acute saline or sodium butyrate (300 mg/kg) injection. Twenty minutes after this treatment, mice received a saline injection, and 10 min later, they were tested once in the rota-rod under the same schedule used in the training phase. This test was conducted to assess a possible effect of sodium butyrate in motor coordination per se. The third phase of this experiment addressed the motor impairing effects of ethanol and the development of tolerance to this effect and consisted of six trials (one trial per day on consecutive days). These trials were identical to that described earlier, but ethanol (2.5 g/kg), instead of saline, was administered to saline or sodium butyrate-pretreated mice.
Dependence/withdrawal
Opioid dependence was induced by repeated injections of morphine based on the procedure described by Maldonado et al (1997). Mice received morphine injections twice a day (9 AM and 9 PM) with progressively increasing doses (20, 40, 60, 80, and 100 mg/kg, i.p.) in their home cages for 5 consecutive days. Saline or sodium butyrate (150 mg/kg) was intraperitoneally injected immediately before each morphine injection. On the 6th day, mice were pretreated with saline or sodium butyrate followed by a last morphine (100 mg/kg, i.p., 9 AM) injection. After 90 min, mice were individually placed in cubic metacrylate boxes (40 × 40 × 40 cm). Fifteen minutes later, morphine withdrawal was precipitated by a subcutaneous naloxone injection (1 mg/kg) and mice behavior was videotaped for 15 additional minutes. Several signs of the morphine withdrawal syndrome were evaluated by an observer blind to treatment conditions. Ethanol dependence was induced by repeated injection of ethanol according to a self-developed experimental protocol adapted from Gililland and Finn (2007) using the reduction of locomotion as an index of ethanol withdrawal intensity (Kliethermes, 2005). Our experimental procedure had two main phases, namely, dependence induction and withdrawal assessment. In the first one, four separate groups of mice received an injection of saline or sodium butyrate (150 mg/kg, i.p.) in their home cages immediately followed by an ethanol (4 g/kg, i.p.) or saline challenge. This treatment was repeated twice a day (9 AM and 9 PM) for 5 consecutive days. The second phase consisted of a single test session conducted 15 h after the last saline/ethanol injection. In this test, mice locomotion was assessed during 20 min in an open field and following the same experimental conditions described above.
Western Blotting and Immunohistochemistry
Mice striatal tissue was rapidly dissected from anterior 6 mm coronal sections using a chilled acrylic mouse brain slicer matrix (Zivic Instruments) with 2 mm coronal section slice intervals, frozen in dry ice and stored at −70°C until further protein extraction. Sample protein concentration was determined and equal amounts of protein were loaded in each lane. Western blot analyses were carried out using Western Lightning ECL kit (Perkin-Elmer, Boston, MA). Immunohistochemistry analyses were performed as described earlier (Lopez de Armentia et al, 2007). See Supplementary Methods for additional details.
Microarray and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT–PCR) Analyses
For microarray analysis, striatal tissue was dissected as described above and placed on RNAlater solution (Qiagen, Venlo, The Netherlands). Equal amounts of total RNA from four animals were pooled, processed, and hybridized to Mouse Gene 1.0 ST genechips (Affymetrix, Santa Clara, CA). Three to six biological replicates were prepared for each experimental condition (saline–saline, N=6; saline–morphine, N=3; butyrate–morphine, N=3). Microarray data were processed, normalized, and statistically analyzed using GeneSpring GX. This dataset is accessible at the GEO database (accession number GSE17731). For qRT–PCR, cDNA was prepared from eight independent mice per group. Real-time quantitative PCR was performed using ABI 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA) and amplified using SYBR GreenER qPCR reagent mix (Invitrogen, Carlsbad, CA). Each sample was assayed in duplicate and normalized using GAPDH levels. Primer sequences for qRT–PCR amplification were designed using Primer Express Software v2.0 (Applied Biosystems) and are available on request.
Statistics
All data are depicted as mean±SEM. Most data were analyzed using ANOVAs, followed by Tuckey HSD post hoc tests because it does not require a significant interaction between factors and it is highly conservative against type I error. Experimental designs involving the comparison of multiple dependent variables were analyzed using between-groups MANOVA. Comparisons involving only two means were compared by means of Student's t-test for independent samples. For clarity, details of statistical comparisons that did not yield significant (p<0.05) differences are not fully described in the results section.
RESULTS
Sodium Butyrate Administration Enhances Cocaine-Induced Locomotor Sensitization
To confirm and extend the earlier findings on the ability of HDACi to enhance psychostimulants-induced behaviors, we assessed the effects of sodium butyrate on cocaine-induced locomotor sensitization. Our experimental protocol was divided in two phases: induction and challenge (see Figure 1a for details). Mice were divided into eight groups and injected with either saline or cocaine (10 mg/kg, i.p.) in the presence or absence of sodium butyrate pretreatment (100, 150, or 300 mg/kg, i.p.). The administration of sodium butyrate enhanced the development of cocaine-induced locomotor sensitization without affecting locomotion in saline-treated mice (Figure 1b). A three-way ANOVA (pretreatment × treatment × days) revealed that the (cocaine) treatment as well as the days factors reached statistical significance (F1,85=144.08, p<0.001 and F5,425=9.16, p<0.001), whereas the pretreatment (sodium butyrate) factor did not. Further, all bifactorial (F3,85=3.37, p<0.05; F5,425=10.73, p<0.001; and F15,425=2.56, p<0.001) as well as the three-way interaction (F15,425=1.99, p<0.05) yielded a significant effect. Tuckey HSD post hoc comparisons revealed that repeated cocaine injections in saline-pretreated mice led to a progressive enhancement of locomotion, confirming the development of locomotor sensitization (p<0.05). This behavior was boosted in mice pretreated with sodium butyrate. Notably, the effect of sodium butyrate was highly persistent. A challenge test conducted 7 days later, in which the same eight groups of mice received cocaine (10 mg/kg, i.p.), but not sodium butyrate, revealed that the mice that had received moderate (150 mg/kg) or high (300 mg/kg) doses of sodium butyrate concurrently with cocaine during the treatment phase exhibited higher locomotion in the challenge test than those that were pretreated with saline (Figure 1c). Two-way ANOVA (pretreatment × treatment) comparing the locomotor scores in this test revealed a significant effect for both factors (F3,85=3.57, p<0.05 and F1,85=56.84, p<0.001, respectively) as well as for their interaction (F3,85=3.63, p<0.05), whereas post hoc comparisons showed significant increases for the two highest sodium butyrate concentrations (p<0.01). As expected, regardless of their pretreatment conditions, all the mice that did not receive cocaine during the sensitization induction phase exhibited enhanced locomotion in the challenge phase in response to acute cocaine administration (Figure 1c, compare white bars with dashed line).

Effects of sodium butyrate administration on cocaine-induced locomotor sensitization. (a) Scheme of the behavioral training and treatment groups used for the sensitization experiment. The development and expression of locomotor sensitization was evaluated using a protocol divided into two phases: induction and challenge. (b) Sodium butyrate enhanced the ability of repeated cocaine (10 mg/kg) injections to promote locomotor sensitization, but it did not alter locomotion in saline-treated mice. (c) The effects of sodium butyrate were persistent and those mice receiving this HDACi as coadjuvant during the induction phase exhibited higher locomotion when re-exposed to cocaine in the challenge phase 7 days later. B100, B150, and B300 denotes pretreatment with 100, 150, or 300 mg/kg of sodium butyrate, respectively. The horizontal line of panel 1c depicts average locomotion of saline-treated groups at the induction phase. In all panels +p<0.05, *p<0.01 as compared with SC group.
Sodium Butyrate Administration Enhances Ethanol-Induced Locomotor Sensitization, but not Ethanol Tolerance or Withdrawal
We next investigated the effects of sodium butyrate in ethanol-induced behaviors. First, we evaluated the effects of this HDACi on ethanol (2.5 g/kg, i.p.)-induced sensitization using the same experimental scheme described earlier for cocaine (Figure 2a). As depicted in Figure 2b, we observed that sodium butyrate enhanced ethanol-induced locomotor sensitization. A three-way ANOVA (pretreatment × treatment × days) revealed a significant effect of the three main factors (F3,106=4.39, p<0.001; F1,106=162.98, p<0.001; and F5,503=19.16, p<0.001, respectively). Two bifactorial interactions reached statistical significance (pretreatment × treatment: F3,106=6.21, p<0.001 and treatment × days: F5,530=29.43, p<0.001). Post hoc comparisons revealed that all doses of sodium butyrate significantly enhanced ethanol-induced locomotion as compared with the saline-pretreated group (100 mg/kg: p<0.05, 150 mg/kg: p<0.001; 300 mg/kg: p<0.001, respectively). The same sodium butyrate doses did not affect the daily scores of saline-treated mice (Figure 2b). Again, these differences between groups were persistent and were expressed in a challenge test conducted 7 days later in which all mice received a single ethanol injection (Figure 2c). A two-way ANOVA (pretreatment × treatment) comparing the locomotor scores on this challenge test revealed that both main factors as well as their interaction reached statistical significance (F3,92=3.51, p<0.05; F1,92=86.17, p<0.001; and F3,92=5.60, p<0.01; respectively). Thus, mice that had received sodium butyrate and ethanol during the sensitization induction phase exhibited higher locomotion than the group that received saline as a coadjuvant treatment of ethanol (p<0.01 in all cases). All mice that had been treated with saline during the induction phase equally reacted to an acute ethanol administration.

Effects of sodium butyrate administration on ethanol-induced behaviors. (a) Scheme of the behavioral training and treatment groups used for the sensitization experiment. The development and expression of locomotor sensitization was evaluated using a protocol divided into two phases: induction and challenge. (b) Co-administration of sodium butyrate and ethanol (2.5 g/kg) increased the development of locomotor sensitization (in all panels of this figure: +p<0.05, *p<0.01 as compared with SE group). (c) Mice that had received sodium butyrate and ethanol during the induction phase exhibited higher locomotion when re-challenged 7 days later with ethanol (2.5 g/kg) than those that had been only exposed to ethanol. B100, B150, and B300 denotes pretreatment with 100, 150, or 300 mg/kg of sodium butyrate, respectively. The horizontal line depicts average locomotion of saline-treated groups at the induction phase. (d) The administration of sodium butyrate (300 mg/kg) did not modify the motor-incoordinating effects of ethanol (2.5 g/kg) or the development of tolerance as this treatment was repeated. (e) Fifteen hours after the termination of an intense ethanol administration regimen (4 g/kg, twice daily; 5 days) a significant reduction on spontaneous locomotion was observed (p<0.05). The magnitude of this ethanol withdrawal sign was not modified in mice that had received sodium butyrate (150 mg/kg) with each ethanol injection.
In addition to locomotor sensitization, the repeated administration of ethanol can cause other relevant behavioral adaptations, such as tolerance and withdrawal symptoms. We extended our study to other behavioral effects of ethanol and found that sodium butyrate co-administration did not affect the development of tolerance to the motor-incoordinating effects of ethanol (Figure 2d). Similarly, we also observed that the co-administration of this HDACi during the induction of alcohol dependence did not affect the hypolocomotion associated with ethanol withdrawal (Figure 2e). These effects were not caused by changes in ethanol pharmacokinetics (Supplementary Table 1).
Sodium Butyrate Administration Enhances Morphine-Induced Sensitization and CPP, but not Tolerance or Withdrawal
To extend and confirm this intriguing dissociation on the effects of HDACis on different drug-induced behavioral phenomena, we assessed the effect of sodium butyrate on several behavioral effects of chronic morphine administration. First, we evaluated the effects of this HDACi on morphine-induced sensitization using the same protocol described for cocaine and ethanol (Figure 3a). A three-way ANOVA (pretreatment × treatment × days) revealed a significant effect of the three main factors (F3,87=12.12, p<0.001; F1,87=36.66, p<0.001; and F5,435=41.01, p<0.001, respectively). The dyadic interactions treatment × days (F5,435=47.22, p<0.001) and pretreatment × treatment (F3,87=10.99, p<0.001) reached statistical significance. Tuckey HSD post hoc comparisons revealed that all mice treated with morphine displayed a significant increase in locomotion across days, and this effect was significantly boosted in mice pretreated with moderate (150 mg/kg, p<0.05) or high doses of sodium butyrate (300 mg/kg, p<0.01). The same doses of sodium butyrate did not affect locomotion of saline-treated mice. As observed for cocaine- and ethanol-treated mice, the co-administration of sodium butyrate (150 or 300 mg/kg) selectively enhanced the development of morphine-induced locomotor sensitization (Figure 3b). Notably, as observed for cocaine- and ethanol-treated mice, the effects of sodium butyrate persisted and were still apparent in a challenge test conducted 7 days after the last sodium butyrate injection. As in earlier experiments, in the challenge phase, all mice received morphine, but not sodium butyrate. We found that those mice that had concurrently received intermediate or high doses of sodium butyrate and morphine during the induction phase exhibited higher locomotion than the group that had received saline as the coadjuvant of morphine (Figure 3c; p<0.01).

Effects of sodium butyrate administration on morphine-induced behaviors. (a) Scheme of the behavioral training and treatment groups used for the sensitization experiment. The development and expression of locomotor sensitization was evaluated using a protocol divided into two phases: induction and challenge. (b) The development of morphine-induced locomotor sensitization was enhanced in mice treated with intermediate (150 mg/kg) or high (300 mg/kg) doses of sodium butyrate. (c) Expression of locomotor sensitization in the same eight groups of mice 7 days after the termination of the induction phase. All mice were challenged with a single morphine (20 mg/kg) injection. The horizontal line depicts the averaged locomotion score of all groups receiving saline injections during the last test of the induction phase. The boosting effect of sodium butyrate still persisted when the mice were re-exposed to morphine 7 days after the last injection. B100, B150, and B300 denotes pretreatment with 100, 150, or 300 mg/kg of sodium butyrate, respectively. (d) The analysis of locomotor sensitization in four additional groups of mice receiving sodium butyrate (0 or 300 mg/kg) and morphine (10 or 20 mg/kg) revealed that repeated administration of the lower dose of morphine did not result in locomotor sensitization regardless of the coadjuvant (saline or sodium butyrate) treatment. In contrast, repeated administration of a higher dose of morphine resulted in the development of locomotor sensitization, an effect that was boosted by the administration of sodium butyrate. These data were further analyzed by a linear-regression-based procedure (straight lines on the figure; see results section for further details). (e) Sodium butyrate (150 mg/kg) increases morphine-induced CPP. (f) Sodium butyrate (300 mg/kg) does not affect the development of tolerance to the analgesic effects of morphine (5 mg/kg, tail-flick test). (g) Effects of sodium butyrate (150 mg/kg) on naloxone-precipitated morphine withdrawal. For all panels of this figure: +p<0.05 and *p<0.01 as compared with SM group.
We also compared the interaction between a sodium butyrate dose (300 mg/kg) and two doses of morphine (10 and 20 mg/kg) that differed in their ability to promote locomotor sensitization under the treatment conditions described above (Figure 3d). A three-way ANOVA (pretreatment × morphine dose × days) revealed a significant effect of all three main factors (F1,20=6.05, p<0.05; F1,20=56.62, p<0.001; and F5,100=11.01, p<0.001, respectively) as well as the bifactorial interactions pretreatment × morphine dose (F1,20=6.20, p<0.05) and pretreatment × days (F5,100=3.72; p<0.001). Follow-up Tuckey HSD comparisons revealed that the administration of sodium butyrate did not affect the locomotor scores observed after repeated injections of a low dose of morphine (10 mg/kg), but it significantly boosted the acute and chronic effects of a higher dose (20 mg/kg, p<0.01). We further analyzed these effects of sodium butyrate by calculating the linear-regression equations describing the dynamic changes in morphine-induced locomotion across the different days of the induction phase. The equations corresponding to the regression lines describing the changes of morphine-induced locomotion over time in each group were SM10=9283.4+774.69x, r2=0.953; BM10=9092.0+825.47x, r2=0.683; SM20=20101.4+2284.9x, r2=0.941; BM20=32289.4+3280.3x, r2=0.857. These results indicated that the intercept and the slope values of the regression line of the two groups treated with a low dose of morphine unable to trigger locomotor sensitization were almost identical and independent of the pretreatment conditions. Conversely, in mice receiving a higher morphine dose, both the intercept and the slope values of the regression line corresponding to the group of mice co-treated with sodium butyrate were larger than in the group co-treated with saline. These results indicate that sodium butyrate not only enhanced morphine locomotion, but also exerted a positive modulation of the mechanisms leading to the sensitization of this response with repeated morphine administration. Taken together, these data support an interactive effect rather than a merely additive effect of sodium butyrate and morphine in locomotor sensitization.
We also examined other addiction-related behaviors and found that sodium butyrate enhanced morphine-induced CPP (Figure 3e, t23=2.30, p<0.05). In contrast, but in close parallelism to the results obtained in experiments involving repeated ethanol administration, the same doses of this HDACi affect neither the development of tolerance to the analgesic effects of morphine (Figure 3f) nor the intensity of morphine withdrawal (Figure 3g).
Acute Administration of Drugs of Abuse does not Modify Bulk Chromatin Acetylation in the Striatum, but Triggers Histone H3 Phosphorylation
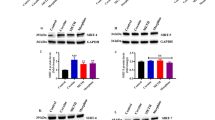
We assessed the ability of the drugs morphine, ethanol, and cocaine, at the concentrations used in our behavioral experiments (20 mg/kg, 2.5 g/kg and 10 mg/kg, respectively), to induce changes in the acetylation state of bulk chromatin in striatal neurons. To this end, we assessed acetylation of the four nucleosome histones, H2A, H2B, H3, and H4, as well as phosphorylation of histone H3. Although the administration of sodium butyrate increased bulk histone aceylation in a dose-dependent manner (Supplementary Figure S1), we found that neither of the three drugs had a significant effect on bulk histone acetylation levels. We, however, confirmed that cocaine administration increased phosphorylation of histone H3 at Ser10 (p<0.05, Figure 4a) (Brami-Cherrier et al, 2005; Kumar et al, 2005; Stipanovich et al, 2008). This increase in H3 phosphorylation was also probably responsible for the increase in H3 phosphoacetylation observed after cocaine administration (Brami-Cherrier et al, 2005; Kumar et al, 2005; Stipanovich et al, 2008). Similar results (Figure 4b) were obtained in response to higher doses of morphine (60 mg/kg), ethanol (6 g/kg i.p.), and cocaine (40 mg/kg i.p.). To confirm these results and, at the same time, gain spatial resolution and sensitivity in our assays, we performed immunohistochemistry experiments in sections of the brain of animals treated with high doses of morphine, cocaine, and ethanol using antibodies against the same histone modifications. As observed in the western blot analyses of striatal extracts, we could not detect significant changes in histone acetylation for neither of the four nucleosome histones (Supplementary Figure S2), but we found striking differences in the abitity of drugs to enhance histone H3 phosphorylation (Figure 4c; Supplementary Figure S3). Thus, whereas phospho-H3 antibody in brain sections of control mice only labeled proliferating cells in the subventricular zone (Supplementary Figure S3), the injection of cocaine caused, as reported earlier (Brami-Cherrier et al, 2005), a robust increase in histone phosphorylation in broad regions of the dorsal striatum, nucleus accumbens, and olfactory tubercle (Supplementary Table 2). Interestingly, we found that morphine increased histone H3 phosphorylation in the same brain regions than cocaine, although the magnitude and number of cells affected were lower, which may explain the absence of significant changes in response to morphine in our western blot analysis. Ethanol injection did not cause any obvious change in striatal histone H3 phosphorylation.

Modification of striatal bulk chromatin by drugs of abuse. (a, b) Upper schemes: drug administration dose (a: low, b: high) and protocol. Left: bar graph summarizing data from immunoblot analysis. Data (mean±SEM, four mice per group) are expressed relative to saline-treated control subjects, after normalization to β-actin. *p<0.05. Right: Representative immunoblots. (c) Representative immunostaining of coronal sections showing phospho-H3 reactivity at the medial portion of the dorsal striatum. Histone modification was analyzed 30 min after injection of saline (0.9% NaCl), morphine (60 mg/kg), ethanol (6 g/kg), and cocaine (40 mg/kg) intraperitoneal administration (three mice per experimental condition were analyzed and produced similar results). Scale bar: 100 μm. The diagram at the left indicates the striatal area showed in the pictures.
We also explored the interaction at the level of histone acetylation between sodium butyrate and acute administration of cocaine (10 mg/kg), ethanol (2.5 g/kg), or morphine (20 mg/kg). Sodium butyrate at the dose used in our behavioral analyses did not cause a significant increase in histone acetylation, neither alone nor in combination with either one of these drugs (Supplementary Figure S4A). In the case of morphine, we also examined whether changes in bulk chromatin acetylation became evident after chronic drug administration. For this purpose, mice received the same dose and treatment described earlier for behavioral sensitization and the tissue was collected 48 h after the last morphine injection. Repeated morphine injection did not cause changes in the bulk level of acetylation of neither of the four nucleosome histones or in the phosphorylation of histone H3 as examined by western blot analysis (Supplementary Figure S4B). The lack of significant changes in bulk chromatin acetylation after drug administration does not discard the fact that more subtle changes could take place at specific genomic loci or restricted neuronal populations.
Earlier Co-administration of Sodium Butyrate Alters the Transcriptional Response to Morphine Re-Exposure
To further investigate the molecular bases of sodium butyrate action on long-lasting behavioral responses to morphine, we screened for potential substrates of their interaction by performing a genome-wide comparison of the striatal transcriptome after chronic administration of morphine in the absence or presence of sodium butyrate. To this end, additional groups of mice underwent the same protocol described in the sensitization experiments (Figure 5a). Striatal RNA was extracted 1 h after the morphine challenge in groups of four animals that received either saline (N=6), or morphine (N=3), or the sodium butyrate–morphine co-treatment (N=3) during the sensitization induction phase. One-way ANOVA of microarray data identified 240 differentially expressed probe sets, corresponding to 197 identified genes (Figure 5b; Supplementary Table 3). Most of these genes (80%) were upregulated in response to the morphine challenge including both genes earlier identified as important in neuroplasticity, such as Arc and Nfkbia, and novel genes that may be highly relevant in additive behavior, such as Ttr and Kcnj13, which showed the largest response to chronic morphine administration in our study (Supplementary Figure S5). The list of genes altered in the mice that received the morphine–sodium butyrate co-treatment (BM group) was very similar to that obtained for the mice that received only morphine (SM group). Interestingly, in 105 genes (53% of those identified in the ANOVA), the sodium butyrate pretreatment led to an increase in expression when compared with animals treated with morphine alone. This upregulation was significant for 26 genes (13%), which represent interesting candidates to mediate HDACi effects; only 14 genes (7%) showed the opposite behavior. Of note is the presence, among the genes with an increased transcriptional response to morphine, of several circadian clock genes, namely Per1, Rev-erbα, and Cry1, whose expression was consistently and significantly affected by the HDACi pretreatment, indicating that the earlier described transcriptional feedback loop involving these genes was enhanced (Figure 5c). Other genes highly relevant in the context of drug addiction, such as c-fos, nr4a1, Zbtb16, and fosB, also showed the same trend toward enhanced upregulation by co-administration of sodium butyrate and morphine. We validated relevant changes by qRT–PCR (Figure 5d and e), confirming the boosting effect of sodium butyrate on a component of the transcriptional response to morphine.

Chronic inhibition of HDAC alters the genetic program activated by morphine administration. (a) Scheme of the regime injections and treatment groups used in the microarray experiment. (b) Hierarchical clustering of the 197 genes that were significantly affected by treatment in one-way ANOVA of microarray data and showed a fold change larger than 1.3 in at least one comparison between treatments. Candidate gene names for further validation are shown. (c) Several circadian rhythms genes showed a differential regulation in response to morphine (20 mg/kg) as result of their differential treatment during the sensitization induction phase. The upper inset shows that the changes in expression found in our analysis are in agreement with current models of their regulatory interactions (Etchegaray et al, 2003). (d, e) Quantitative real-time RT–PCR of selected transcripts identified in our screening as selectively upregulated by the co-treatment (d) or equally induced by morphine or morphine plus sodium butyrate (e). (*, **, ***p<0.05, p< 0.01, or p<0.001 as compared with SS group; (#, ##p<0.05 or p< 0.01 as compared with SM group). Data are presented as fold change (mean±SEM; eight mice per group).
DISCUSSION
Four important conclusions can be drawn from this study. First, the HDAC inhibitor sodium butyrate enhances some behavioral responses to drugs of abuse, such as locomotor sensitization or CPP. This interaction was observed for drugs of abuse of different pharmacological families, including psychostimulants, opioids, and ethanol. Second, the modulatory effects of histone acetylation cannot be generalized to all the behavioral effects derived from chronic exposure to these drugs of abuse (eg, tolerance and dependence). Third, we show that morphine, as earlier shown for cocaine, induces the phosphorylation of histone H3 in a restricted population of striatal neurons. In contrast, we could not detect significant changes in the acetylation state of bulk chromatin in striatal neurons triggered by either drug. Fourth, gene profiling analysis indicates that the interaction between morphine and HDACi first revealed at the behavioral level had a clear transcriptional correlate on specific loci highly relevant in neuroplasticity and addiction.
More specifically, our data suggest that HAT/HDAC activities have a prominent and selective function on the development of long-lasting behavioral effects, such as CPP and sensitization. These effects arise from non-homeostatic neuroplastic responses to drugs of abuse and are considered highly relevant in the development and maintenance of addictive behavior. Conversely, acetylation-related processes seem less relevant in the development of drug tolerance and dependence (as measured by the intensity of drug withdrawal), which result from transient homeostatic adaptations occurring within the cells and circuits directly stimulated by each drug and are not considered core symptoms of addiction (Hyman et al, 2006). This view is in agreement with earlier pharmacological and genetic studies on psychostimulants-induced sensitization and CPP (Bilbao et al, 2008; Kalda et al, 2007; Kumar et al, 2005; Levine et al, 2005; Renthal et al, 2007), as well as with the few preceding studies indicating that HDACis do not facilitate the development of tolerance (Wang et al, 2007) and that histone acetylation might be more involved in the expression than in the induction of drug dependence (Pandey et al, 2008). Future studies should further explore this intriguing dissociation.
This study also provides the most comprehensive analysis to date of posttranslational modifications of histones in the chromatin of striatal neurons in response to drugs of abuse. Earlier analyses on histone acetylation in response to cocaine (Brami-Cherrier et al, 2007, 2005; Cassel et al, 2006) or ethanol (Kim and Shukla, 2006; Pandey et al, 2008) have produced somewhat conflicting results. Despite the dose-dependent hyperacetylation observed in response to sodium butyrate, our analysis failed to reveal significant changes in bulk chromatin acetylation after acute administration of cocaine, ethanol, or morphine to saline or sodium butyrate-pretreated mice. These negative findings might result in the insufficient sensitivity of the techniques used here (ie, western blot and immunohistochemistry) to detect small increases over already high basal levels of histone acetylation, or more likely, of the unsuitability of these techniques to reveal subtle modifications of the chromatin that are presumably restricted to the promoters of specific genes in the nuclei of particular neuronal ensembles that are part of a large and cellularly heterogeneous brain structure such as the striatum. On the other hand, in agreement with earlier studies (Brami-Cherrier et al, 2005; Kumar et al, 2005; Stipanovich et al, 2008), we found that cocaine administration caused the phosphorylation and/or phosphoacetylation of histone H3. The identification of these changes using western blot and immunohistochemical procedures was probably favored by the very low basal level of H3 phosphorylation and/or phosphoacetylation in saline-treated mice. Further, our results also present the first evidence indicating that the activation of this signaling pathway is not exclusive of cocaine, as morphine administration also increased H3 phosphorylation in several brain areas, although to a lower extent than cocaine. In contrast, ethanol administration did not result in any change on bulk histone acetylation/phosphorylation identifiable by western blot or immunistochemical procedures. Ethanol could still promote changes in histone H3 phosphorylation that are below the sensitivity of our assays. Alternatively, it is also possible that ethanol does not activate the signaling pathway that leads to H3 phosphorylation, but can still interact with sodium butyrate.
We also observed that HDACi and morphine interact with the regulation of the transcription of several genes. Thus, our microarray analysis, in addition to confirm a number of transcriptional targets of morphine (Korostynski et al, 2007; McClung et al, 2005) and reveal novel ones, provided a short list of candidate genes to have a function in HDACi-mediated enhancement of non-homeostatic behavioral responses to morphine. Particularly remarkable is the case of circadian clock genes. In this regard, we observed an enhanced upregulation of Per1 in mice that received the sodium butyrate as coadjuvant treatment of repeated morphine injections. Although we did not assess chromatin acetylation at this specific locus, it has been shown that both HDACi (Naruse et al, 2004) and cocaine (Renthal et al, 2009) can induce histone acetylation at the Per1 promoter and increase its transcription. Interestingly, the enhanced expression of Per1 in mice co-treated with sodium butyrate and morphine was associated to increased downregulation of Cry1 and upregulation of Rev-erbα (Figure 5c). These changes are in good agreement with current models indicating that Rev-erbα acts as a potent repressor of Cry1 expression and that Cry1 works as a negative regulator of Per1 expression (Etchegaray et al, 2003). The potential behavioral relevance of these changes in Per1 expression is highlighted by studies showing that both mice and Drosophila mutants, lacking, respectively, Per1 or Per, failed to show sensitization to cocaine (Abarca et al, 2002; Andretic et al, 1999), and the reduction of Per1 activity in mice by DNAzyme led to a reduction in CPP for morphine (Li et al, 2008). The mechanisms by which circadian clock genes regulate addictive behavior remain elusive (Perreau-Lenz et al, 2007), but probably involve their regulatory function on dopamine receptor responsiveness (Andretic and Hirsh, 2000).
Although circadian clock genes are appealing candidates for mediating the interaction between sodium butyrate and morphine, the transcriptional program activated by chronic morphine is broad (Korostynski et al, 2007; McClung et al, 2005) and its interaction with sodium butyrate complex (Figure 5; Supplementary Figure S5). Besides circadian genes, our results show that morphine and sodium butyrate interact on the regulation of the expression of several other transcription factors that may lead to further transcriptional changes, such as fosB, the activity-regulated transcription factors Npas4 and Nr4al, and the transcriptional repressor Zbtb16 that positively regulates the ERK pathway and can potentially enhance drug effects. Genes encoding proteins involved in neurite outgrowth and structural changes (Mpp7, Btg2, Cdc42ep2, Rem2, Cdh9) or that may contribute to the enhanced behavioral response observed during sensitization, such as the translation enhancer Rbm3, were also differentially regulated. All these genes represent interesting candidates for further analysis of epigenetic regulation of addiction-related behaviors.
Taken together, our behavioral, histone posttranslational modification and gene profiling studies support a scenario in which drug-induced changes in the chromatin would be restricted to specific genomic loci relevant to neuronal plasticity, rather than global, genome-wide, changes in chromatin acetylation and gene expression. These loci are the most likely sites in which HDACi and drugs of abuse interact to promote non-homeostatic neuroadaptations that underlie behavioral phenomena relevant to the context of addictive behavior, such as CPP and sensitization. This view is supported by a number of recent studies exploring changes in histone phosphorylation and acetylation at the promoters of Per1, c-fos, fosB, bdnf, and NK1R, among other genes, in response to the acute administration of HDACi or drugs of abuse (Kumar et al, 2005; Levine et al, 2005; Renthal et al, 2008a, 2009, 2007; Russo et al, 2009; Schroeder et al, 2008). However, it should be also noted that HDACs deacetylate not only nucleosome histones, but also a much wider range of cellular proteins and can, therefore, also influence cell physiology and animal behavior through non-genomic mechanisms (Glozak et al, 2005; Spange et al, 2009). Indeed, some behavioral consequences observed after the administration of HDACi might be difficult to reconcile with the temporal requirements of the genomic actions of these compounds, which imply not only gene transcription but also the synthesis of proteins de novo and their transport to cellular structures/organelles. In agreement with this view, others (Kumar et al, 2005) and we have observed a rapid enhancement of cocaine-, ethanol- or morphine-induced locomotion when these drugs were injected simultaneously or shortly after (ie, 20 min) HDACi administration. Therefore, it is possible that these compounds influence drug-induced behaviors by epigenetic and non-epigenetic mechanisms. Initial support for this dual action of HDACi was obtained in our detailed analysis on the effects of sodium butyrate on morphine-induced locomotor sensitization, which revealed that this compound affected both the intercept and the slope of the regression line describing the progressive enhancement of locomotion observed after repeated administration of this drug. Future research should further explore the relative contribution of the different molecular changes triggered by sodium butyrate on the behavioral effects of its co-administration with drugs of abuse.
In summary, our results confirm and extend earlier reports on the ability of HDACi to modify the behavioral effects of drugs of abuse belonging to different pharmacological families. This modulatory activity is complex and might imply different mechanisms when considering acute vs chronic drug administration. HDAC inhibition also seems to differentially affect addiction-related behaviors associated with non-homeostatic neuroplasticity and those resulting from homeostatic adaptations to the drug. Future studies should explore further how drugs of abuse can trigger changes in histone acetylation and phosphorylation at specific genomic loci and determine whether these transient chromatin modifications can lead to more stable and specific molecular marks, such as histone or DNA methylation (Borrelli et al, 2008) that could better account for the persistence of addiction features revealed in our locomotor sensitization studies, in which the boosting effect of HDAC inhibition was maintained even 1 week after the last sodium butyrate injection. Our microarray screen has revealed a number of interesting candidates for such in-depth epigenetic analysis.
See related commentary by Adachi and Monteggia on page 2619.
Accession codes
References
Abarca C, Albrecht U, Spanagel R (2002). Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc Natl Acad Sci USA 99: 9026–9030.
Andretic R, Chaney S, Hirsh J (1999). Requirement of circadian genes for cocaine sensitization in Drosophila. Science 285: 1066–1068.
Andretic R, Hirsh J (2000). Circadian modulation of dopamine receptor responsiveness in Drosophila melanogaster. Proc Natl Acad Sci USA 97: 1873–1878.
Bilbao A, Parkitna JR, Engblom D, Perreau-Lenz S, Sanchis-Segura C, Schneider M et al (2008). Loss of the Ca2+/calmodulin-dependent protein kinase type IV in dopaminoceptive neurons enhances behavioral effects of cocaine. Proc Natl Acad Sci USA 105: 17549–17554.
Borrelli E, Nestler EJ, Allis CD, Sassone-Corsi P (2008). Decoding the epigenetic language of neuronal plasticity. Neuron 60: 961–974.
Brami-Cherrier K, Lavaur J, Pages C, Arthur JS, Caboche J (2007). Glutamate induces histone H3 phosphorylation but not acetylation in striatal neurons: role of mitogen- and stress-activated kinase-1. J Neurochem 101: 697–708.
Brami-Cherrier K, Valjent E, Herve D, Darragh J, Corvol JC, Pages C et al (2005). Parsing molecular and behavioral effects of cocaine in mitogen- and stress-activated protein kinase-1-deficient mice. J Neurosci 25: 11444–11454.
Cassel S, Carouge D, Gensburger C, Anglard P, Burgun C, Dietrich JB et al (2006). Fluoxetine and cocaine induce the epigenetic factors MeCP2 and MBD1 in adult rat brain. Mol Pharmacol 70: 487–492.
Chao J, Nestler EJ (2004). Molecular neurobiology of drug addiction. Annu Rev Med 55: 113–132.
Etchegaray JP, Lee C, Wade PA, Reppert SM (2003). Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature 421: 177–182.
Gililland KR, Finn DA (2007). The impact of gonadectomy and adrenalectomy on acute withdrawal severity in male and female C57BL/6J and DBA/2J mice following a single high dose of ethanol. Alcohol Clin Exp Res 31: 1846–1857.
Glozak MA, Sengupta N, Zhang X, Seto E (2005). Acetylation and deacetylation of non-histone proteins. Gene 363: 15–23.
Hyman SE, Malenka RC, Nestler EJ (2006). Neural mechanisms of addiction: the role of reward-related learning and memory. Annu Rev Neurosci 29: 565–598.
Kalda A, Heidmets LT, Shen HY, Zharkovsky A, Chen JF (2007). Histone deacetylase inhibitors modulates the induction and expression of amphetamine-induced behavioral sensitization partially through an associated learning of the environment in mice. Behav Brain Res 181: 76–84.
Kim JS, Shukla SD (2006). Acute in vivo effect of ethanol (binge drinking) on histone H3 modifications in rat tissues. Alcohol Alcohol 41: 126–132.
Kliethermes CL (2005). Anxiety-like behaviors following chronic ethanol exposure. Neurosci Biobehav Rev 28: 837–850.
Korostynski M, Piechota M, Kaminska D, Solecki W, Przewlocki R (2007). Morphine effects on striatal transcriptome in mice. Genome Biol 8: R128.
Kumar A, Choi KH, Renthal W, Tsankova NM, Theobald DE, Truong HT et al (2005). Chromatin remodeling is a key mechanism underlying cocaine-induced plasticity in striatum. Neuron 48: 303–314.
Levine AA, Guan Z, Barco A, Xu S, Kandel ER, Schwartz JH (2005). CREB-binding protein controls response to cocaine by acetylating histones at the fosB promoter in the mouse striatum. Proc Natl Acad Sci USA 102: 19186–19191.
Li SX, Wang ZR, Li J, Peng ZG, Zhou W, Zhou M et al (2008). Inhibition of Period1 gene attenuates the morphine-induced ERK-CREB activation in frontal cortex, hippocampus, and striatum in mice. Am J Drug Alcohol Abuse 34: 673–682.
Lopez de Armentia M, Jancic D, Olivares R, Alarcon JM, Kandel ER, Barco A (2007). cAMP response element-binding protein-mediated gene expression increases the intrinsic excitability of CA1 pyramidal neurons. J Neurosci 27: 13909–13918.
Maldonado R, Saiardi A, Valverde O, Samad TA, Roques BP, Borrelli E (1997). Absence of opiate rewarding effects in mice lacking dopamine D2 receptors. Nature 388: 586–589.
McClung CA, Nestler EJ (2008). Neuroplasticity mediated by altered gene expression. Neuropsychopharmacology 33: 3–17.
McClung CA, Nestler EJ, Zachariou V (2005). Regulation of gene expression by chronic morphine and morphine withdrawal in the locus ceruleus and ventral tegmental area. J Neurosci 25: 6005–6015.
Naruse Y, Oh-hashi K, Iijima N, Naruse M, Yoshioka H, Tanaka M (2004). Circadian and light-induced transcription of clock gene Per1 depends on histone acetylation and deacetylation. Mol Cell Biol 24: 6278–6287.
Pandey SC, Ugale R, Zhang H, Tang L, Prakash A (2008). Brain chromatin remodeling: a novel mechanism of alcoholism. J Neurosci 28: 3729–3737.
Perreau-Lenz S, Zghoul T, Spanagel R (2007). Clock genes running amok. Clock genes and their role in drug addiction and depression. EMBO Rep 8 Spec No: S20–S23.
Renthal W, Carle TL, Maze I, Covington III HE, Truong HT, Alibhai I et al (2008a). Delta FosB mediates epigenetic desensitization of the c-fos gene after chronic amphetamine exposure. J Neurosci 28: 7344–7349.
Renthal W, Kumar A, Xiao G, Wilkinson M, Covington III HE, Maze I et al (2009). Genome-wide analysis of chromatin regulation by cocaine reveals a role for sirtuins. Neuron 62: 335–348.
Renthal W, Maze I, Krishnan V, Covington III HE, Xiao G, Kumar A et al (2007). Histone deacetylase 5 epigenetically controls behavioral adaptations to chronic emotional stimuli. Neuron 56: 517–529.
Renthal W, Nestler EJ (2008b). Epigenetic mechanisms in drug addiction. Trends Mol Med 14: 341–350.
Russo SJ, Wilkinson MB, Mazei-Robison MS, Dietz DM, Maze I, Krishnan V et al (2009). Nuclear factor kappaB signaling regulates neuronal morphology and cocaine reward. J Neurosci 29: 3529–3537.
Schroeder FA, Penta KL, Matevossian A, Jones SR, Konradi C, Tapper AR et al (2008). Drug-induced activation of dopamine D(1) receptor signaling and inhibition of class I/II histone deacetylase induce chromatin remodeling in reward circuitry and modulate cocaine-related behaviors. Neuropsychopharmacology 33: 2981–2992.
Spange S, Wagner T, Heinzel T, Kramer OH (2009). Acetylation of non-histone proteins modulates cellular signalling at multiple levels. Int J Biochem Cell Biol 41: 185–198.
Stipanovich A, Valjent E, Matamales M, Nishi A, Ahn JH, Maroteaux M et al (2008). A phosphatase cascade by which rewarding stimuli control nucleosomal response. Nature 453: 879–884.
Sweatt JD (2009). Experience-dependent epigenetic modifications in the central nervous system. Biol Psychiatry 65: 191–197.
Tsankova N, Renthal W, Kumar A, Nestler EJ (2007). Epigenetic regulation in psychiatric disorders. Nat Rev Neurosci 8: 355–367.
Wang Y, Krishnan HR, Ghezzi A, Yin JC, Atkinson NS (2007). Drug-induced epigenetic changes produce drug tolerance. PLoS Biol 5: e265.
Acknowledgements
We thank Alicia Dosda, Gemma Caballer, and Maria Jimenez for technical assistance and Luis Valor for critical reading of the manuscript. Research at AB's laboratory was supported by the European Commission grant MEXT-CT-2003-509550, the Spanish Ministry of Science and Innovation Grants SAF2008-00611 and CSD2007-00023, and grants from Fundación Ramón Areces and Fundació la Marató de TV3. Research at CSS's laboratory was funded by a grant of the Conselleria d'Educació de la Generalitat Valenciana (GV/2007/098). CSS holds a Ramón y Cajal contract and JLA holds a Juan de la Cierva contract supported by the Ministry of Science and Innovation.
Author information
Authors and Affiliations
Corresponding authors
Additional information
DISCLOSURE
The authors declare no conflict of interest.
Supplementary Information accompanies the paper on the Neuropsychopharmacology website (http://www.nature.com/npp)
Supplementary information
Rights and permissions
About this article
Cite this article
Sanchis-Segura, C., Lopez-Atalaya, J. & Barco, A. Selective Boosting of Transcriptional and Behavioral Responses to Drugs of Abuse by Histone Deacetylase Inhibition. Neuropsychopharmacol 34, 2642–2654 (2009). https://doi.org/10.1038/npp.2009.125
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2009.125
Keywords
This article is cited by
-
Single nucleus transcriptomic analysis of rat nucleus accumbens reveals cell type-specific patterns of gene expression associated with volitional morphine intake
Translational Psychiatry (2022)
-
Npas4 impairs fear memory via phosphorylated HDAC5 induced by CGRP administration in mice
Scientific Reports (2021)
-
Molecular Mechanism of Neuroprotective Effect of Melatonin on Morphine Addiction and Analgesic Tolerance: an Update
Molecular Neurobiology (2021)
-
Interruption of continuous opioid exposure exacerbates drug-evoked adaptations in the mesolimbic dopamine system
Neuropsychopharmacology (2020)
-
CBP and SRF co-regulate dendritic growth and synaptic maturation
Cell Death & Differentiation (2019)
