
Abstract
Body fluid conditions are continuously monitored in the brain to regulate thirst and salt-appetite sensations. Angiotensin II drives both thirst and salt appetite; however, the neural mechanisms underlying selective water- and/or salt-intake behaviors remain unknown. Using optogenetics, we show that thirst and salt appetite are driven by distinct groups of angiotensin II receptor type 1a-positive excitatory neurons in the subfornical organ. Neurons projecting to the organum vasculosum lamina terminalis control water intake, while those projecting to the ventral part of the bed nucleus of the stria terminalis control salt intake. Thirst-driving neurons are suppressed under sodium-depleted conditions through cholecystokinin-mediated activation of GABAergic neurons. In contrast, the salt appetite-driving neurons were suppressed under dehydrated conditions through activation of another population of GABAergic neurons by Nax signals. These distinct mechanisms in the subfornical organ may underlie the selective intakes of water and/or salt and may contribute to body fluid homeostasis.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Change history
31 January 2017
In the PDF and print versions of this article initially published, the year in the online publication date was given as 2017 instead of 2016. The error has been corrected in the PDF version of the article.
References
Stricker, E.M. & Sved, A.F. Thirst. Nutrition 16, 821–826 (2000).
Rowland, N.E., Farnbauch, L.J. & Crews, E.C. Sodium deficiency and salt appetite in ICR: CD1 mice. Physiol. Behav. 80, 629–635 (2004).
Noda, M. & Hiyama, T.Y. Sodium sensing in the brain. Pflugers Arch. 467, 465–474 (2015a).
Kenney, W.L. & Chiu, P. Influence of age on thirst and fluid intake. Med. Sci. Sports Exerc. 33, 1524–1532 (2001).
Geerling, J.C. & Loewy, A.D. Central regulation of sodium appetite. Exp. Physiol. 93, 177–209 (2008).
Gabor, A. & Leenen, F.H. Central neuromodulatory pathways regulating sympathetic activity in hypertension. J. Appl. Physiol. 113, 1294–1303 (2012).
Johnson, A.K. The sensory psychobiology of thirst and salt appetite. Med. Sci. Sports Exerc. 39, 1388–1400 (2007).
McKinley, M.J. et al. The sensory circumventricular organs of the mammalian brain. Adv. Anat. Embryol. Cell Biol. 172, III–XII, 1–122, back cover (2003).
Hiyama, T.Y. et al. Nax channel involved in CNS sodium-level sensing. Nat. Neurosci. 5, 511–512 (2002).
Noda, M. & Hiyama, T.Y. Sodium-level-sensitive sodium channel and salt-intake behavior. Chem. Senses 30 (Suppl. 1), i44–i45 (2005).
Noda, M. The subfornical organ, a specialized sodium channel, and the sensing of sodium levels in the brain. Neuroscientist 12, 80–91 (2006).
Watanabe, E. et al. Sodium-level-sensitive sodium channel Nax is expressed in glial laminate processes in the sensory circumventricular organs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 290, R568–R576 (2006).
Hiyama, T.Y. et al. Endothelin-3 expression in the subfornical organ enhances the sensitivity of Nax, the brain sodium-level sensor, to suppress salt intake. Cell Metab. 17, 507–519 (2013).
Noda, M. & Hiyama, T.Y. The Nax channel: What it is and what it does. Neuroscientist 21, 399–412 (2015b).
Hiyama, T.Y., Watanabe, E., Okado, H. & Noda, M. The subfornical organ is the primary locus of sodium-level sensing by Nax sodium channels for the control of salt-intake behavior. J. Neurosci. 24, 9276–9281 (2004).
Watanabe, E. et al. Nav2/NaG channel is involved in control of salt-intake behavior in the CNS. J. Neurosci. 20, 7743–7751 (2000).
Hiyama, T.Y. et al. Autoimmunity to the sodium-level sensor in the brain causes essential hypernatremia. Neuron 66, 508–522 (2010).
Shimizu, H. et al. Glial Nax channels control lactate signaling to neurons for brain [Na+] sensing. Neuron 54, 59–72 (2007).
Paul, M., Poyan Mehr, A. & Kreutz, R. Physiology of local renin-angiotensin systems. Physiol. Rev. 86, 747–803 (2006).
Buggy, J. & Fisher, A.E. Evidence for a dual central role for angiotensin in water and sodium intake. Nature 250, 733–735 (1974).
Fitzsimons, J.T. Angiotensin, thirst, and sodium appetite. Physiol. Rev. 78, 583–686 (1998).
Premer, C., Lamondin, C., Mitzey, A., Speth, R.C. & Brownfield, M.S. Immunohistochemical localization of AT1a, AT1b, and AT2 angiotensin II receptor subtypes in the rat adrenal, pituitary, and brain with a perspective commentary. Int. J. Hypertens. 2013, 175428 (2013).
Sugaya, T. et al. Angiotensin II type 1a receptor-deficient mice with hypotension and hyperreninemia. J. Biol. Chem. 270, 18719–18722 (1995).
Matsusaka, T. et al. Angiotensin receptor blocker protection against podocyte-induced sclerosis is podocyte angiotensin II type 1 receptor-independent. Hypertension 55, 967–973 (2010).
Vong, L. et al. Leptin action on GABAergic neurons prevents obesity and reduces inhibitory tone to POMC neurons. Neuron 71, 142–154 (2011).
Tamamaki, N. et al. Green fluorescent protein expression and colocalization with calretinin, parvalbumin, and somatostatin in the GAD67-GFP knock-in mouse. J. Comp. Neurol. 467, 60–79 (2003).
Oka, Y., Ye, M. & Zuker, C.S. Thirst driving and suppressing signals encoded by distinct neural populations in the brain. Nature 520, 349–352 (2015).
Kato, S. et al. A lentiviral strategy for highly efficient retrograde gene transfer by pseudotyping with fusion envelope glycoprotein. Hum. Gene Ther. 22, 197–206 (2011).
Gladden, L.B. Lactate metabolism: a new paradigm for the third millennium. J. Physiol. (Lond.) 558, 5–30 (2004).
Willis, G.L., Hansky, J. & Smith, G.C. Ventricular, paraventricular and circumventricular structures involved in peptide-induced satiety. Regul. Pept. 9, 87–99 (1984).
Simpson, J.B. & Routtenberg, A. Subfornical organ: site of drinking elicitation by angiotensin II. Science 181, 1172–1175 (1973).
McKinley, M.J., Badoer, E. & Oldfield, B.J. Intravenous angiotensin II induces Fos-immunoreactivity in circumventricular organs of the lamina terminalis. Brain Res. 594, 295–300 (1992).
Simpson, J.B. & Routtenberg, A. Subfornical organ lesions reduce intravenous angiotensin-induced drinking. Brain Res. 88, 154–161 (1975).
Nation, H.L., Nicoleau, M., Kinsman, B.J., Browning, K.N. & Stocker, S.D. DREADD-induced activation of subfornical organ neurons stimulates thirst and salt appetite. J. Neurophysiol. 115, 3123–3129 (2016).
Abbott, S.B., Machado, N.L., Geerling, J.C. & Saper, C.B. Reciprocal control of drinking behavior by median preoptic neurons in mice. J. Neurosci. 36, 8228–8237 (2016).
Li, Z. et al. Role of AT2 receptor in the brain in regulation of blood pressure and water intake. Am. J. Physiol. Heart Circ. Physiol. 284, H116–H121 (2003).
Ahmed, A.S., Dai, L., Ho, W., Ferguson, A.V. & Sharkey, K.A. The subfornical organ: a novel site of action of cholecystokinin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 306, R363–R373 (2014).
Noble, F. & Roques, B.P. CCK-B receptor: chemistry, molecular biology, biochemistry and pharmacology. Prog. Neurobiol. 58, 349–379 (1999).
Weiland, T.J., Voudouris, N.J. & Kent, S. The role of CCK2 receptors in energy homeostasis: insights from the CCK2 receptor-deficient mouse. Physiol. Behav. 82, 471–476 (2004).
Garfield, A.S. et al. A parabrachial-hypothalamic cholecystokinin neurocircuit controls counterregulatory responses to hypoglycemia. Cell Metab. 20, 1030–1037 (2014).
D'Agostino, G. et al. Appetite controlled by a cholecystokinin nucleus of the solitary tract to hypothalamus neurocircuit. eLife 5, e12225 (2016).
Sakuta, H., Nishihara, E., Hiyama, T.Y., Lin, C.H. & Noda, M. Nax signaling evoked by an increase in [Na+] in CSF induces water intake via EET-mediated TRPV4 activation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 311, R299–R306 (2016).
Zardetto-Smith, A.M., Beltz, T.G. & Johnson, A.K. Role of the central nucleus of the amygdala and bed nucleus of the stria terminalis in experimentally-induced salt appetite. Brain Res. 645, 123–134 (1994).
Noda, M. & Sakuta, H. Central regulation of body-fluid homeostasis. Trends Neurosci. 36, 661–673 (2013).
Han, X. et al. A high-light sensitivity optical neural silencer: development and application to optogenetic control of non-human primate cortex. Front. Syst. Neurosci. 5, 18 (2011).
Acknowledgements
We thank Y. Yanagawa (Gunma University, Japan) for the GAD67-GFP mice and K. Deisseroth (Stanford University, USA) for the hChR2(H134R) gene. We also thank S. Miura and T. Hashimoto for their technical assistance and A. Kodama for her secretarial assistance. This work was supported by MEXT/JSPS KAKENHI (grant numbers; 24220010 to M.N.; 25136723 and 26293043 to T.Y.H.; and 14J00037 to T. Matsuda.), Takeda Science Foundation, Brain Science Foundation, the Salt Science Research Foundation and the NOVARTIS Foundation (Japan) for the Promotion of Science. This work was supported also by the Okazaki ORION project and Cooperative Study Program of National Institute for Physiological Sciences.
Author information
Authors and Affiliations
Contributions
T.Y.H. and M.N. designed the research; T. Matsuda and T.Y.H. performed the experiments; T. Matsuda, T.Y.H. and M.N. analyzed the data; F.N. and T. Matsusaka developed AT1aloxP/loxP mice; A.F. developed AT1a-KO mice; Kenta Kobayashi and Kazuto Kobayashi provided the viral vectors; and M.N., T.Y.H. and T. Matsuda wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Visualization of AT1a-positive cells using AT1alacZ/+ mouse.
(a–f) Immunohistochemical detection of β-gal in the mouse brain. Coronal sections at the respective anteroposterior positions from the bregma, -0.71 (a), +0.38 (b), -7.47 (c), +1.21 (d), -0.11 (e), and -1.79 (f) mm of AT1alacZ/+ mice. AcbC, core region of the accumbens nucleus; AcbSh, shell region of the accumbens nucleus; Arc, arcuate hypothalamic nucleus; CPu, caudate putamen; DMD, dorsal part of the dorsomedial hypothalamic nucleus; LH, lateral hypothalamic area; LSD, dorsal part of the lateral septal nucleus; mfb, medial forebrain bundle; MnPO, median preoptic nucleus; MPA, medial preoptic area; MPO, medial preoptic nucleus; NTS, nucleus of the solitary tract; PVN, paraventricular nucleus; SON, supraoptic nucleus; VLPO, ventrolateral preoptic nucleus. Scale bars; 50 μm (red), 250 μm (white).
Supplementary Figure 2 AT1a-positive neurons in area postrema and PVN were not activated under the Na-depleted condition.
Left: immunohistochemical detection of Fos in area postrema (AP) and PVN of AT1alacZ/+ mice under the Na-depleted condition. Right: summary of Fos-positive cell counts in the respective brain regions (n = 5 mice each, U(AP) = 6, P = 0.2100; U (PVN) = 16, P = 0.5308). Scale bars, 50 μm. ns, not significant; Mann-Whitney U-tests. Data show mean ± s.e.m.
Supplementary Figure 3 Inhibition of AT1 signaling in the brain.
(a) Left: immunohistochemical detection of Fos in SFO and OVLT under the Na-depleted condition. Right: summary of Fos-positive cell counts in these brain loci (n = 5 mice each; U (SFO) = 25, P = 0.0090; U (OVLT) = 25, P = 0.0090). (b) Effects of the continuous intracerebroventricular infusion of losartan (10 μg/h) on water and 0.3 M NaCl intake by WT mice under the Na-depleted condition (n = 6 mice for vehicle, n = 8 mice for losartan; U (Water) = 38, P = 0.0810; U (NaCl) = 48, P = 0.0024). Scale bars, 50 μm. bw, body weight; ns, not significant; **P < 0.01; Mann-Whitney U-tests. Data show mean ± s.e.m.
Supplementary Figure 4 Local deletion of the Agtr1a gene expression in the SFO of AT1aloxP/loxP mouse.
Immunohistochemical detection of Cre and in situ hybridization of AT1a mRNA in the SFO of AT1aloxP/loxP mouse with (AAV-Cre) or without (Control) AAV-Cre virus injection. Scale bar, 50 μm. Expression of AT1a proteins was markedly reduced by Cre recombinase.
Supplementary Figure 5 Most though not all AT1a-positive neurons express neuronal nitric oxide synthase (nNOS) in SFO.
Immunohistochemical detection of β-gal and nNOS proteins in the SFO of AT1alacZ/+ mouse. Scale bar, 50 μm.
Supplementary Figure 6 Projection targets of glutamatergic and GABAergic neurons in SFO.
(a) Injection of AAV-DIO-EGFP into the SFO of the Vglut2-Cre mouse. Top: immunohistochemical detection of EGFP in the injection site. Bottom: projection sites. (b–e) Top: injections of HiRet-EGFP into OVLT (b), vBNST(c), PVN (d), and SON (e). Bottom: immunohistochemical detection of EGFP in the injection sites and retrogradely labeled cells (SFO). (f) Left: injection of AAV-DIO-EGFP into the SFO of the Vgat-Cre mouse. Middle and right: immunohistochemical detection of EGFP in the injection site and projection sites. (g) Left: injection of tetramethylrhodamine-conjugated dextran (Dextran) into the SFO of the WT mouse. Middle and right: fluorescence images of tetramethylrhodamine in the injection site and projection sites. Scale bars; 50 μm (SFO), 100 μm (OVLT, MnPO, vBNST, PVN, and SON).
Supplementary Figure 7 Manipulation of neuronal activity by optical excitation and silencing.
(a) Left: injection of AAV-DIO-ChR2-EGFP or AAV-DIO-ArchT-GFP into the SFO of the Vglut2-Cre mouse. Right: cell-attached recordings of the action potential firing in ChR2- and ArchT-positive SFO cells in the slice prepared from the mice with respective virus infection. Regarding ArchT-positive neurons, the experiments were performed in the presence of Ang II (0.1 μM). Blue and yellow lines indicate the period of the respective light exposure. Blue or yellow light caused excitation or suppression of the firing activity, respectively. Scale bars, 1 s. (b) Top: injection of AAV-DIO-ChR2-EGFP into the SFO of the Vglut2-Cre mouse. Bottom: immunohistochemical detection of Fos and EGFP after the optical excitation of the OVLT of mice with (ChR2) or without (Control) infection of AAV-DIO-ChR2-EGFP. After the optical exposure, the number of Fos-positive cells was drastically elevated in the OVLT with ChR2-EGFP expression. The position of the tip of optic fiber (Opt. fiber) was set just above OVLT. Scale bars, 50 μm. (c) Top: injection of HiRet-EGFP or HiRet-ArchT-EGFP into the vBNST of the WT mouse. Bottom: immunohistochemical detection of Fos and EGFP after the optical silencing of SFO under the Na-depleted condition. Arrow heads indicate double-positive cells (left panel). After the optical silencing, Fos expression was not observed in the ArchT-GFP expressing neurons (right panel), and no double-positive cells were detected. Scale bars, 50 μm.
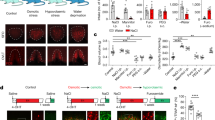
Supplementary Figure 8 Optical silencing of the SFO→OVLT pathway.
(a) Top: experimental protocol to observe water intake induced by the furosemide injection and gray scale heat maps of water intake by WT and AT1a-KO mice after the furosemide injection under the water- and Na-depleted condition (W/Na-D). Bottom: summary of the one-bottle test (n = 10 mice each; U (WT) = 4.5, P = 0.0007; U(AT1a-KO) = 52.5, P = 0.8789). (b) Top, left: injection of HiRet-Cre into the OVLT of the AT1aloxP/loxP mouse. Top, right: experimental protocol of the one-bottle test under W/Na-D, and gray scale heat maps of water intake under W/Na-D. Bottom, left: summary of the one-bottle test (n = 7 mice each; U = 49, P = 0.0022). For the summary, mice, in which more than 200 (/mm2) Cre-positive cells were detected in SFO, were used. Bottom, right: the relationship between the number of Cre-positive cells in SFO and water intake in the one-bottle test (n = 11 mice each; r(WT) = 0.0608, P = 0.8589; r(loxP-flanked Agtr1a) = -0.8152, P = 0.0022; Pearson correlation analysis). Linear regression lines for WT-Cre and AT1aloxP/loxP-Cre are shown in black and red, respectively. (c) Left: experimental protocols of one-bottle test by WT mice injected HiRet-ArchT-GFP into OVLT with the optical stimulation of SFO under W/Na-D, and gray scale heat maps of water intake. Right: summary of water intake (n = 8 mice each; U = 58, P = 0.0074). bw, body weight; ns, not significant; **P < 0.01; all tests are Mann-Whitney U-tests unless otherwise stated. Data show mean ± s.e.m.
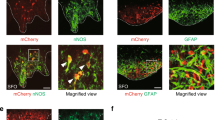
Supplementary Figure 9 Two separate SFO neurons project to vBNST and OVLT.
Top: HiRet-EGFP and HiRet-mCherry were injected into the OVLT and vBNST of the WT mouse, respectively. Bottom: immunohistochemical detections of EGFP (left panel) and mCherry (middle panel) in the retrogradely labeled cells in SFO. Scale bar, 50 μm.
Supplementary Figure 10 The SFO→vBNST pathway is involved in salt-intake but not water-intake behavior.
(a) Left: immunohistochemical detection of Fos in vBNST under the Na-depleted condition. Right: summary of Fos-positive cell counts (n = 5 mice each; U = 1, P = 0.0216). Scale bar, 50 μm. (b) Electrolytic lesions in bilateral vBNST. Representative coronal sections show the lesioned areas. Scale bar, 600 μm. (c) Left: gray scale heat maps of water and 0.3 M NaCl intakes showing the effects of lesions on their intakes under the Na-depleted condition. Right: summary of intake volumes (n = 8 mice each; U (Water) = 50, P = 0.0658; U (NaCl) = 63, P = 0.0013). (d–f) Top: injection of HiRet-Cre into OVLT (d), PVN (e), and SON (f) of the AT1aloxP/loxP mouse. Bottom: the relationship between the number of Cre-positive cells in SFO and 0.3 M NaCl intake in the two-bottle test under the Na-depleted condition (n = 11 mice for WT and n = 14 mice for AT1aloxP/loxP in d; r(WT) = -0.0729, P = 0.8311; r(loxP-flanked Agtr1a) = 0.1727, P = 0.5547: n = 9 mice each in e; r(WT) = 0.6492, P = 0.0584; r(loxP-flanked Agtr1a) = 0.3791, P = 0.3142: n = 8 mice for WT and n = 9 mice for AT1aloxP/loxP in f; r(WT) = -0.3860, P = 0.3448; r(loxP-flanked Agtr1a) = -0.0902, P = 0.8174; Pearson correlation analysis). Linear regression lines are shown in black (WT-Cre) and red (AT1aloxP/loxP-Cre). (g, h) Optical silencing of the SFO→vBNST pathway on water intake under the water-depleted (g) or water- and Na-depleted (h) condition. The relationship between the number of ArchT-GFP-positive cells in SFO and water intake with or without the optical stimulation (n = 7 mice for g; r(Opt–) = -0.2307, P = 0.6185; r (Opt+) = -0.1153, P = 0.8054: n = 9 mice for h; r (Opt–) = 0.3666, P = 0.3317; r (Opt+) = 0.1612, P = 0.6785; Pearson correlation analysis). bw, body weight; ns, not significant; *P < 0.05; **P < 0.01; all tests are Mann-Whitney U-tests unless otherwise stated. Data show mean ± s.e.m.
Supplementary Figure 11 Modulation of salt-intake behavior by GABAergic neurons in SFO.
Gray scale heat maps of water and 0.3 M NaCl intakes by individual mice under the Na-depleted condition with (Opt+) or without (Opt–) the optical excitation of GABAergic neurons in SFO. Summary of the data is shown in Fig. 7g. n = 7 mice each. bw, body weight.
Supplementary Figure 12 Neural activities of SFO(→vBNST) neurons and GABAergic neurons in the presence of CCK.
(a) Schematic drawing of the retrograde labeling of the SFO(→vBNST) neurons in GAD67-EGFP mice by using CTb-555. (b) Schematic drawing of electrical stimulation of a GABAergic neuron (green) and electrophysiological recording from a CTb-labelled neuron synaptically connected in SFO (red). (c) Representative spike frequency histogram and summary showing the effects of CCK (10 nM) and Ang II (0.1 μM) on the firing activities of the GABAergic neuron and the SFO(→vBNST) neuron in the SFO of WT mice (n = 8 slices each; U(GABAergic neuron) = 30, P = 0.8746; U (Projection neuron) = 28.5, P = 0.7525). Blue values are the mean ± s.e.m. of mice, respectively. ns, not significant; Mann-Whitney U-tests.
Supplementary Figure 13 A schematic overview of controls for thirst and salt appetite from SFO.
SFO is characterized by the presence of neuronal cell bodies and extensive networks of fenestrated capillaries that allow circulating Ang II to be released into the intercellular space. Their ventricular side is partitioned by an ependymal cell layer facing the third ventricle. Nax channels populate the perineural processes of astrocytes and ependymal cells, enveloping a population of GABAergic neurons. Salt neurons innervating vBNST receive inhibitory synaptic inputs from GABAergic neurons.
Under the water-depleted condition (left), Ang II and [Na+] both increase in SFO. Nax in glial cells is activated, which stimulates anaerobic glycolysis in glial cells, and eventually promotes the release of lactate, the end product of anaerobic glycolysis18. Lactate is metabolized to ATP in GABAergic neurons, leading to depolarization through a KATP channel-dependent mechanism and the up-regulation of the firing activity18. The GABAergic neurons thus activated by the Nax signal suppress salt neurons, but not water neurons. Water neurons innervate OVLT, and the selective activation of water neurons results in water intake under the water-depleted condition. However, some experimental data suggest that AT2 receptor-positive neurons may be also involved in water-intake control, and that some other water neurons are also present in SFO and OVLT, which are activated by signals of Na+, osmolality, or other dipsogenic hormones (see Discussion). Under the Na-depleted condition (right), salt neurons which innervate vBNST are activated by Ang II and enhance salt appetite. Under the Na-depleted condition, CCK is upregulated in SFO, and the secreted CCK suppresses the Ang II-dependent neural activities of water neurons via GABAergic neurons. This is a mechanism for the suppression of water intake under the Na-depleted condition. Under the wate r- and Na-depleted condition, both water and Na intakes are stimulated, because neither the Nax signal nor CCK signal is induced.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–13 and Supplementary Results and Discussion (PDF 2037 kb)
Rights and permissions
About this article

Cite this article
Matsuda, T., Hiyama, T., Niimura, F. et al. Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ. Nat Neurosci 20, 230–241 (2017). https://doi.org/10.1038/nn.4463
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.4463
This article is cited by
-
TMEM63B channel is the osmosensor required for thirst drive of interoceptive neurons
Cell Discovery (2024)
-
Comprehensive mapping of Epithelial Na+ channel α expression in the mouse brain
Brain Structure and Function (2024)
-
Sodium intake: a double-edged sword wielded by the brain
Science China Life Sciences (2024)
-
A kidney-brain neural circuit drives progressive kidney damage and heart failure
Signal Transduction and Targeted Therapy (2023)
-
Thirst: neuroendocrine regulation in mammals
Veterinary Research Communications (2023)
