
Abstract
Phenylketonuria (PKU) is a genetic disease that is characterized by an inability to metabolize phenylalanine (Phe), which can result in neurotoxicity. To provide a potential alternative to a protein-restricted diet, we engineered Escherichia coli Nissle to express genes encoding Phe-metabolizing enzymes in response to anoxic conditions in the mammalian gut. Administration of our synthetic strain, SYNB1618, to the Pahenu2/enu2 PKU mouse model reduced blood Phe concentration by 38% compared with the control, independent of dietary protein intake. In healthy Cynomolgus monkeys, we found that SYNB1618 inhibited increases in serum Phe after an oral Phe dietary challenge. In mice and primates, Phe was converted to trans-cinnamate by SYNB1618, quantitatively metabolized by the host to hippurate and excreted in the urine, acting as a predictive biomarker for strain activity. SYNB1618 was detectable in murine or primate feces after a single oral dose, permitting the evaluation of pharmacodynamic properties. Our results define a strategy for translation of live bacterial therapeutics to treat metabolic disorders.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Accession codes
Primary accessions
BioProject
-
PRJNA482064
Referenced accessions
GenBank/EMBL/DDBJ
-
KGM29850.1
NCBI Reference Sequence
-
U35383.1
References
de Groot, M.J., Hoeksma, M., Blau, N., Reijngoud, D.J. & van Spronsen, F.J. Pathogenesis of cognitive dysfunction in phenylketonuria: review of hypotheses. Mol. Genet. Metab. 99 (Suppl. 1), S86–S89 (2010).
Anderson, P.J. et al. Are neuropsychological impairments in children with early-treated phenylketonuria (PKU) related to white matter abnormalities or elevated phenylalanine levels? Dev. Neuropsychol. 32, 645–668 (2007).
Bilder, D.A. et al. Psychiatric symptoms in adults with phenylketonuria. Mol. Genet. Metab. 108, 155–160 (2013).
Romani, C. et al. The impact of phenylalanine levels on cognitive outcomes in adults with phenylketonuria: Effects across tasks and developmental stages. Neuropsychology 31, 242–254 (2017).
Vockley, J. et al. Phenylalanine hydroxylase deficiency: diagnosis and management guideline. Genet. Med. 16, 188–200 (2014).
Riva, M.A. et al. Work activity and phenylalanine levels in a population of young adults with classic PKU. Med. Lav. 108, 118–122 (2017).
Longo, N. et al. Long-term safety and efficacy of sapropterin: the PKUDOS registry experience. Mol. Genet. Metab. 114, 557–563 (2015).
Longo, N. et al. Long-term developmental progression in infants and young children taking sapropterin for phenylketonuria: a two-year analysis of safety and efficacy. Genet. Med. 17, 365–373 (2015).
Somaraju, U.R. & Merrin, M. Sapropterin dihydrochloride for phenylketonuria. Cochrane Database Syst. Rev. 16, CD008005 (2010).
Bell, S.M. et al. Formulation and PEGylation optimization of the therapeutic PEGylated phenylalanine ammonia lyase for the treatment of phenylketonuria. PLoS One 12, e0173269 (2017).
Sarkissian, C.N. et al. Preclinical evaluation of multiple species of PEGylated recombinant phenylalanine ammonia lyase for the treatment of phenylketonuria. Proc. Natl. Acad. Sci. USA 105, 20894–20899 (2008).
Bourget, L. & Chang, T.M. Effects of oral administration of artificial cells immobilized phenylalanine ammonia-lyase on intestinal amino acids of phenylketonuric rats. Biomater. Artif. Cells Artif. Organs 17, 161–181 (1989).
Chang, T.M., Bourget, L. & Lister, C. A new theory of enterorecirculation of amino acids and its use for depleting unwanted amino acids using oral enzyme-artificial cells, as in removing phenylalanine in phenylketonuria. Artif. Cells Blood Substit. Immobil. Biotechnol. 23, 1–21 (1995).
Chang, T.M. Therapeutic applications of polymeric artificial cells. Nat. Rev. Drug Discov. 4, 221–235 (2005).
Ruder, W.C., Lu, T. & Collins, J.J. Synthetic biology moving into the clinic. Science 333, 1248–1252 (2011).
Sarkissian, C.N. et al. A different approach to treatment of phenylketonuria: phenylalanine degradation with recombinant phenylalanine ammonia lyase. Proc. Natl. Acad. Sci. USA 96, 2339–2344 (1999).
Durrer, K.E., Allen, M.S. & Hunt von Herbing, I. Genetically engineered probiotic for the treatment of phenylketonuria (PKU); assessment of a novel treatment in vitro and in the PAHenu2 mouse model of PKU. PLoS One 12, e0176286 (2017).
Sonnenborn, U. Escherichia coli strain Nissle 1917-from bench to bedside and back: history of a special Escherichia coli strain with probiotic properties. FEMS Microbiol. Lett. 363, fnw212 (2016).
Schultz, M. Clinical use of E. coli Nissle 1917 in inflammatory bowel disease. Inflamm. Bowel Dis. 14, 1012–1018 (2008).
Joeres-Nguyen-Xuan, T.H., Boehm, S.K., Joeres, L., Schulze, J. & Kruis, W. Survival of the probiotic Escherichia coli Nissle 1917 (EcN) in the gastrointestinal tract given in combination with oral mesalamine to healthy volunteers. Inflamm. Bowel Dis. 16, 256–262 (2010).
Kurtz, C. et al. Translational development of microbiome-based therapeutics: kinetics of E. coli Nissle and engineered strains in humans and nonhuman primates. Clin. Transl. Sci. 11, 200–207 (2018).
Hwang, I.Y. et al. Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models. Nat. Commun. 8, 15028 (2017).
Danino, T. et al. Programmable probiotics for detection of cancer in urine. Sci. Transl. Med. 7, 289ra84 (2015).
Somabhai, C.A., Raghuvanshi, R. & Nareshkumar, G. Genetically engineered Escherichia coli Nissle 1917 synbiotics reduce metabolic effects induced by chronic consumption of dietary fructose. PLoS One 11, e0164860 (2016).
Chen, Z. et al. Incorporation of therapeutically modified bacteria into gut microbiota inhibits obesity. J. Clin. Invest. 124, 3391–3406 (2014).
Palmer, J.D. et al. Engineered probiotic for the inhibition of salmonella via tetrathionate-induced production of Microcin H47. ACS Infect. Dis. 4, 39–45 (2018).
Williams, J.S., Thomas, M. & Clarke, D.J. The gene stlA encodes a phenylalanine ammonia-lyase that is involved in the production of a stilbene antibiotic in Photorhabdus luminescens TT01. Microbiology 151, 2543–2550 (2005).
MacDonald, M.J. & D'Cunha, G.B. A modern view of phenylalanine ammonia lyase. Biochem. Cell Biol. 85, 273–282 (2007).
Cosgriff, A.J. et al. A study of AroP-PheP chimeric proteins and identification of a residue involved in tryptophan transport. J. Bacteriol. 182, 2207–2217 (2000).
Motta, P., Molla, G., Pollegioni, L. & Nardini, M. Structure-function relationships in L-amino acid deaminase, a flavoprotein belonging to a novel class of biotechnologically relevant enzymes. J. Biol. Chem. 291, 10457–10475 (2016).
Hou, Y. et al. Two-Step production of phenylpyruvic acid from L-phenylalanine by growing and resting cells of engineered Escherichia coli: process optimization and kinetics modeling. PLoS One 11, e0166457 (2016).
Hou, Y. et al. Production of phenylpyruvic acid from L-phenylalanine using an L-amino acid deaminase from Proteus mirabilis: comparison of enzymatic and whole-cell biotransformation approaches. Appl. Microbiol. Biotechnol. 99, 8391–8402 (2015).
He, G. et al. Noninvasive measurement of anatomic structure and intraluminal oxygenation in the gastrointestinal tract of living mice with spatial and spectral EPR imaging. Proc. Natl. Acad. Sci. USA 96, 4586–4591 (1999).
Pantaleone, D.P., Geller, A.M. & Taylor, P.P. Purification and characterization of an L-amino acid deaminase used to prepare unnatural amino acids. J. Mol. Catal. 11, 795–803 (2001).
Baek, J.-O., Seo, J.-W., Kwon, O., Seong, Su-Il, Kim, Ik.-H. & Kim, C.H. Heterologous expression and characterization of L-amino acid deaminase from Proteus mirabilis in Escherichia coli. Chin. J. Biotechnol. 24, 21–29 (2008).
Boysen, A., Møller-Jensen, J., Kallipolitis, B., Valentin-Hansen, P. & Overgaard, M. Translational regulation of gene expression by an anaerobically induced small non-coding RNA in Escherichia coli. J. Biol. Chem. 285, 10690–10702 (2010).
Durand, S. & Storz, G. Reprogramming of anaerobic metabolism by the FnrS small RNA. Mol. Microbiol. 75, 1215–1231 (2010).
Unden, G. et al. Control of FNR function of Escherichia coli by O2 and reducing conditions. J. Mol. Microbiol. Biotechnol. 4, 263–268 (2002).
Jones, S.A. et al. Respiration of Escherichia coli in the mouse intestine. Infect. Immun. 75, 4891–4899 (2007).
Wehrmann, A., Phillipp, B., Sahm, H. & Eggeling, L. Different modes of diaminopimelate synthesis and their role in cell wall integrity: a study with Corynebacterium glutamicum. J. Bacteriol. 180, 3159–3165 (1998).
Wright, O., Delmans, M., Stan, G.B. & Ellis, T. GeneGuard: A modular plasmid system designed for biosafety. ACS Synth. Biol. 4, 307–316 (2015).
Mandell, D.J. et al. Biocontainment of genetically modified organisms by synthetic protein design. Nature 518, 55–60 (2015).
Wright, O., Stan, G.B. & Ellis, T. Building-in biosafety for synthetic biology. Microbiology 159, 1221–1235 (2013).
Shedlovsky, A., McDonald, J.D., Symula, D. & Dove, W.F. Mouse models of human phenylketonuria. Genetics 134, 1205–1210 (1993).
Williams, R.A., Mamotte, C.D. & Burnett, J.R. Phenylketonuria: an inborn error of phenylalanine metabolism. Clin. Biochem. Rev. 29, 31–41 (2008).
Caldwell, J., Moffatt, J.R. & Smith, R.L. Post-mortem survival of hippuric acid formation in rat and human cadaver tissue samples. Xenobiotica 6, 275–280 (1976).
Hoskins, J.A. & Gray, J. Phenylalanine ammonia lyase in the management of phenylketonuria: the relationship between ingested cinnamate and urinary hippurate in humans. Res. Commun. Chem. Pathol. Pharmacol. 35, 275–282 (1982).
Layman, D.K. Dietary Guidelines should reflect new understandings about adult protein needs. Nutr. Metab. (Lond.) 6, 12 (2009).
Sawitzke, J.A. et al. Recombineering: in vivo genetic engineering in E. coli, S. enterica, and beyond. Methods Enzymol. 421, 171–199 (2007).
Thomason, L.C., Sawitzke, J.A., Li, X., Costantino, N. & Court, D.L. Recombineering: genetic engineering in bacteria using homologous recombination. Curr. Protoc. Mol. Biol. 106, 1.16.11–1.16.39 (2014).
Miller, J.H. Experiments in Molecular Genetics (Cold Spring Harbor Laboratory Press, 1972).
Acknowledgements
We thank J. Collins, M. Charbonneau, A. Brennan and E. Wolffe for their discussions and comments on the manuscript, and B. Peters for assistance with graphics.
Author information
Authors and Affiliations
Contributions
V.M.I., D.J.L., S.E.R., N.L. and A.B.F. were responsible for strain construction and performance of in vitro experiments. P.J.R. conducted the analysis of in vitro promoter activity. V.M.I. and C.L.A. analyzed the data. B.N.H. performed mouse experiments. M.J.C. and Y.A.M. performed mass spectrometry analysis. M.M.M. and C.G.B. grew cells in a bioreactor. V.M.I., K.A.W., P.F.M., C.K., S.E.G. and D.F. helped supervise the project. All of the authors helped to prepare the manuscript.
Corresponding author
Ethics declarations
Competing interests
Authors hold stock in Synlogic and may gain or lose financially through publication. For information or material, please direct correspondence to vincent@synlogictx.com.
Integrated supplementary information
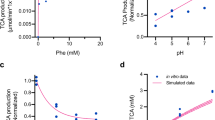
Supplementary Figure 1 PAL and LAAD reaction products in vitro and in vivo.
Enzymatic reactions with Phe and associated metabolites. (a) PAL or LAAD activity results in the formation of trans-cinnamate (TCA) or phenylpyruvate (PP) respectively. (b) The SYNB1618-specific metabolite trans-cinnamate is converted systemically in the host to hippurate, which is excreted and may be measured in urine.
Supplementary Figure 2 Promoter activity of a Pfnrs-lacZ fusion in aerobic and anaerobic conditions in E. coli Nissle.
An E. coli Nissle strain containing a low copy plasmid with a PfnrS-lacZ transcriptional promoter fusionwas grown overnight with vigorous aerobic shaking and back-diluted 1:100 into fresh culture media in a flask with vigorous shaking. At time 0, the culture was split; half remained shaking and the other half was incubated statically in an anaerobic chamber. At the indicated time points, 1mL of culture was removed and promoter activity was determined by assessing β-galactosidase levels. Anaerobic induction was observed. Data is shown for 2 independent replicate cultures.
Supplementary Figure 3 Induction of PfnrS-GFP promoter activity in a bioreactor model
An E. coli Nissle strain containing a low copy plasmid with a PfnrS-GFP transcriptional promoter fusion was grown overnight and then used to inoculate 2 mL wells of a BioLector microbioreactor system with the volume of overnight culture indicated (2 replicates each of 10, 100, 150, and 200μL inoculum and 1 replicate of 50μL inoculum). Oxygen transfer rate remained constant over time. Dissolved oxygen content (solid lines) in the culture media was shown to decrease over time with culture growth, with lower inocula resulting in slower consumption of oxygen. Increased GFP detection (right y-axis, dotted lines), and thus PfnrS-mediated expression, was associated with decreasing dissolved oxygen content of the culture media, consistent with PfnrS promoter activation under oxygen-limitation.
Supplementary Figure 4 Analysis of the PfnrS promoter in vivo.
Activation of the PfnrS promoter was measured in vivo. (a) Initial in vitro profiling of the Pfnrs promoter was performed by flow cytometry. Aerobic (+O2, left histogram) and anaerobic (-O2, right histogram) cultures were analyzed with a negative control strain of EcN without GFP (red), a positive control strain expressing GFP constitutively (blue), and a strain expressing GFP under control of the PfnrS promoter (green). As expected, the negative control showed no detectable GFP signal in either aerobic or anaerobic conditions, the positive control strain showed equivalent GFP expression in either aerobic or anaerobic conditions, and the PfnrS-GFP fusion strain only expressed GFP under anaerobic conditions (Note: While the constitutive control was expressed from the chromosome, PfnrS-GFP was expressed from a plasmid, which may have led to more variable and higher level expression observed in the historgrams). This experiment was repeated independently with similar results (b) The aerobically grown strains analyzed in (a) were dosed orally to C57BL/6 mice and recovered 4h post-dose by flushing excised cecal tissue with PBS As demonstrated in (a), the PfnrS-GFP fusion strain was off at the time of dosing. Cecal effluents were analyzed by flow cytometry. For each plot, 40,000 counted events are shown. The y-axis represents the SSC and the x-axis the GFP channel. Plot 1 shows analysis of the background and autofluorescence of cecal effluent in a mouse that did not receive cells. Plot 2 shows analysis of cecal effluent from a mouse dosed with control EcN (no GFP), and no fluorescence is observed. Plot 3 shows analysis of cecal effluent after dosing with EcN expressing GFP constitutively, and a GFP fluorescent population is observed and gated on. This gate was applied to all plots. Plot 4 shows analysis of cecal effluent after dosing with EcN containing a PfnrS-GFP fusion that were OFF at the time of dosing. GFP fluorescence was observed in cecal effluent (increased abundance of events counted in the GFP-positive gate). When normalized to CFU counts recovered from the effluent and by subtracting cecal effluent background (Plot 1), the gated populations account for ~76 % (constitutive GFP control) and ~64 % (PfnrS-GFP) of the cells recovered by plating. Though considerable variability exists in CFU counting, especially from biological samples, the conclusion is that a majority of the PfnrS-controlled cells appear to enter the “ON” state post-dosing. This experiment was repeated independently with similar results
Supplementary Figure 5 Effect of dapA deletion on SYNB1618 growth in vitro
To characterize the growth of E. coli Nissle (EcN) and therapeutic candidate SYNB1618, which contains a mutation in the dapA gene, both strains were incubated in LB that did (+) or did not (-) contain diaminopimelic acid (DAP; 100 μg/mL) at 37 °C for 960 minutes under constant shaking. The OD600 was measured every 10 minutes to assess cell growth over time. The average OD ± standard deviation of 6 replicate cultures is plotted for each time point. Data shows that SYNB1618 is unable to grow without the addition of exogenous DAP to the growth media.
Supplementary Figure 6 Enterorecirculation of Phe
Proposed mechanism of enterorecirculation of Phenylalanine. Proteins are continuously replenished in the intestinal lumen though the introduction and digestion of dietary protein and from glandular secretions as well as through turnover of the intestinal epithelial cells. Intestinal proteolysis generates a pool of Phe containing peptides and free Phe which may then be reabsorbed back into the body as they pass down the intestine. Peptides and free Phe may be converted into proteins systemically which may be reintroduced into the intestine through glandular secretions and amino acid reabsorption (see Chang and Lister, 1995).
Supplementary Figure 7 In vivo PAL activity in engineered strains is increased upon co-expression with pheP
ENU2 mice on Phe-deficient diet were housed in metabolic cages and orally gavaged with 3 × 1010 flask-grown cells of E. coli Nissle strains expressing the gene encoding PAL from a plasmid, with or without co-expression of a chromosomally integrated pheP gene (Strains PAL or PAL/PheP respectively, Supplementary Table 3, n = 9) Each dot represents a metabolic cage of 3 mice/cage. Urine was collected for 4h and analyzed for HA content by LC-MS/MS. Bars represent the average urinary HA recovery of the 3 cages.
Supplementary Figure 8 Conversion efficiency of oral trans-cinnamate to urinary hippurate in non-human primates
NHPs (n = 6) were orally administered 13C-trans-cinnamate (13C-TCA) and urine was collected over 6h. Each bar represents a single NHP subject. 13C-Hippurate (13C-HA) was measured in the urine by mass spectroscopy. The percentage of urinary 13C-HA recovered as a function of 13C-TCA administered was calculated and used as a normalization factor for HA recovery in subsequent experiments. This factor accounts for TCA that is not converted to HA or that is lost to incomplete urinary collection, thus allowing a more accurate description of strain activity.
Supplementary Figure 9 SYNB1618-specific metabolite detection in serum of non-human primates
Using LC-MS/MS, serum concentrations of d5-HA (a) and d5-TCA (b) were determined in non-human primates administered d5-Phe and SYNB1618 orally. No detectable d5-HA or d-TCA was detected when d5-Phe was administered in the absence of SYNB1618 (data not shown). The presence of these metabolites demonstrates SYNB1618-specific activity in these animals.
Supplementary Figure 10 Schematic of the SYNB1618 genome
Schematic of clinical candidate SYNB1618. SYNB1618 contains chromosomally inserted genes encoding PheP, a high affinity phenylalanine (Phe) transporter that can bring Phe into the cell, PAL (stlA), which converts Phe into trans-cinnamic acid (TCA), and LAAD (pma), which converts Phe to phenylpyruvate (PP). Regulation of these components is carried out by anaerobic- IPTG-, and L-arabinose-inducible promoters, for activation in the mammalian gut or in vitro. The locations of the genomic modification sites in SYNB1618 are shown, with kbp designation indicating the chromosomal position relative to the 0/5.4 Mb reference marker. The chromosomal origin of replication is shown as a red line. Italicized gene names in parenthesis refer to the upstream and downstream genes surrounding the inserted gene.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–10 (PDF 796 kb)
Supplementary Tables
Supplementary Tables 1–5 (PDF 397 kb)
Rights and permissions
About this article

Cite this article
Isabella, V., Ha, B., Castillo, M. et al. Development of a synthetic live bacterial therapeutic for the human metabolic disease phenylketonuria. Nat Biotechnol 36, 857–864 (2018). https://doi.org/10.1038/nbt.4222
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nbt.4222
This article is cited by
-
Enhancing tumor-specific recognition of programmable synthetic bacterial consortium for precision therapy of colorectal cancer
npj Biofilms and Microbiomes (2024)
-
A programmable protease-based protein secretion platform for therapeutic applications
Nature Chemical Biology (2024)
-
Delivery of a sebum modulator by an engineered skin microbe in mice
Nature Biotechnology (2024)
-
Cas9-assisted biological containment of a genetically engineered human commensal bacterium and genetic elements
Nature Communications (2024)
-
Advanced probiotics: bioengineering and their therapeutic application
Molecular Biology Reports (2024)
