
Increased expression of sirtuin proteins has been shown to enhance lifespan in several organisms. New data indicate that some of the reported effects may have been due to confounding factors in experimental design. Here, experts discuss the significance of these data for research into ageing. See Letter p.482
The paper in brief
-
Previous work1,2,3,4 had shown that increased expression of the yeast protein Sir2 and of related sirtuin proteins in the nematode Caenorhabditis elegans and the fruitfly Drosophila melanogaster extends lifespan.
-
Burnett et al.5 believed that the organisms' genetic backgrounds, because they had not been controlled, might have affected those results, and so they repeated the experiments after standardizing the backgrounds.
-
By means of genetic backcrossing (Fig. 1), the authors show that sirtuin overexpression and longevity in C. elegans are separable.
Figure 1: Eliminating effects of confounding genetic sequences. 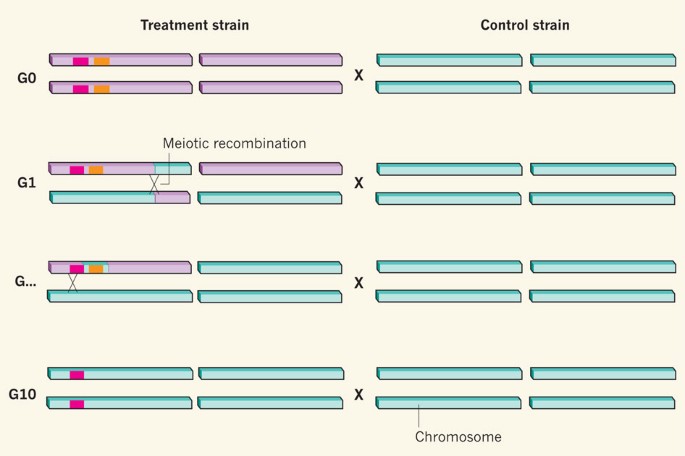
Burnett et al.5 show that genetic backcrossing is essential to ensure that comparisons between treatment and control strains of an organism are not confounded by differences unrelated to the gene of interest (pink). Confounding effects may be due to a large effect of a nearby gene (orange) or small effects of many genes throughout the genome (occurring within the purple elements). At generation 0 (G0, no backcrossing), these influences predominate. With backcrossing, processes such as independent chromosome segregation and meiotic recombination introduce variants from the control strain (blue elements) into the treatment strain, reducing the possibility of confounding effects. Although most differences between the two strains are rapidly lost (at a rate of roughly 50% per generation), closely linked differences often persist for many generations. It is only after recombination occurs sufficiently closely to the gene of interest (at G10 or so) that genetic backgrounds can be considered properly controlled and that differences can be attributed to the gene of interest.
-
Two Drosophila strains that had been reported to be long-lived as a result of sirtuin overexpression were not long-lived when compared with a genetically appropriate control strain.
-
The study refutes yet another previous observation — that the increase in fly lifespan that occurs with dietary restriction depends on sirtuins.
-
In an accompanying Brief Communication Arising, Viswanathan and Guarente6 show that sirtuin overexpression leads to a smaller increase in C. elegans lifespan than previously reported.

A valuable background check
David B. Lombard & Scott D. Pletcher
Burnett and colleagues' in-depth study5 of the influence of Sir2 overexpression on lifespan in C. elegans and Drosophila, as well as on diet-modulated longevity in the fly, shows that the protein has no effect, challenging previous publications1,2,4. But since when does Nature publish negative results?
Well, sirtuins are exceptional. Anything that can influence ageing captures the public's imagination, and — since the reports that sirtuins are involved in increasing lifespan in invertebrates — research into these proteins has exploded. Moreover, the seven mammalian sirtuins have been implicated in the suppression of numerous age-associated diseases, including neurodegenerative disorders, cardiac dysfunction, hearing loss and neoplasia7.
Meanwhile, heated controversies have arisen regarding aspects of sirtuin biology. These involve the proteins' roles in the response to dietary restriction; the mechanism of action of reported sirtuin activators; and whether the role of Sir2 in promoting longevity — first identified in yeast — is evolutionarily conserved in other organisms. The disagreements fostered complaints of publication bias and disproportionate scientific and commercial emphasis on the role of sirtuins in longevity. So, here we are.
Burnett et al.5 were thorough. They generated an impressive collection of lifespan experiments, and these were replicated and the results contributed by several laboratories. The data were supported by backcrossing and by genetic and biochemical analyses. But an accompanying report from Viswanathan and Guarente6, although confirming some of Burnett and co-workers' findings, shows that worms overexpressing Sir2 retain modestly increased (10–13%) lifespan after backcrossing — consistent with data reported elsewhere8.
At best, therefore, these papers indicate that Sir2 overexpression is just one of more than 100 genetic manipulations currently known to increase worm and/or fly lifespan to some degree, with many others having larger effects.
Challenging published results is an essential, self-correcting aspect of science.
Given the demonstrated importance of sirtuins in mammals, why rehash the precise role of Sir2 in worm and fly ageing? First, challenging published results is an essential, self-correcting aspect of science. Second, invertebrate models continue to contribute to the understanding of sirtuin biology in mammals, and so future studies must be interpreted in the context of these new data. Third, the new reports5,6 reinforce the importance of rigorous genetic background control when interpreting the effects of single gene mutations. Studies in Drosophila suggest that 8–10 generations of backcrossing are required before the genetic background can be considered reasonably well controlled; this needs to be the standard in worms too (Fig. 1). For genetically engineered mutant mice, where this approach may be impractical, analysis of independent transgenic animal lines and/or reversal of traits upon re-expression of the gene of interest should be undertaken to ensure that that gene is indeed responsible for the effect under investigation.
Perhaps the most interesting issue is the apparent disconnect between these new results and the powerful effects of sirtuins on age-associated disease in mammals. Maybe sirtuins have their strongest effects on specific aspects of physiological homeostasis and stress responses, rather than in modulating ageing per se. For example, consistent with published data9, Burnett et al.5 show that Sir2 overexpression makes worms resistant to toxic protein aggregates. Also, increased expression of the related SIRT1 protein in mice suppresses metabolic dysfunction and the development of certain types of cancer, without increasing overall lifespan10. Although roles for other mammalian sirtuins in promoting longevity have not been explored, such studies are under way in several laboratories. So the latest reports5,6 are unlikely to be the final word on sirtuins and ageing.
Don't write sirtuins off
Carles Cantó & Johan Auwerx
The role of Sir2 and its mammalian counterpart SIRT1 as lifespan regulators, proposed on the basis of work done exclusively in simple organisms, has been grounds for intensive discussion. Studies on yeast lifespan were the first to cast doubt on the role of sirtuins in longevity11. And Burnett and colleagues' elegant work5 puts a final nail in the coffin. Let's not forget, however, that an overwhelming body of evidence indicates that sirtuins have crucial roles in metabolic homeostasis.
One caveat of most of the genetic work in simple organisms is that it is rarely accompanied by biochemical studies.
The new paper, indeed, dampens the longevity claims assigned to Sir2. But one caveat of most of the genetic work in simple organisms is that it is rarely accompanied by biochemical studies that reveal the extent to which the manipulations of a gene or protein are translated into altered activity. Sirtuins are protein deacetylase enzymes, and so in this case it remains unclear how manipulation of the gene encoding Sir2 changes the acetylation state of its main target proteins. Such information would establish beyond doubt that the absence of the Sir2 gene is not compensated for by increased activity of another member of the sirtuin family.
As for the relevance of these data5,6 to the role of sirtuins in mammals, it was previously shown that mice overexpressing SIRT1 do not live longer12. Initially, this lack of effect on lifespan was attributed either to insufficient expression of the SIRT1 transgene or to possible confounding functional compensation of protein deacetylation by other Sir2-related proteins. However, if Burnett and colleagues' results can be extended to mice, then the earlier mouse data12 are not so astonishing after all. The main role of SIRT1 in mammals might not be directly related to lifespan regulation.
If not to control lifespan, what is the main function of SIRT1? The answer may lie in the observation that the activity of SIRT1 depends strictly on the levels of the coenzyme NAD+, which acts as a co-substrate for the deacetylation reaction that SIRT1 catalyses. Changes in NAD+ levels, which reflect the metabolic activity of the cell, hence modulate SIRT1 activity according to the cellular energy status. This NAD+ dependence makes SIRT1 an attractive integrative node for metabolic sensing and for transcriptional regulation, as SIRT-1 influences post-translational modification of transcription factors and histone proteins3.
In contrast to the observations on lifespan, the effects of SIRT1-related proteins favouring metabolic flexibility are based on a wealth of genetic, physiological and pharmacological evidence, and are conserved from yeast to mammals. In mammals, SIRT1 mediates metabolic and transcriptional adaptations to situations of energy stress and nutrient deprivation by enhancing respiration by the mitochondria, the cell's energy producers. Mice overexpressing SIRT1 are therefore protected from the metabolic damage caused by a high-fat diet10.
Similarly, indirect activation of SIRT1 by the compound resveratrol protects against metabolic and age-related diseases13, curbing the lifespan reduction induced by high-calorie diets14, even though it has no effect on lifespan in mice fed regular chow15. Conversely, outbred mice lacking SIRT1 show deficiencies in metabolism and cannot respond properly to the lifespan-increasing effects of calorie restriction12. This highlights how the metabolic adaptations that SIRT1 induces might indirectly influence mammalian lifespan.
Indeed, although in light of Burnett and colleagues' findings5 the appeal of sirtuins as a sensu stricto lifespan determinant might be gone, SIRT1 activation remains a promising approach to delaying general age-related physiological decline and to treating numerous inherited and acquired diseases characterized by defective mitochondrial function. The astonishing ability of SIRT1 to enhance 'healthspan' by promoting metabolic fitness will also guarantee it a long life as a subject for further exciting research.
References
Tissenbaum, H. A. & Guarente, L. Nature 410, 227–230 (2001).
Rogina, B. & Helfand, S. L. Proc. Natl Acad. Sci. USA 101, 15998–16003 (2004).
Cantó, C. & Auwerx, J. Trends Endocrinol. Metab. 20, 325–331 (2009).
Bauer, J. H. et al. Aging 1, 38–48 (2009).
Burnett, C. et al. Nature 477, 482–485 (2011).
Viswanathan, M. & Guarente, L. Nature 10.1038/nature10440 (2011).
Guarente, L. N. Engl. J. Med. 364, 2235–2244 (2011).
Rizki, G. et al. PLoS Genet. 7, e1002235 (2011).
Parker, J. A. et al. Nature Genet. 37, 349–350 (2005).
Herranz, D. et al. Nature Commun. 1, 3 (2010).
Kaeberlein, M. & Powers, R. W. III Ageing Res. Rev. 6, 128–140 (2007).
Herranz, D. & Serrano, M. Nature Rev. Cancer 10, 819–823 (2010).
Lagouge, M. et al. Cell 127, 1109–1122 (2006).
Baur, J. A. et al. Nature 444, 337–342 (2006).
Pearson, K. J. et al. Cell Metab. 8, 157–168 (2008).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Lombard, D., Pletcher, S., Cantó, C. et al. Longevity hits a roadblock. Nature 477, 410–411 (2011). https://doi.org/10.1038/477410a
Published:
Issue Date:
DOI: https://doi.org/10.1038/477410a
This article is cited by
-
Balancing NAD+ deficits with nicotinamide riboside: therapeutic possibilities and limitations
Cellular and Molecular Life Sciences (2022)
-
Alternsumkehr und Lebensspannenverlängerung
Zeitschrift für Gerontologie und Geriatrie (2013)
-
2011 Editors' choice
Nature (2011)
-
A midlife crisis for sirtuins
Nature Reviews Molecular Cell Biology (2011)
