
Abstract
Poplar has 192 annotated R2R3 MYB genes, of which only three have been shown to play a role in the regulation of secondary cell wall formation. Here we report the characterization of PtrMYB152, a poplar homolog of the Arabidopsis R2R3 MYB transcription factor AtMYB43, in the regulation of secondary cell wall biosynthesis. The expression of PtrMYB152 in secondary xylem is about 18 times of that in phloem. When expressed in Arabidopsis under the control of either 35S or PtrCesA8 promoters, PtrMYB152 increased secondary cell wall thickness, which is likely caused by increased lignification. Accordingly, elevated expression of genes encoding sets of enzymes in secondary wall biosynthesis were observed in transgenic plants expressing PtrMYB152. Arabidopsis protoplast transfection assays suggested that PtrMYB152 functions as a transcriptional activator. Taken together, our results suggest that PtrMYB152 may be part of a regulatory network activating expression of discrete sets of secondary cell wall biosynthesis genes.
Similar content being viewed by others


Introduction
In Arabidopsis thaliana (Arabidopsis), there are nearly 200 genes encoding MYB transcription factors1, which are classified according to the number of N-terminal DNA binding domain repeats. R2R3 MYB proteins containing two N-terminal DNA binding domain repeats are the largest MYB transcription factor subfamily with 126 members1,2. R2R3 MYB transcription factors control several aspects of plant growth and development. For example, GLABRA1 (GL1) and WEREWOLF (WER) are involved in determining cell fate during trichome and root hair cell differentiation, respectively3,4, while AtMYB77 regulates lateral root formation5 and ASYMMETRIC LEAVES1 (AS1) regulates shoot morphogenesis and leaf patterning6. Recently, several R2R3 MYB transcription factors have been found to regulate secondary cell wall biosynthesis in Arabidopsis7,8,9. Similar findings were observed in Populus spp. (poplar) and Eucalyptus spp. (Eucalyptus)10,11,12.
Unlike primary cell walls, which are synthesized at the cell plate when cells divide and during cell expansion in growing cells, secondary cell walls are deposited in defined cell types such as tracheary elements and fibers after cell expansion has ceased. The massive deposition of lignin, cellulose and hemicelluloses inside primary walls gives secondary walls their characteristic thickness and strength13. Genetic analyses using the Arabidopsis inflorescence stems, roots and in vitro secondary cell walls induced in cell culture, have identified a transcription factor network that regulates secondary cell wall biosynthesis7,14,15. In the network, several closely related NAC domain transcription factors, including SECONDARY WALL-ASSOCIATED NAC DOMAIN PROTEIN1 (SND1), NAC SECONDARY WALL THICKENING PROMOTING FACTOR1 (NST1), NST2, VASCULAR-RELATED NAC DOMAIN6 (VDN6) and VND7 have been identified as master regulators that are capable of modulating the entire biosynthetic pathways of the secondary wall components cellulose, xylan and lignin7,14,16,17,18. These NAC domain transcription factors can directly activate the expression of secondary wall specific biosynthetic genes7,19,20 and activate the expression of several downstream transcription factor genes that also directly regulate secondary wall component biosynthetic genes7,20.
Among the identified downstream transcription factor genes, two encode NAC domain transcription factors (SND2 and SND3) and one encodes the KNOTTED ARABIDOPSIS THALIANA7 (KNAT7) KNOTTED1-like homeodomain (KNOX) transcription factor, which has recently been shown to negatively regulate secondary cell wall biosynthesis via interaction with OVATE FAMILY PROTEIN4 (OFP4)20, a transcription factor from a newly identified transcription repressor family21. All other downstream transcription factors identified thus far encode for R2R3 MYB transcription factors, including AtMYB46, AtMYB52, AtMYB54, AtMYB58, AtMYB63, AtMYB85 and AtMYB1037,16,19. Among these, AtMYB46, AtMYB58, AtMYB63, AtMYB83 and AtMYB103 have all been shown to be the direct targets of the master regulators SND1/VND6/VND77,18,19,22. AtMYB20, AtMYB42 and AtMYB43 have also been shown to be regulated by SND17, but their functions in secondary cell wall biosynthesis have not been characterized. Ectopic expression of AtMYB46 alone has been shown to be sufficient to induce the entire secondary cell wall biosynthesis program, while AtMYB58 and AtMYB63 specifically activate lignin biosynthesis genes during secondary cell wall formation. AtMYB75 has also been shown to regulate secondary cell wall biosynthesis by interacting with KNAT78,9, though it is unclear if its expression is regulated by NAC domain master switch transcription factors.
Populus trichocarpa is a good model system for studying wood development, perenniality, phenology and ecological interactions, processes that cannot be truly evaluated in annual model plants systems, such as Arabidopsis23,24. With growing interests in the use of lignocellulose as a source of biomass for bioenergy, insight into the control and regulation of secondary cell wall biosynthesis will help guide genetic improvement strategies for energy crops, such as Populus25.
In Populus, there are 192 annotated genes encoding R2R3 MYB transcription factors26. To date, several lines of evidence support the involvement of R2R3 MYB transcription factors in the regulation of secondary cell walls biosynthesis in poplar. First, several poplar MYB genes have been shown to be highly expressed during secondary vascular tissue formation27,28. Second, PttMYB21, an ortholog of Arabidopsis AtMYB46 has been shown to be expressed primarily in xylem tissues12. Third, Populustrichocarpa PtrMYB03 and PtrMYB20, PtrMYB21 paralogs, were also shown to be orthologous to Arabidopsis AtMYB46 and its paralog AtMYB83 that can activate the biosynthetic pathways for cellulose, xylan and lignin when overexpressed in Arabidopsis11. Finally, overexpression of PtrWND2B, a NAC domain master switch transcription factor in poplar induced the expression of several poplar R2R3 MYB genes including PtrMYB03, PtrMYB018, PtrMYB20, PtrMYB21, PtrMYB028 and PtrMYB19229. However, the expression of PtrMYB152 was not affected in transgenic poplar overexpressing PtrWND2B. Since PtrMYB152 and its paralog PtrMYB018 are orthologous to Arabidopsis AtMYB43 and its paralog AtMYB2029, it would be of great interest to determine if PtrMYB152 is involved in the regulation of secondary cell wall biosynthesis.
Results
PtrMYB152 is a poplar homologue of Arabidopsis R2R3 MYB transcription factor AtMYB43
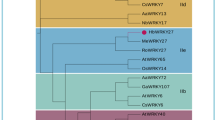
The entire amino acid sequences of each of the five Arabidopsis R2R3 MYB proteins AtMYB20, AtMYB43, AtMYB46, AtMYB58 and AtMYB63 were used in BLAST searches of the poplar protein database (www.phytozome.net), to identify the most closely related poplar MYBs for each of them. Based on their predicted amino acid sequence similarities to their Arabidopsis homologues, poplar genes encoding 5 R2R3 MYBs including PtrMYB018, PtrMYB021, PtrMYB028, PtrMYB152 and PtrMYB192 were identified. PtrMYB018 was identified as the most closely related poplar MYB to AtMYB20, however, when using the amino acid sequence of the identified poplar MYBs to do BLAST, we found AtMYB43 is the Arabidopsis MYB with highest amino acid sequence similarities to PtrMYB018, similar to the results obtained by Wilkins et al26, thus AtMYB20 was excluded from phylogenetic analysis. As shown in Figure 1A, PtrMYB018 and PtrMYB152 are paralogs and related to AtMYB43; PtrMYB028 and PtrMYB192 are paralogs closely related to AtMYB58 and AtMYB63 and PtrMYB021 is closely related to AtMYB4611,12,26.

PtrMYB152 is a homolog of Arabidopsis R2R3 MYB transfection factor AtMYB43.
(A) Phylogenetic analysis of poplar homologues of Arabidopsis R2R3 MYB transcription factors. All bootstrap values were >80 (out of 100 replicates) (B) Relative expression levels of PtrMYB152 in poplar xylem and phloem by real time RT-PCR analysis. Expression of C672 was used as a reference to normalize the expression of poplar genes and expression level of PtrMYB152 in phloem was set as 1.
PtrMYB152 is highly expressed in xylem and encodes a transcriptional activator
Among the 5 poplar R2R3 MYB genes, all but PtrMYB152 have been shown to be induced by NAC domain master switch transcription factor PtrWND2B in poplar29. So we decided to further investigate if PtrMYB152 is involved in secondary cell wall formation. We first compared its expression in poplar cells undergoing secondary wall development and in developing phloem by quantitative RT-PCR (qRT-PCR). The results showed that PtrMYB152 is highly expressed in secondary xylem, i.e., about 18 times higher than in phloem (Figure 1B).
To determine if PtrMYB152 functions as transcriptional activator or repressor, we employed a protoplast transfection system. PtrMYB152 was fused to an N-terminal GAL4 DNA binding domain (GD) and plasmids harboring this fusion or the GD domain alone as a control were co-transfected with a plasmid containing the GAL4-GUS reporter gene into protoplasts. Co-transfection with GD alone had little effect on the expression of the reporter gene, while co-transfection with the GD-PtrMYB152 fusion resulted in the activation of the reporter gene (Figure 2). This implies that PtrMYB152 functions as a transcriptional activator.

PtrMYB152 is transcriptional activator.
A protoplast transfection system was used to assay the ability of PtrMYB152 to activate the GUS reporter gene. Effectors and reporter (diagrammed on the top of the figure) plasmids were co-transfected into protoplasts isolated from Arabidopsis rosette leaves. GUS activity was assayed after protoplasts had been incubated in darkness for 20–22 h.
Tissue specific expression of the PtrCesA8 promoter in Arabidopsis
The finding that the PtrMYB152 functions as a transcription activator (Figure 2) and is highly expressed in poplar xylem (Figure 1B) prompted us to further investigate its potential functions in secondary cell wall biosynthesis by heterologously expressing PtrMYB152 gene in Arabidopsis wild-type plants. We employed the commonly used 35S promoter to ectopically express the genes at high levels. In addition, we employed the poplar CesA8 promoter30, from the poplar homolog of the Arabidopsis CesA8 gene associated with secondary cell wall biosynthesis31 for its ability to specifically direct expression to developing xylem of secondary walls in Arabidopsis. Since PtrCesA8 enzyme function is required for the biosynthesis of cellulose in secondary walls and it is specifically expressed in cells undergoing secondary wall thickening in poplar, expression of PtrMYB152 under the control of PtrCesA8 will be activated in cells undergoing secondary wall thickening and may work in a feedback mechanism to constitutively active secondary cell wall biosynthesis genes.
The PtrCesA8 promoter (~3000 bp PtrCesA8 promoter fragment) was first shown to be fully active by fusing it to the GUS reporter gene to generate PtrCesA8prom:GUS and GUS activity was examined histochemically in transgenic Arabidopsis plants. Expression patterns were similar among five individual transgenic lines examined. At the seedling stage, GUS activity was mainly detected in the veins of cotyledons, rosette leaves and hypocotyls and in the stele of roots (Figure 3A–E). In the inflorescence stem, GUS activity was primarily observed in the vascular bundles and interfascicular fibers (Figure 3F, G), suggesting that the promoter could be used to direct PtrMYB152 expression preferentially to developing xylem.

Expression of PtrCesA8prom:GUS in Arabidopsis transgenic plants.
5-bromo-4-chloro-3-indolyl-β-D-glucuronide (X-Gluc) was used as a substrate for histochemical staining for GUS activity. (A) whole seedling, (B) close-up views of shoot, (C) cotyledon, (D) hypocotyls, (E) root tip, (F) upper stem section and (G) lower stem section.
Heterologous expression of PtrMYB152 affects secondary cell wall development
Constructs of PtrMYB52 with an HA tag at its N-terminus under the control of the 35S and PtrCesA8 promoters were made and transformed into Arabidopsis. Multiple lines for each construct were obtained and four lines analyzed in the T2 to T4 generations for each construct. Phenotypes observed were consistent in the four independent lines.
Transgenic lines expressing PtrMYB152 were assessed for variation in secondary cell wall thickening and chemical content. Cross sections were taken from the bases of the inflorescence stems from transgenic plants and control wild-type (Col-0) plants at principal growth stage 6.1032 when the inflorescence stems of the plants were at similar height of ~12 cm with two to three fully expanded siliques. Preliminary observation of secondary cell wall thickness and morphology using cross-sections stained with toluidine blue showed that heterologous expression of PtrMYB152 under the control of either the 35S promoter or the PtrCesA8 promoter had interfascicular fibers with significantly thicker cell walls than wild-type Col-0 (data not shown). A more detailed view of the effects of PtrMYB152 on secondary cell wall biosynthesis was obtained by examining sections using transmission electron microscopy (TEM). This analysis revealed that all vessel, interfascicular fiber and xylary fiber secondary cell walls in the transgenic plants were thicker than those of wild type (Figure 4, Table 1).

Secondary cell wall thickening in Arabidopsis transgenic plants expressing PtrMYB152.
Cross-sections from basal inflorescence stems of wild type and transgenic plants at principal growth stage 6.10.If, interfascicular fiber; ve vessel; xf, xylary fiber; p, phloem. Bars, 10 μm.
Cross-sections were also stained with phloroglucinol to visualize lignin deposition. As shown in Figure 5, more intense phloroglucinol staining was observed in sections from the transgenic lines relative to wild-type, suggesting an increase in lignin content in the transgenic plants.

Secondary cell wall lignification in Arabidopsis transgenic plants expressing PtrMYB152.
Phloroglucinol stained sections from basal inflorescence stems of wild type and transgenic plants at principal growth stage 6.10. Bars, 50 μm.
Cell wall chemistry of PtrMYB152 expressing lines
Since observations from phloroglucinol-stained stem cross sections suggested an increase in lignin content in transgenic plants expressing PtrMYB152, cell wall chemistry was assessed in inflorescence stems of transgenic plants relative to wild-type plants. Because chemical analysis requires relative large amounts of dry samples, pools of stems from three independent homozygous lines expressing each construct were used for chemistry analysis.
As shown in Table 2, total lignin content in the transgenic plants was elevated, with an 24% and 18% increase for 35:PtrMYB152 and PtrCesA8:PtrMYB152 relative to wild-type respectively. We further examined the relative amounts of syringyl (S) and guaiacyl (G) monomers in the inflorescence stem cell walls, which showed that the S/G monomer ratio was higher in the transgenic plants than wild-type plants (Table 3). The significantly increased interfascicular fiber cells wall thicknesses of the transgenic lines may explain this alteration, since such cells develop the bulk of secondary walls in the Arabidopsis inflorescence stem and fiber cells are enriched in S-lignin33.
PtrMYB152 regulates secondary cell wall biosynthetic gene expression
To test the effect of heterologous expression of PtrMYB152 on potential target genes, we examined the expression of a suite of secondary cell wall biosynthetic genes in Arabidopsis PtrMYB152 expressing lines that exhibited altered cell wall properties. The analyses were carried out using seedlings, pooled from three independent lines for each construct, to ensure tissue of uniform age for each genotype. Using seeding rather the stems also allowed us to identify genes that may be directly activated by PtrMYB152. As shown in Figure 6, the transgenic lines had increased expression of several lignin biosynthetic genes including 4CL1 (At1g51680), HCT (At5g48930), C3H1 (At2g40890), CCoAOMT1 (4g34050) and CAD6 (Atg37970). This suggests that PtrMYB152 regulate secondary cell wall biosynthesis by activating a subset of secondary cell wall biosynthetic genes.

Lignin biosynthetic genes expression in transgenic plants.
RNA was isolated from 7-day old seedlings of transgenic and wild-type plants. qRT-PCR was used to detect the expression of related genes. Expression of ACTIN2 was used as a control reference gene. Expression of each gene in wild type was set as 1.
Discussion
In Arabidopsis, secondary cell wall biosynthesis is controlled by a transcription factor network. Several closely related NAC domain transcription factors, including SND1, NST1, NST2, VDN6 and VND7 work together to activate several downstream transcription factor genes and the downstream transcription factors directly activate secondary cell wall components biosynthesis genes7,14,15,17. Most of the downstream transcription factors identified so far are R2R3 MYB transcription factors, which play important roles in regulating secondary cell wall biosynthesis7,15. In poplar, several R2R3 MYB transcription factors have also been shown to be involved in the regulation of secondary cell wall biosynthesis11,12,27,28,29.
In this report, we provide evidence showing that PtrMYB152 is involved in the regulation of secondary cell wall biosynthesis. In poplar, there are a total of 192 genes encoding R2R3 MYB transcription factors26, however, their functions in plant growth and development remain largely unknown. By searching for poplar homologues of Arabidopsis R2R3 MYBs that are known to be involved in the regulation of secondary cell wall biosynthesis, we identified five poplar MYBs including PtrMYB018, PtrMYB021, PtrMYB028, PtrMYB152 and PtrMYB192 (Figure 1A). Among them, PtrMYB018, PtrMYB021, PtrMYB028 and PtrMYB192 have been shown to be regulated by PtrWND2B and involved in the regulation of secondary cell wall biosynthesis, however, the expression of PtrMYB152 was not affected in PtrWND2B transgenic poplar28. Real-time RT-PCR results showed that PtrMYB152 is differentially expressed in developing xylem and phloem in poplar and its expression level in secondary xylem is about 18-fold great than in phloem (Figure 1B), indicating PtrMYB152 may be involved in the regulation of secondary cell wall formation. Consistent with this hypothesis, phenotypic analysis showed that overexpression of PtrMYB152 increased secondary cell wall thickness of vessel and fiber cells in Arabidopsis inflorescence stems (Figure 4, Table 1). Chemical analysis suggested an increase in lignification and other changes in cell wall composition in the stems of transgenic plants (Figure 5, Table 2, Table 3). These results demonstrate that PtrMYB152 regulates secondary cell wall biosynthesis in Arabidopsis.
When recruited to the promoter region of the Gal4-GUS reporter gene via a fused GD domain in a protoplast transient express system, PtrMYB152 activated GUS expression, indicating that it is a transcription activator (Figure 2). Consistent with this observation, real-time RT-PCR showed that some genes required for the biosynthesis of major secondary cell wall components cellulose, xylan and lignin are activated by overexpression of PtrMYB152 (Figure 6), indicating that PtrMYB152 controls secondary cell biosynthesis by inducing the expression of discrete sets of secondary cell wall biosynthesis genes.
Secondary cell walls are formed by deposition of cell wall polymers mainly lignin, cellulose and xylan after cell expansion has ceased13,34 and this process is regulated by a transcription factor regulatory network7,15. Because spatially and temporally expression of related transcription factor genes may be important for their proper functions, in addition to the 35S promoter, we also used a tissue specific PtrCesA8 promoter (Figure 3) to drive the expression of PtrMYB152. We found that overexpression of PtrMYB152 by either the PtrCesA8 promoter or the 35S promoter resulted in similar but slightly different effects on secondary cell wall thickening in Arabidopsis (Figure 4). These data suggest that spatially and temporally regulated expression may play a role in the proposed functions of PtrMYB152.
Our data on the poplar MYB transcription factor PtrMYB152, combined with other recent studies of poplar genes encoding orthologs of Arabidopsis MYB, NAC domain and homeodomain proteins in the secondary cell wall regulatory network support the idea that the transcription regulatory network governing secondary cell wall biosynthesis is largely conserved in poplar and Arabidopsis. Figure 7 summarizes these data, which indicate that for each of the known Arabidopsis transcriptional regulators, poplar orthologs with the similar functions have been identified. These transcription factors provide new tools for understanding and manipulating wood formation in poplar and other tree species. It should be noted that by examining four in-house secondary cell wall formation related transcriptomic datasets, Cassan-Wang et al38 identified new potential transcription factor genes that are regulated by MYB transcription factors and are involved in cell wall biosynthesis38, so it is likely that PtrMYB152 may also regulate genes encoding other transcription factors, which in turn activate some of the secondary cell well biosynthesis genes.

Poplar orthologs of Arabidopsis transcription factors in the cell wall transcription regulatory network.
Arabidopsis transcription factors are indicated in blue, poplar transcription factors in pink. Arabidopsis NAC domain transcription factors (e.g., NST1, SND1, VND6, VND7) act as master regulators and directly activate the MYB and KNAT7 targets shown, as well as biosynthetic genes and Arabidopsis MYB genes work in feed-forward mechanism to activate biosynthetic target genes (see Demura and Ye25, Zhao and Dixon35 and Zhong et al.22 for recent reviews). Arabidopsis KNAT7 is transcriptional repressor that directly or indirectly down regulates biosynthetic target genes20. AtMYB43 has been implicated in the Arabidopsis network, but is not known to be a direct target of NAC domain master regulators and its target genes have not been identified. Poplar NAC domain orthologs of Arabidopsis master regulators have been identified by Zhong et al.19 (PtrWND genes) and Ohtani et al.36 (PtVNS genes). The poplar KNAT7 ortholog was described by Li et al.37. Poplar MYB genes are named according to Wilkins et al.26 and are described in this study, McCarthy et al.11 and Winzell et al.12. Solid lines indicate known direct targets in Arabidopsis; dashed lines indicate regulator relationships but unknown direct targets. Arrows, positive regulation; line with block, negative regulation.
In summary, our study provides evidence that PtrMYB152 regulates secondary cell wall biosynthesis in way similar to that of Arabidopsis MYBs. Information gained from this study may be used to guide to genetic modification of poplars to make it an improved feedstock for biofuels.
Methods
Identification of poplar homologues of Arabidopsis R2R3 MYB transcription factors
To identify poplar homologues of Arabidopsis R2R3 MYB transcription factors that are known to be involved in regulating secondary cell wall biosynthesis, the entire amino acid sequences of Arabidopsis AtMYB20 (At1g66230), AtMYB43 (At5g16600), AtMYB46 (At5g12870), AtMYB58 (At1g16490) and AtMYB63 (At1g79180) were used in BLAST searches of the Populus trichocarpa proteome (www.phytozome.net). Full-length amino acid sequences of the selected poplar and Arabidopsis MYBs were subjected to phylogenetic analysis using AliBee-Multiple Alignment software, release 2.0 (http://www.genebee.msu.su/services/malign_reduced.html) and the most closely related homologs, based on this analysis, were chosen for in-depth characterization. We used poplar MYB gene names according to the nomenclature of Wilkins et al.26, corresponding gene names are as follows: PtrMYB018, POPTR_0004s08480; PtrMYB021, POPTR_0009s05860; PtrMYB028, POPTR_0005s09930; PtrMYB152, POPTR_0017s02850; PtrMYB192; POPTR_0007s08190.
Plant materials and growth conditions
Populus trichocarpa tissue was collected from about 15-year-old field-grown poplar trees in July and used for PtrMYB152 gene cloning and gene expression assays.
Arabidopsis thaliana (Arabidopsis) ecotype Columbia (Col-0) was used for plant transformation. For RNA isolation from Arabidopsis seedlings, seeds were sterilized and sown on ½ strength Murashige & Skoog (MS) basal medium with vitamins (PlantMedia, http://www.plantmedia.com). For plant transformation or phenotypic analysis, seeds were directly sown into soil and grown at 22°C, with a 14/10 hour photoperiod at approximately 120 μmol m−2s−1.
RNA isolation, RT-PCR and quantitative RT-PCR (qRT-PCR)
Total RNA from poplar tissue was isolated using PureLink Plant RNA Reagent (Invitrogen), cleaned with RNeasy Plant Mini Kit (Qiagen) and treated with RNase-Free DNase (Qiagen) as described previously39,40. Total RNA from Arabidopsis seedlings was isolated using RNeasy Plant Mini Kit and treated with RNase-Free DNase set according to the manufacturer's instructions. Three different pools of tissues were generated for each sample and analyzed independently as biological replicates.
Two μg of total RNA was used for reverse transcriptase synthesis using the Omniscript RT Kit (Qiagen) according to the manufacturer's instructions. ACTIN2 (ACT2) and poplar elongation factor C672 gene were used as controls for qRT-PCR. Gene primers use for RT-PCR analysis of PtrMYB152 are: 5′-TCCACTAATATCGTATCTGAAC-3′ and 5′-TAGCGGAACTTCATCTATGCA-3′. Primers for RT-PCR analysis of expression of Arabidopsis genes were as described by Li et al.20,27.
Constructs
To generate the PtrCesA8prom:GUS construct, poplar DNA was isolated from the developing xylem collected from field grown poplar tree using DNeasy Plant Mini Kit (Qiagen) according to the manufacturer's instructions. A fragment that covers the region −2905 to +1 of the start codon of PtrCesA8 gene was amplified by PCR using isolated DNA as the template. PCR products were then cloned into pUC19 vector to drive the expression of the GUS reporter gene.
To generate HA or GD tagged constructs for PtrMYB152, the full-length open-reading frame (ORF) of PtrMYB152 was amplified by RT-PCR using RNA isolated from the developing xylem collected from field grown poplar and the PCR products cloned in frame with an N-terminal HA or GD tag into the pUC19 vector under the control of either the double 35S enhancer promoter of CaMV41 or the PtrCesA8 promoter.
For plant transformation, corresponding constructs in pUC19 vector were digested with EcoRI, then subcloned into the binary vector pPZP211. Five-week-old plants with several mature flowers on the main inflorescence were used for transformation with various constructs via Agrobacterium tumefaciens (GV3101) using the floral dip method. Phenotypes of transgenic plants were examined in the T1 generation and confirmed in T2 to T4 generations. For all transgenic plants, at least 4 transgenic lines with similar phenotypes were collected and evaluated. Overexpression of PtrMYB152 in related lines was confirmed by RT-PCR.
Plasmid DNA isolation, protoplast transfection and β-glucuronidase (GUS) activity assay
All reporter and effector plasmids used in transfection assays were prepared using the EndoFree Plasmid Maxi Kit (Qiagen, Valencia, CA). Protoplasts isolation, transfection and GUS activity assays were performed as described previously41.
Histochemical staining for GUS activity
5-bromo-4-chloro-3-indolyl-β-D-glucuronide (X-Gluc; Rose Scientific Ltd) was used as the substrate for the histochemical staining of GUS activity in seven-day old seedlings and stem sections from 6-week old plant.
Microscopy
To study inflorescence stem development, primary stems from soil grown plants at principal growth stage 6.1032 were collected and sections ~2 cm away from the bases of the stems were used for phenotypic analysis.
The sections were stained either in aqueous 0.02% toluidine blue O (Sigma) or in phloroglucinol (saturated solution in 2 M HCI) and viewed immediately using an Olympus AX70 light microscope. Tissue embedding and light and transmission electron microscopy (TEM) have been described previously20. Photos were taken under an Olympus AX70 light microscope or a Hitachi H7600 PC-TEM (Hitachi Ltd., Tokyo, Japan). Cell wall thickness was measured from TEM micrographs using ImageJ software (http://rsb.info.nih.gov/ij/index.html, Maryland, USA). For each genotype, secondary cell wall measurements were taken from at least 50 separate cells at standardized positions. The measurements were subjected to statistical analysis using the Student's t test (http://www.graphpad.com/quickcalcs/ttest1.cfm). The quantitative differences between wild type and transgenic lines in all data sets were shown to be statistically significant.
Chemical analysis
Five-cm stem fragments from the bases of primary inflorescence stems were used as material for chemical analysis. Lignin content was determined by a modified Klason method as described42.
Ten mg of ground, extract-free oven-dried flour was used for lignin monomer composition analysis, using a downscaled thioacidolysis procedure followed by gas chromatographic analysis as described in details by Robinson and Mansfield43.
References
Dubos, C. et al. MYB transcription factors in Arabidopsis. Trends Plant Sci 15, 573–581 (2010).
Stracke, R., Werber, M. & Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr Opin Plant Bio 4, 447–456 (2001).
Oppenheimer, D. G., Herman, P. L., Sivakumaran, S., Esch, J. & Marks, M. D. A myb gene required for leaf trichome differentiation in Arabidopsis is expressed in stipules. Cell 67, 483–493 (1999).
Lee, M. M. & Schiefelbein, J. WEREWOLF, a MYB-related protein in Arabidopsis, is a position-dependent regulator of epidermal cell patterning. Cell 67, 473–483 (1999).
Shin, R. et al. The Arabidopsis transcription factor MYB77 modulates auxin signal transduction. Plant Cell 19, 2440–2453 (2007).
Byrne, M. E. et al. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 408, 967–971 (2000).
Zhong, R., Lee, C., Zhou, J., McCarthy, R. L. & Ye, Z. H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 20, 2763–2782 (2008).
Bhargava, A., Mansfield, S. D., Hall, H. C., Douglas, C. J. & Ellis, B. E. MYB75 functions in regulation of secondary cell wall formation in the Arabidopsis inflorescence stem. Plant Physiol 154, 1428–1438 (2010).
Bhargava, A. et al. The interacting MYB75 and KNAT7 transcription factors modulate secondary cell wall deposition both in stems and seed coat in Arabidopsis. Planta 237, 1199–1211 (2013).
Legay, S. et al. EgMYB1, an R2R3 MYB transcription factor from eucalyptus negatively regulates secondary cell wall formation in Arabidopsis and poplar. New Phytol 188, 774–786 (2010).
McCarthy, R. L. et al. The poplar MYB transcription factors, PtrMYB3 and PtrMYB20, are involved in the regulation of secondary wall biosynthesis. Plant Cell Physiol 51, 1084–1090 (2010).
Winzell, A., Aspeborg, H., Wang, Y. & Ezcurra, I. Conserved CA-rich motifs in gene promoters of Pt x tMYB021-responsive secondary cell wall carbohydrate-active enzymes in Populus. Biochem Biophys Res Commun 394, 848–853 (2010).
Liepman, A. H., Wightman, R., Geshi, N., Turner, S. R. & Scheller, H. V. Arabidopsis -a powerful model system for plant cell wall research. Plant J 61, 1107–1121 (2010).
Kubo, M. et al. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev 19, 1855–1860 (2005).
Zhong, R. & Ye, Z. H. Regulation of cell wall biosynthesis. Curr Opin Plant Biol 10, 564–572 (2007).
Zhong, R., Demura, T. & Ye, Z. H. SND1, a NAC domain transcription factor, is a key regulater of secondary wall synthesis in fibers of Arabidopsis. Plant Cell 18, 3158–3170 (2006).
Mitsuda, N. et al. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 19, 270–280 (2007).
Ohashi-Ito, K., Oda, Y. & Fukuda, H. Arabidopsis VASCULAR-RELATED NAC-DOMAIN6 directly regulates the genes that govern programmed cell death and secondary wall formation during xylem differentiation. Plant Cell 22, 3461–3473 (2010).
Zhong, R., Lee, C. & Ye, Z. H. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol Plant 3, 1087–1103 (2010).
Li, E., Wang, S., Liu, Y., Chen, J.-G. & Douglas, C. J. OVATE FAMILY PROTEIN4 (OFP4) interaction with KNAT7 regulates secondary cell wall formation in Arabidopsis thaliana. Plant J 67, 328–341 (2011).
Wang, S. et al. Arabidopsis Ovate Family Proteins, a novel transcriptional repressor family, control multiple aspects of plant growth and development. PLoS ONE 6, e23896 (2011).
Zhong, R., Lee, C. & Ye, Z. H. Evolutionary conservation of the transcriptional network regulating secondary cell wall biosynthesis. Trends Plant Sci 15, 625–632 (2010).
Groover, A. T. What genes make a tree a tree? Trends in Plant Sci 10, 210–214 (2005).
Jansson, S. & Douglas, C. J. Populus: a model system for plant biology. Annu Rev Plant Biol 58, 435–458 (2007).
Demura, T. & Ye, Z. H. Regulation of plant biomass production. Curr Opin Plant Biol 13, 200–204 (2010).
Wilkins, O., Nahal, H., Foong, J., Provart, N. J. & Campbell, M. M. Expansion and diversification of the Populus R2R3-MYB family of transcription factors. Plant physiol 149, 981–993 (2009).
Karpinska, B. et al. MYB transcription factors are differentially expressed and regulated during secondary vascular tissue development in hybrid aspen. Plant Mol Biol 56, 255–270 (2004).
Wang, M., Qi, X., Zhao, S., Zhang, S. & Lu, M. Z. Dynamic changes in transcripts during regeneration of the secondary vascular system in Populus tomentosa Carr. revealed by cDNA microarrays. BMC Genomics 10, 215 (2009).
Zhong, R., McCarthy, R. L., Lee, C. & Ye, Z. H. Dissection of the transcriptional program regulating secondary wall biosynthesis during wood formation in poplar. Plant Physiol 157, 1452–1468 (2011).
Kumar, M. et al. An update on the nomenclature for the cellulose synthase genes in Populus. Trends Plant Sci 14, 248–254 (2009).
Chen, Z. et al. Disruption of the cellulose synthase gene, AtCesA8/IRX1, enhances drought and osmotic stress tolerance in Arabidopsis. Plant J 43, 273–283 (2005).
Boyes, D. C. et al. Growth stage-based phenotypic analysis of Arabidopsis: a model for high throughput functional genomics in plants. Plant Cell 13, 1499–1510 (2001).
Chapple, C. C., Vogt, T. & Eills, B. E. Somerville CR: An Arabidopsis mutant defective in the general phenylpropanoid pathway. Plant Cell 4, 1413–1424 (1992).
Pauly, M. & Keegstra, K. Cell-wall carbohydrates and their modification as a resource for biofuels. Plant J. 54, 559–68 (2008).
Zhao, Q. & Dixon, R. A. Transcriptional networks for lignin biosynthesis: more complex than we thought? Trends in Plant Sci 16, 227–233 (2011).
Ohtani, M. et al. Ohme-Takagi, T. Demura, A NAC domain protein family contributing to the regulation of wood formation in poplar. Plant J 67, 499–512 (2011).
Li, E. et al. The class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytol 194, 102–115 (2012).
Cassan-Wang, H. et al. Identification of novel transcription factors regulating secondary cell wall formation in Arabidopsis. Front Plant Sci 4, 189 (2013).
Geraldes, A. et al. SNP discovery in black cottonwood (Populus trichocarpa) by population transcriptome resequencing. Mol Ecol Resour Supplemental 1 81–92 (2011).
Liu, S. et al. R2R3 MYB transcription factor PtrMYB192 regulates flowering time in Arabidopsis by activating Flowering Locus C. J Plant Biol. 56, 243–250 (2013).
Wang, S., Tiwari, S. B., Hagen, G. & Guilfoyle, T. J. AUXIN RESPONSE FACTOR7 restores the expression of auxin-responsive genes in mutant Arabidopsis leaf mesophyll protoplasts. Plant Cell 17, 1979–1993 (2005).
Coleman, H. D., Park, J. Y., Nair, R., Chapple, C. & Mansfield, S. D. RNAi mediated suppression of p-coumaroyl-CoA 3′-hydroxylase in hybrid poplar impacts lignin deposition and soluble secondary metabolism. Proc Natl Acad Sci USA 105, 4501–4506 (2008).
Robinson, A. R. & Mansfield, S. D. Rapid analysis of poplar lignin monomer composition by a streamlined thioacidolysis procedure and near-infrared reflectance-based prediction modeling. Plant J 58, 706–714 (2009).
Acknowledgements
We thank Drs. Tom Guilfoyle and Gretchen Hagen (University of Missouri-Columbia) for providing vectors for protoplast transfection assays, the UBC Bioimaging Facility for technical assistance and all members in the Applied Genomics Innovation Program (AGIP) project for their helpful discussion. This work was supported by funds from the Genome British Columbia (www.genomebc.ca) Applied Genomics Innovation Program project 103BIO to C.J.D and S.D.M. and by the Programme for Introducing Talents to Universities (B07017) and a startup fund from Northeast Normal University (www.nenu.edu.cn) to S.W. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
S.W., J.G.C., S.D.M. and C.J.D. designed the research. S.W. performed homolog identification, gene cloning, transgenic plants generation, protoplast transfection and drafted the manuscript. E.L. performed phenotype analysis and qRT-PCR analysis. I.P. performed chemical analysis. J.G.C., S.D.M. and C.J.D. modified the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. The images in this article are included in the article's Creative Commons license, unless indicated otherwise in the image credit; if the image is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the image. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article

Cite this article
Wang, S., Li, E., Porth, I. et al. Regulation of secondary cell wall biosynthesis by poplar R2R3 MYB transcription factor PtrMYB152 in Arabidopsis. Sci Rep 4, 5054 (2014). https://doi.org/10.1038/srep05054
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep05054
This article is cited by
-
Genomic variation and candidate genes dissect quality and yield traits in Boehmeria nivea (L.) Gaudich
Cellulose (2024)
-
Comprehensive analysis of the MYB transcription factor gene family in Morus alba
BMC Plant Biology (2022)
-
Rootstock effects on scion gene expression in maritime pine
Scientific Reports (2021)
-
Expression profiles of genes regulated by BplMYB46 in Betula platyphylla
Journal of Forestry Research (2019)
-
Localization of gene expression, tissue specificity of Populus xylosyltransferase genes by isolation and functional characterization of their promoters
Plant Cell, Tissue and Organ Culture (PCTOC) (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
