
Abstract
The outer membrane structure is common in Gram-negative bacteria, mitochondria and chloroplasts, and contains outer membrane β-barrel proteins (OMPs) that are essential interchange portals of materials1,2,3. All known OMPs share the antiparallel β-strand topology4, implicating a common evolutionary origin and conserved folding mechanism. Models have been proposed for bacterial β-barrel assembly machinery (BAM) to initiate OMP folding5,6; however, mechanisms by which BAM proceeds to complete OMP assembly remain unclear. Here we report intermediate structures of BAM assembling an OMP substrate, EspP, demonstrating sequential conformational dynamics of BAM during the late stages of OMP assembly, which is further supported by molecular dynamics simulations. Mutagenic in vitro and in vivo assembly assays reveal functional residues of BamA and EspP for barrel hybridization, closure and release. Our work provides novel insights into the common mechanism of OMP assembly.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
Electron microscopy density maps and atomic models have been deposited in the Electron Microscopy Data Bank (EMDB) and PDB, respectively, with accession codes EMD-33763 and 7YE4 (BAM–pair1+4-EspP), EMD-16137 and 8BNZ (BAM–pair1-EspP), EMD-33765 and 7YE6 (BAM–pair2-EspP), and EMD-16138 and 8BO2 (BAM–pair3-EspP). Previously reported structures (5D0Q, 5D0O, 6V05, 5EKQ, 7TT5, 7TT7, 7RI4, 2QOM and 3SLJ) were used as reference for structural comparative analysis.
References
Lundquist, K., Billings, E., Bi, M., Wellnitz, J. & Noinaj, N. The assembly of β‐barrel membrane proteins by BAM and SAM. Mol. Microbiol. 115, 425–435 (2021).
Walther, D. M., Rapaport, D. & Tommassen, J. Biogenesis of β-barrel membrane proteins in bacteria and eukaryotes: evolutionary conservation and divergence. Cell. Mol. Life Sci. 66, 2789–2804 (2009).
Tommassen, J. Assembly of outer-membrane proteins in bacteria and mitochondria. Microbiology 156, 2587–2596 (2010).
Gentle, I., Gabriel, K., Beech, P., Waller, R. & Lithgow, T. The Omp85 family of proteins is essential for outer membrane biogenesis in mitochondria and bacteria. J. Cell Biol. 164, 19–24 (2004).
Noinaj, N., Gumbart, J. C. & Buchanan, S. K. The β-barrel assembly machinery in motion. Nat. Rev. Microbiol. 15, 197–204 (2017).
Knowles, T. J., Scott-Tucker, A., Overduin, M. & Henderson, I. R. Membrane protein architects: the role of the BAM complex in outer membrane protein assembly. Nat. Rev. Microbiol. 7, 206–214 (2009).
Sasaki, K. et al. VDAC: old protein with new roles in diabetes. Am. J. Physiol. Physiol. 303, C1055–C1060 (2012).
Bender, A. et al. TOM40 mediates mitochondrial dysfunction induced by α-synuclein accumulation in Parkinson’s disease. PLoS ONE 8, e62277 (2013).
Webb, C. T., Heinz, E. & Lithgow, T. Evolution of the β-barrel assembly machinery. Trends Microbiol. 20, 612–620 (2012).
Malinverni, J. C. et al. YfiO stabilizes the YaeT complex and is essential for outer membrane protein assembly in Escherichia coli. Mol. Microbiol. 61, 151–164 (2006).
Wu, T. et al. Identification of a multicomponent complex required for outer membrane biogenesis in Escherichia coli. Cell 121, 235–245 (2005).
Tellez, R. Jr & Misra, R. Substitutions in the BamA β-barrel domain overcome the conditional lethal phenotype of a ΔbamB ΔbamE strain of Escherichia coli. J. Bacteriol. 194, 317–324 (2012).
Imai, Y. et al. A new antibiotic selectively kills Gram-negative pathogens. Nature 576, 459–464 (2019).
Kaur, H. et al. The antibiotic darobactin mimics a β-strand to inhibit outer membrane insertase. Nature 593, 125–129 (2021).
Luther, A. et al. Chimeric peptidomimetic antibiotics against Gram-negative bacteria. Nature 576, 452–458 (2019).
Hart, E. M. et al. A small-molecule inhibitor of BamA impervious to efflux and the outer membrane permeability barrier. Proc. Natl Acad. Sci. USA 116, 21748–21757 (2019).
Mori, H. & Ito, K. The Sec protein-translocation pathway. Trends Microbiol. 9, 494–500 (2001).
Manting, E. H., van der Does, C., Remigy, H., Engel, A. & Driessen, A. J. M. SecYEG assembles into a tetramer to form the active protein translocation channel. EMBO J. 19, 852–861 (2000).
Ranava, D., Caumont-Sarcos, A., Albenne, C. & Ieva, R. Bacterial machineries for the assembly of membrane-embedded β-barrel proteins. FEMS Microbiol. Lett. 365, fny087 (2018).
Noinaj, N. et al. Structural insight into the biogenesis of β-barrel membrane proteins. Nature 501, 385–390 (2013).
Bakelar, J., Buchanan, S. K. & Noinaj, N. The structure of the β-barrel assembly machinery complex. Science 351, 180–186 (2016).
Gu, Y. et al. Structural basis of outer membrane protein insertion by the BAM complex. Nature 531, 64–69 (2016).
Iadanza, M. G. et al. Lateral opening in the intact β-barrel assembly machinery captured by cryo-EM. Nat. Commun. 7, 12865 (2016).
Han, L. et al. Structure of the BAM complex and its implications for biogenesis of outer-membrane proteins. Nat. Struct. Mol. Biol. 23, 192–196 (2016).
Dong, C., Yang, X., Hou, H.-F., Shen, Y.-Q. & Dong, Y.-H. Structure of Escherichia coli BamB and its interaction with POTRA domains of BamA. Acta Crystallogr. Sect. D Biol. Crystallogr. 68, 1134–1139 (2012).
Gatzeva-Topalova, P. Z., Warner, L. R., Pardi, A. & Sousa, M. C. Structure and flexibility of the complete periplasmic domain of BamA: the protein insertion machine of the outer membrane. Structure 18, 1492–1501 (2010).
Kim, K. H., Aulakh, S. & Paetzel, M. Crystal structure of β-barrel assembly machinery BamCD protein complex. J. Biol. Chem. 286, 39116–39121 (2011).
Tomasek, D. et al. Structure of a nascent membrane protein as it folds on the BAM complex. Nature 583, 473–478 (2020).
Xiao, L. et al. Structures of the β‐barrel assembly machine recognizing outer membrane protein substrates. FASEB J. 35, e21207 (2021).
Doyle, M. T. & Bernstein, H. D. Bacterial outer membrane proteins assemble via asymmetric interactions with the BamA β-barrel. Nat. Commun. 10, 3358 (2019).
Doyle, M. T. et al. Cryo-EM structures reveal multiple stages of bacterial outer membrane protein folding. Cell 185, 1143–1156 (2022).
Wu, R. et al. Plasticity within the barrel domain of BamA mediates a hybrid-barrel mechanism by BAM. Nat. Commun. 12, 7131 (2021).
Brunder, W., Schmidt, H. & Karch, H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157: H7 cleaves human coagulation factor V. Mol. Microbiol. 24, 767–778 (1997).
Khan, S., Mian, H. S., Sandercock, L. E., Chirgadze, N. Y. & Pai, E. F. Crystal structure of the passenger domain of the Escherichia coli autotransporter EspP. J. Mol. Biol. 413, 985–1000 (2011).
Barnard, T. J., Dautin, N., Lukacik, P., Bernstein, H. D. & Buchanan, S. K. Autotransporter structure reveals intra-barrel cleavage followed by conformational changes. Nat. Struct. Mol. Biol. 14, 1214–1220 (2007).
Yuan, X. et al. Molecular basis for the folding of β-helical autotransporter passenger domains. Nat. Commun. 9, 1395 (2018).
Peterson, J. H., Hussain, S. & Bernstein, H. D. Identification of a novel post‐insertion step in the assembly of a bacterial outer membrane protein. Mol. Microbiol. 110, 143–159 (2018).
Doyle, M. T. & Bernstein, H. D. BamA forms a translocation channel for polypeptide export across the bacterial outer membrane. Mol. Cell 81, 2000–2012 (2021).
Lundquist, K., Bakelar, J., Noinaj, N. & Gumbart, J. C. C-terminal kink formation is required for lateral gating in BamA. Proc. Natl Acad. Sci. USA 115, E7942–E7949 (2018).
White, P. et al. The role of membrane destabilisation and protein dynamics in BAM catalysed OMP folding. Nat. Commun. 12, 4174 (2021).
Doerner, P. A. & Sousa, M. C. Extreme dynamics in the BamA β-barrel seam. Biochemistry 56, 3142–3149 (2017).
Liu, J. & Gumbart, J. C. Membrane thinning and lateral gating are consistent features of BamA across multiple species. PLoS Comput. Biol. 16, e1008355 (2020).
Roman-Hernandez, G., Peterson, J. H. & Bernstein, H. D. Reconstitution of bacterial autotransporter assembly using purified components. eLife 3, e04234 (2014).
Tang, X. et al. Structural insights into outer membrane asymmetry maintenance in Gram-negative bacteria by MlaFEDB. Nat. Struct. Mol. Biol. 28, 81–91 (2021).
Mastronarde, D. N. SerialEM: a program for automated tilt series acquisition on Tecnai microscopes using prediction of specimen position. Microsc. Microanal. 9, 1182–1183 (2003).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Zhang, K. Gctf: real-time CTF determination and correction. J. Struct. Biol. 193, 1–12 (2016).
Scheres, S. H. W. Semi-automated selection of cryo-EM particles in RELION-1.3. J. Struct. Biol. 189, 114–122 (2015).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 66, 213–221 (2010).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 66, 486–501 (2010).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Varadi, M. et al. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 50, D439–D444 (2022).
Trabuco, L. G., Villa, E., Mitra, K., Frank, J. & Schulten, K. in Single-Particle Cryo-Electron Microscopy: The Path Toward Atomic Resolution: Selected Papers of Joachim Frank with Commentaries (ed. Frank, J.) 433–443 (World Scientific, 2008).
Huang, J. et al. CHARMM36m: an improved force field for folded and intrinsically disordered proteins. Nat. Methods 14, 71–73 (2016).
Jo, S., Kim, T., Iyer, V. G. & Im, W. CHARMM‐GUI: a web‐based graphical user interface for CHARMM. J. Comput. Chem. 29, 1859–1865 (2008).
Phillips, J. C. et al. Scalable molecular dynamics with NAMD. J. Comput. Chem. 26, 1781–1802 (2005).
Klauda, J. B. et al. Update of the CHARMM all-atom additive force field for lipids: validation on six lipid types. J. Phys. Chem. B 114, 7830–7843 (2010).
Darden, T., York, D. & Pedersen, L. Particle mesh Ewald: an N⋅ log (N) method for Ewald sums in large systems. J. Chem. Phys. 98, 10089–10092 (1993).
Feller, S. E., Zhang, Y., Pastor, R. W. & Brooks, B. R. Constant pressure molecular dynamics simulation: the Langevin piston method. J. Chem. Phys. 103, 4613–4621 (1995).
Miyamoto, S. & Kollman, P. A. Settle—an analytical version of the shake and rattle algorithm for rigid water models. J. Comput. Chem. 13, 952–962 (1992).
Acknowledgements
We thank Y. Q. Wei and B. R. Dong for their continuous support, encouragement and advice, C. Ma at Zhejiang University School of Medicine for his assistance in protein purification and Z. Su at SKLB Duyu High Performance Computing Centre of Sichuan University for help in figure preparation. This work was partially supported by the National Key Research and Development Program of China (2018YFA0507700 to X. Zhang, 2021YFA1301900 and 2021YFA1301203 to H.D., 2017YFA0504803 to X. Zhang, and 2021YFA1201201 and 2021YFF1200404 to R.Z.); the National Natural Science Foundation of China (31900039 and 32170029 to X.T., 32000844 to S.C., 81971974 to H.D. and U1967217 to R.Z.); the National Center of Technology Innovation for Biopharmaceuticals (NCTIB2022HS02010 to R.Z.); the Fundamental Research Funds for Central Universities (226-2022-00043 and 226-2022-00192 to R.Z.); the 1.3.5 Project for Disciplines Excellence of West China Hospital, Sichuan University (ZYYC20021 to H.D.); the National Independent Innovation Demonstration Zone Shanghai Zhangjiang Major Projects (ZJZX2020014 to R.Z.); the Starry Night Science Fund at Shanghai Institute for Advanced Study of Zhejiang University (SN-ZJU-SIAS-003/006/009 to R.Z.); and BirenTech Research (BR-ZJU-SIAS-001 to R.Z.); as well as grants from Laboratory and Equipment Management Department, Zhejiang University (SYB202132 to S.C.).
Author information
Authors and Affiliations
Contributions
H.D. and X.T. conceived and designed the experiments. R.Z. and K.C.C. designed the molecular models and simulations. X.T., H.D., C.S. and Q.L. made the constructs for protein expression. X.T., C.S., Q.L. and Z.Z. expressed and purified the proteins. C.S. and Q.L. did the mutagenesis, the transport assays and the cell-based assays. C.S., Q.L. and Z.Z. prepared the samples. S.C. and X. Zhang undertook data collection, processed electron microscopy data and performed structure construction. R.Z., K.C.C. and T.X. performed the MD simulations. X.W., X. Zhu, G.L. and B.L. analysed the experimental data and prepared the figures. H.D., X.T. and C.D. did the model building and refinement. H.D. and X.T. wrote the manuscript. X. Zhang, R.Z., S.C. and C.D. revised the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Nikolaus Pfanner, Alessandra Polissi, Robert Tampe and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
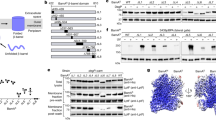
Extended Data Fig. 1 Analysis of the purified BAM-EspP proteins containing cysteine pair1+4, pairs1, pair2 and pair3.
a, Scheme of outer membrane protein (OMP) transport and assemble from the cytoplasm to the outer membrane via BAM. Newly synthesized OMP is translocated across the IM through SecYEG to the periplasm, where chaperons SurA and Skp carry the nascent OMP to BAM for assembly. b, Schematic diagram of recombinant BamABCDE and maltose-binding protein (MBP) fused EspP protein containing β-barrel and short passenger peptide (upper panel). Schematic BAM-EspP complex showing positions of cysteine pair mutations (lower panel). Cysteine pair 1: BamA(G431C)-EspP(N1293C), cysteine pair 2: BamA(N427C)-EspP(R1297C), cysteine pair 3: BamA (S425C)-EspP(S1299C) and cysteine pair 4: BamA(G781C)-EspP(A1043C). c-j, SDS-PAGE (c-f) and SEC profile (g-j) of purified BAM-EspP proteins by size exclusion chromatography using a Superdex200 Increase 10/300 column (void volume≈4.8ml) in the absence or presence of DTT. The results shown are representative data of experiments n = 3. For gel source data, see Supplementary Fig. 1.
Extended Data Fig. 2 Assessment of disulfide bond formations of cysteine pair1, pair2 and pair3.
In vivo disulfide bond formations of the BamA cysteine mutants N431C (a), N427C (b) and S425C (c) and the EspP cysteine mutants R1297C, N1295C, R1297C and S1299C. The crosslinked BamA-MBPEspP, the immature MBPEspP and mature EspP were detected by anti-Flag for EspP (upper two blots) and anti-Myc for BamA (lower two blots) in the absence and presence of DTT. Non-specific band is indicated with *. The results shown are representative data of experiments n = 3. For source data, see Supplementary Fig. 1.
Extended Data Fig. 3 Physiological assessment of the crosslinked BAM-EspP complexes.
a–d, In vivo folding state of the crosslinked BAM-EspP by cysteine pair1 (a), pair1+4 (b), pair2 (c) and pair3 (d) were monitored over 90 mins. MBP-EspP intermediate, mature EspP and BamA were detected by western blots under reducing conditions. EspP mutant of the C-terminal loop deletion (del S1281-D1289 of ECL6) was used as a positive control. Anti-RpoB immunoblotting was used as loading control. The results shown are representative data of experiments n = 3. e–h, The 200-ns molecular dynamics (MD) stimulations of the crosslinked (green line) and unlinked wildtype (purple line) BAM-EspP complex in lipidic nanodisc (e,f) or unlinked wildtype BAM-EspP complex in nanodisc/PL bilayer (red line) and LPS and PL membrane (blue line) (g,h). For blot source data, see Supplementary Fig. 1.
Extended Data Fig. 4 Flow charts for cryo-EM structure determination of BAM-pair1+4-EspP and BAM-pair1-EspP complexes in nanodisc.
a,f, Representitive cryo-EM microscope images of the BAM-pair1+4-EspP (a) and BAM-pair1-EspP (f) complexes. b,g, Scheme of data collection, two- and three- dimensional classifications, refinements and the local cryo-EM resolutions of the density maps for BAM-pair1+4-EspP (b) and BAM-pair1-EspP (g). c, Selected two-dimensional classification images of BAM-pair1+4-EspP. d,h, Gold-standard Fourier Shell Correction (FSC) curves for structure resolution determination for BAM-pair1+4-EspP (d) and BAM-pair1-EspP (h). e,i, Cryo-EM maps of the selected BamA and EspP β-strands of BAM-pair1+4-EspP (e) and BAM-pair1-EspP (i).
Extended Data Fig. 5 Flow charts for cryo-EM structure determination of BAM-pair2-EspP and BAM-pair3-EspP complexes in nanodisc.
a,f, Representative cryo-EM microscope image of BAM-pair2-EspP (a) and BAM-pair3-EspP (f) complex. b,g, Cryo-EM image collection, reconstitution, two-dimensional and three-dimensional classifications, refinements and local resolutions of the density map for BAM-pair2-EspP (b) and BAM-pair3-EspP (g). c, Selected two-dimensional image of BAM-pair2-EspP complex. d,h, FSC curves for the structure resolution determination for BAM-pair2-EspP (d) and BAM-pair3-EspP (h). e,i, Cryo-EM maps of the selected BamA and EspP β-strands of BAM-pair2-EspP (e) and BAM-pair3-EspP (i).
Extended Data Fig. 6 Superimposition of the BamA-EspP structures with other reported BAM structures.
a, Side views of hybrid barrels of BAM-pair1+4-EspP, showing the interface between the N-terminus of BamA and C-terminus of EspP (left) and the C-terminus of BamA and N-terminus of EspP with exposed hydrophobic side chains (right). The red circle indicates the position of the disulfide bond formed between BamA C1043 and EspP C781. b, Superimposition of the BAM-pair1+4-EspP and BAM-pair1-EspP structures from side view (left) and top view (right) c, Superimposition of the BamA barrel of the BAM-pair1-EspP structure (light green) with that of the closed crystal structure (PDB: 5D0O, salmon) (left) and open crystal structure (PDB: 5D0Q, pale yellow) (right). d, Superimposition of BAM-pair1+4-EspP complex (dark cyan) with BAM-BamA∆L1 complex (PDB: 6V05, dark purple) in the side (left) and top (mid) views of the hybrid barrels and the periplasmic ring (right). e, Superimposition of the BamA and EspP hybrid barrels of pair1 (light green) and pair1+4 (dark cyan) complexes from the top view. The side chains of R1297 of the pair1+4 (red) and pair1 (blue) structures, showing different orientations. f, In vivo folding assessment of BAM-EspP(R1297A) over 90min. MBP-EspP intermediate, mature EspP and BamA were detected by western blots under reducing conditions. The result shown is representative data of experiments n=3. For blot source data, see Supplementary Fig. 1.
Extended Data Fig. 7 In vitro assembly assays set-up.
a, Scheme of in vitro folding assay. Urea denatured EspP is mixed with BAM proteoliposomes in presence of SurA. b, The set-up of the in vitro EspP assembly assay by western blot. The folded EspP and BamA in liposomes are tested by heat denaturation. c, The orientation of BAM complex in liposomes was determined by proteinase K treatment. d, Unfolded EspP was degraded by proteinase K. e, Immunoprecipitation against SurA (Strep) from the in vitro assembly assay to pull out bounded EspP(Flag) variants to show normal presentation of SurA. f, Protein detection of EspP(∆L1) and EspP(∆L6) variants with or without alanine linker restoration following 30min in vivo assembly by western blot against EspP and BamA. Anti-RpoB was used as loading control. g, Cellular viability assays of BamA mutants in the presence of arabinose. The viability assays in the absence of arabinose as shown in Fig. 3h. h, SDS-PAGE of purified BAM complex of BamA mutants of g. i, Liposome reconstituted BamA mutants before and after heat denaturation. The results shown are representative data of experiments n=3. For source data, see Supplementary Fig. 1.
Extended Data Fig. 8 Conformational changes of the structures at different states.
a, Superimposition of the hybrid barrel of pair1+4 and pair2 structures (left), showing the closing lateral gate of the pair2 structure with the interactions on BamA ECL1 (right). b, Superimposition of the BamA barrels of the pair2 structure and the closed crystal structure (PDB: 5D0O, salmon). The zoomed section shows the interactions between BamA β1 and β16 of the closed crystal structure (PDB: 5D0O) (right panel) c, Superimposed BamA barrels of the four BAM-EspP structures, showing the conformational changes of the C-terminal end of β16. d, Superimpositions of the periplasmic ring of the pair1+4, pair2 and pair3 structures. e, Superimpositions of the BamA barrels and the periplasmic rings of the pair1 and pair2 structures, with the reference of the pair1+4 structure. The boxed section is showing curved BamA ECL1 in pair1 and pair2 structures with interactions from S436. f, g, Superimposed BamA barrel of pair1+4, pair2 and the lateral closed crystal (5D0O) structures, showing conformational changes in the BamA β1 periplasmic linker (f), POTRA domains (g, left) and POTRA3 loop (g, right). h–l, Superimposed four BAM-EspP structures with highlighted interactions between POTRA5, PT1 and PT4 (green) and between PT1 and BamD (orange). BAM-EspP pair1+4 in dark cyan, pair1 in light green, pair2 in medium purple and pair3 in royal blue.
Extended Data Fig. 9 EspP barrel closes in a zipper-like fashion.
a–e, EspP barrels from the pair1+4 (a), pair1 (b), pair2 (c), pair3 (d) and the crystal mature EspP (PDB: 2QOM, yellow orange) (e) structures (upper panels). The hydrogen bonds formed at the β-seam of the EspP barrel of the corresponding structures are shown in the lower panels. f–l, MD simulation of the EspP from our pair2 structure showing sequential formation of hydrogen bonds within the time monitored (f–k). The monitored hydrogen bonds are indicated in different colors. Structural comparison of our pair2 and the crystal folded EspP structures during the simulated barrel closure (j,l). m, The in vivo disulfide bond formation upon cysteine mutation of the observed interacted residues in the EspP of the pair2 structure. The result shown is representative data of experiments n = 3. For blot source data, see Supplementary Fig. 1.
Extended Data Fig. 10 Structural comparisons of BAM assembly intermediate with the reported crystal BAM structures.
Superimposition of pair1+4 (dark cyan), pair1 (light green), pair2 (medium purple), pair3 (purple) and crystal structures PDB: 5D0Q (pale yellow), 5EKQ (marine) and 5D0O (salmon). a–f, Conformational changes in the BamA barrel of the superimposed structures, showing state transition from the lateral opening to close. g–l, The overall conformational changes of BamA of the superimposed structures from the open state to close state. m–r, Top view of the POTRA1-5 of the superimposed structures from the open to the close states, showing the clockwise rotation of the POTRA domains.
Extended Data Fig. 11 Structural comparisons of the reported ligand bound BamA with our BAM-EspP structures.
a–c, Superimposition of the reported BamA-EspP PDB: 7TT5 (dirty violet) or 7TT7 (olive) with our pair1+4 (dark cyan) (a), pair1 (light green) (b) and pair2 (medium purple) (c) structures, showing the distinct intermediate conformations of BamA and EspP barrel. The lower panels are 90° rotation of the upper panels along x-axis, showing the top view of the hybrid barrels. d–g, Superimposition of BamA from the reported BAM-EspPβ9-12 (PDB: 7RI4) (light pink), closed crystal PDB: 5D0O (light orange) (d) and pair1+4 (dark cyan) structures (e-g), showing the distinct conformations of BamA POTRA domains (d,e) and barrel lateral opening (f) of the BAM-EspPβ9-12 structure. The EspPβ9-12 (medium orchid) is sandwiched between β1 and β16 of BamA (g).
Supplementary information
Supplementary Fig. 1
Uncropped gels and immunoblots. Cropped regions shown in the main and Extended Data Figures are indicated with red dashed lines.
Supplementary Video 1
Dynamics of BAM-EspP for the view of BAM. State transitions of the BAM-EspP complex, showing conformational changes of BAM and EspP for the view of the lateral gate of the BamA β-barrel.
Supplementary Video 2
Dynamics of BAM-EspP for the view of EspP. State transitions of the BAM-EspP complex, showing conformational changes of BAM and EspP for the view of the EspP β-barrel.
Supplementary Video 3
Dynamics of BAM-EspP from the front and top views. State transitions of the BAM-EspP complex, showing conformational changes of BAM and EspP from the front and top views.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Shen, C., Chang, S., Luo, Q. et al. Structural basis of BAM-mediated outer membrane β-barrel protein assembly. Nature 617, 185–193 (2023). https://doi.org/10.1038/s41586-023-05988-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-023-05988-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
