Abstract
Allergic asthma is a T cell-dependent inflammatory lung disease that results from complex interactions between genetic predisposition and environmental factors, including exposure to lipopolysaccharide (LPS). In this study, we have shown that airway LPS exposure was sufficient to induce airway hyperreactivity (AHR) and eosinophil recruitment in mice that had previously experienced an acute episode of allergic asthma. LPS-induced disease reactivation depended on the activation of allergen-specific CD4+ T cells by a subset of lung langerin+ dendritic cells (DCs) that retained the allergen. Upon LPS exposure, migration of langerin+ DCs from lungs to draining lymph nodes increased and LPS-exposed langerin+ DCs instructed CD4+ T cells toward a T helper (Th) 2 response. Selective depletion of langerin+ DCs prevented LPS-induced eosinophil recruitment and T-cell activation, further demonstrating a critical role for langerin+ DCs in disease reactivation. This finding provides a possible explanation for the subclinical worsening of asthmatics following exposure to low-dose LPS.
Similar content being viewed by others


Introduction
Allergic asthma is an inflammatory lung disease characterized by airway hyperreactivity (AHR), excessive mucus production, lung eosinophilia, and chronic inflammation associated with long-term airway remodeling.1 As other atopic disorders, asthma results from complex interactions between genetic predisposition and multiple environmental influences, including exposure to lipopolysaccharide (LPS) and other microbial products.2 Although exposure to LPS early in life protects against asthma,3, 4 exposure to environmental LPS in adults increases the risk factor to develop the disease,5 and asthma severity is related to the LPS levels in the house environment independently of dust mite concentration.6, 7 Furthermore, asthmatic patients exhibited a significant decrease in lung function and enhanced AHR upon inhalation of low levels of LPS in the absence of allergen,8, 9 suggesting that exposure to LPS by itself could cause subclinical airway disease in these individuals. Other studies in mice have shown that the amount of LPS at the time of airway sensitization determined whether allergen-specific T helper (Th)1 or Th2 T-cell responses were induced.10 Furthermore, antigen inhalation in the presence of LPS prevents airway tolerance by inhibiting the development of regulatory T cells and by promoting effector T-cell generation.11 At the time of allergen challenge, LPS administration results in exacerbation of airway inflammation, both by increasing activation of mast cells and Th2 cells and by promoting the recruitment of Th2 lymphocytes to the lungs.12, 13 In addition, signaling through the LPS receptor, Toll-like receptor-4, drives airway Th2 responses in mice exposed to house dust mite.14, 15 Therefore, LPS increases the susceptibility or the severity to asthma or protects from this disease depending on the time and the level of exposure. Despite these studies, the mechanisms by which LPS by itself, independently of the allergen, influences the disease in asthmatic individuals have never been investigated. To address this issue, we have developed a novel experimental model of airway disease in which BALB/c mice were sensitized and challenged with a recombinant antigen and exposed several weeks later to a single low dose of LPS.
Results
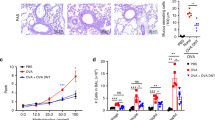
We first determined at what time the airway disease had completely vanished in BALB/c mice that had been sensitized and challenged with the LACK (Leishmania analog of the receptors of activated C kinase) antigen.16 When compared with phosphate-buffered saline (PBS)-challenged mice, LACK-challenged animals exhibited cardinal features of allergic asthma, including AHR, prominent lung eosinophilia, and high titers of LACK-specific serum IgE. At 5 days after antigen challenge, LACK-challenged mice exhibited AHR as demonstrated by increased airway resistance and decreased compliance (Figure 1a). This was not the case at later time points (Figure 1b). Challenge with LACK also induced an increase in the number of eosinophils, lymphocytes, and total cells in bronchoalveolar lavage fluid (BALF). The number of eosinophils reached a plateau between days 3 and 10, declined thereafter, and these cells were undetectable by day 21 (Figure 1d). T cells were readily detectable in BALF at day 3 and were still present at day 21, although in reduced numbers (Figure 1e). Neutrophils were detectable in BALF maximally at day 1 and declined to baseline levels by day 21 (Figure 1f). The increased cellularity observed at day 21 postchallenge (when compared with day 0) mostly comprised a mixture of lymphocytes (Figure 1e) and macrophages (data not shown). Serum LACK-specific IgE concentrations increased significantly at day 5 and remained high up to day 21 after challenge (Figure 1g). As LACK-sensitized and -challenged mice have cleared eosinophils from the airways and do not exhibit any increase in AHR at 3 weeks after antigen challenge, as also evidenced by the fact that resistance in PBS-challenged mice and postasthmatic PBS-exposed mice are not statistically significant (P=0.3, Figures 1a and 2b), these mice will be referred to as “postasthmatic” henceforth.

Airway hyperreactivity (AHR), airway inflammation, and antibody response in LACK (Leishmania analog of the receptors of activated C kinase)-sensitized and challenged mice. LACK-sensitized mice were challenged with (a, b) LACK or phosphate-buffered saline (PBS) or (c–f) LACK and analyzed (a) 5 days or (b–f) at the indicated days after challenge. (a) Airway resistance (R, left panel) and lung dynamic compliance (C, right panel) were measured in response to inhaled methacholine using a FlexiVent in anesthetized and tracheotomized mice. Data are shown for mice that have been challenged with LACK (filled squares) or PBS. (b) AHR was measured by whole-body plethysmography in response to inhaled methacholine (12 mg ml–1) at the indicated days following challenge with LACK (filled bars) or PBS (empty bars). (c–f) Bronchoalveolar lavage fluid (BALF) cells were counted and analyzed by fluorescence-activated cell sorting (FACS). (c) Total number of cells. (d) Number of eosinophils. (e) Number of lymphocytes. (f) Number of neutrophils. (g) Sera of LACK-sensitized and challenged mice were analyzed for LACK-specific IgE contents at the indicated days following LACK challenge. Data are expressed as mean±s.e.m.; n=10 mice per group. Two independent experiments were performed. *P<0.05; **P<0.01; NS, not significant.

Lipopolysaccharide (LPS) induces airway hyperreactivity (AHR) and T cell-mediated lung eosinophilia in postasthmatic mice. (a) Schematic representation of the protocol. Postasthmatic mice were exposed to LPS or phosphate-buffered saline (PBS) 21 days after LACK (Leishmania analog of the receptors of activated C kinase) challenge and analyzed for AHR and bronchoalveolar lavage fluid (BALF) content 3 days later. (b) Airway resistance to inhaled methacholine was measured in anesthetized and tracheotomized postasthmatic mice 3 days after exposure to LPS (filled squares) or PBS (empty squares). Data are expressed as mean±s.e.m.; n=20 mice per group pooled from three experiments. (c) Number (left panel) and phenotype (right panel) of BALF cells in postasthmatic mice upon exposure to LPS (filled bars) or PBS (empty bars). Data are expressed as mean±s.e.m.; n=15 mice per group pooled from three independent experiments. (d, e) Mice were sensitized with LACK, challenged with PBS or LACK at day 0, treated twice or not with antibody to CD4 (GK1.5; 250 μg per mouse) at days 16 and 18, exposed to LPS or PBS at day 21, and analyzed 3 days later. (d) Data show the frequency of eosinophils in BALF in individual mice. Results are expressed as mean±s.e.m.; n=14–35 mice per group pooled from four experiments. (e) Airway resistance to inhaled methacholine was measured in postasthmatic mice 3 days after exposure to PBS (empty symbols) or LPS (filled symbols). Data are expressed as mean±s.e.m.; n=10 mice per group. (f) Thy1.1+/+ RAG-1−/− WT15 T-cell receptor (TCR) transgenic CD4+ T cells were injected into naive BALB/c mice that were then sensitized and challenged with LACK. Mice were exposed to PBS (empty bars) or LPS (filled bars) 21 days after LACK challenge. Mediastinal lymph node (MLN) cells were analyzed by fluorescence-activated cell sorting (FACS) 3 days later upon staining with antibodies to Th1.1, CD4, and CD69. The left panel shows the number of WT15 TCR transgenic T cells in mice exposed to PBS (empty bars) or LPS (filled bars). Results are expressed as mean±s.e.m.; n=6 mice per group pooled from two experiments. The right panels show representative FACS profiles after gating on live cells (upper panel) or Thy1.1+ CD4+ T cells (lower panel). In the lower panel, data show CD69 expression for Thy1.1+ CD4+ T cells from LPS- (open histogram) and PBS (gray histogram)-exposed mice. *P<0.05; **P<0.01; ***P<0.001; NS, not significant.
Postasthmatic mice were exposed to a single dose of LPS (500 ng) or PBS and analyzed for both AHR and the presence of eosinophils in BALF 3 days afterwards (Figure 2a). When compared with PBS-exposed postasthmatic mice, LPS-exposed animals exhibited a 20% increase in airway resistance in response to methacholine (Figure 2b). Furthermore, LPS exposure resulted in a threefold increase in the total number of cells in BALF, in the number of neutrophils, and more surprisingly promoted eosinophil recruitment (Figure 2c). In contrast, LPS exposure did not induce eosinophil recruitment in control mice that had been previously sensitized to LACK but challenged with PBS or in postasthmatic mice exposed to LPS 28 days after antigen challenge (data not shown). Most importantly, the increase in the number of both eosinophils and the AHR to methacholine were abrogated in postasthmatic mice that had been treated with an anti-CD4 depleting antibody before LPS exposure (Figure 2d and e ). To further elucidate the role of CD4+ T cells in LPS-induced eosinophil recruitment, naive BALB/c mice were injected with CD4+ T cells from LACK-specific WT15 T-cell receptor (TCR) transgenic mice, sensitized, and challenged with LACK, and further exposed 21 days later to either LPS or PBS. LPS exposure resulted in a three- to four-fold increase in the number of WT15 cells in mediastinal lymph nodes (MLNs) at 3 days after LPS challenge (Figure 2f). Furthermore, the frequency of WT15 cells that upregulated the early activation marker CD69 increased from 20.2±1.0% in PBS-exposed animals to 33.2±5.4% in LPS-exposed mice, whereas the frequency of CD69+ CD4+ T cells among the endogenous T-cell population was 9.3±3.6% in both groups (Figure 2f, and data not shown). Therefore, LPS exposure promoted an allergen-specific T-cell response and thus eosinophil recruitment to the lung in postasthmatic mice.
The reactivation of LACK-specific T cells induced by LPS exposure in postasthmatic mice suggested that antigen-presenting cells loaded with LACK were still present in these animals at 21 days after antigen exposure. To confirm that this was indeed the case, postasthmatic mice were injected with carboxyfluorescein diacetate succinimidyl ester (CFSE)-labeled WT15 CD4+ T cells and further exposed to either LPS or PBS. Although WT15 CD4+ T cells proliferated in the MLNs of both LPS- and PBS-exposed mice, they proliferated more vigorously in LPS-treated animals, as demonstrated by the increased frequency of cells that had completed ⩾5 divisions after 3 days (64.8±4.6 vs. 31.0±9.8%; Figure 3a, left panels). To determine whether proliferation was dependent on the engagement of the TCR with its cognate antigen, postasthmatic mice were co-injected with both WT15 and ovalbumin (OVA)-specific DO11.10 CFSE-labeled TCR transgenic CD4+ T cells, and exposed to either LPS or PBS alone, or to both LPS and OVA. Whereas WT15 T cells proliferated in all groups, DO11.10 T cells only proliferated in mice exposed to both OVA and LPS (Figure 3b). Therefore, LPS-induced T-cell proliferation in postasthmatic mice was dependent on TCR engagement.

Lipopolysaccharide (LPS) enhances the migration of lung dendritic cells (DCs) to the mediastinal lymph nodes (MLNs) and promotes T helper 2 (Th2) differentiation. (a) CD4+ T cells from Thy1.1+/+ RAG-1−/− WT15 T-cell receptor (TCR) transgenic mice were labeled with carboxyfluorescein diacetate succinimidyl ester (CFSE) and injected (2 × 106 per mouse) into postasthmatic mice. Mice were exposed 12 h later to LPS or phosphate-buffered saline (PBS), and MLN cells were analyzed by fluorescence-activated cell sorting (FACS) 3 days later following staining with antibodies to Thy1.1 and CD4. Data show representative FACS profiles after gating on Thy1.1+ WT15 cells. The numbers indicate the frequency (%) of Thy1.1+ CD4+ T cells that have undergone ⩾5 divisions (left panels). Data are expressed as mean±s.d. and are representative of two independent experiments. Data show the fold increase in the number of WT15 cells recovered in the MLNs of LPS-exposed mice when compared with those recovered in PBS-exposed animals (right panel). Results are expressed as mean±s.d.; n=10 mice per group pooled from two experiments. (b) CD4+ T cells from WT15 and DO.11.10 TCR transgenic mice were labeled with CFSE and co-injected (2 × 106 cells of each per mouse) into postasthmatic mice. Mice were exposed 12 h later to either LPS or PBS alone or to both LPS and ovalbumin (OVA). MLN cells were analyzed by FACS 3 days later following staining with antibodies to Thy1.1, KJ.1-26 and CD4. Data show representative FACS profiles after gating on KJ.1-26+ and Thy1.1+ CD4+ cells of two independent experiments with n=5 mice per group. The gates corresponding to WT15 and DO.11.10 T cells are indicated. (c–e) Postasthmatic mice were exposed to LPS or PBS together with fluorescent latex beads 21 days after LACK (Leishmania analog of the receptors of activated C kinase) challenge. The MLN cells were analyzed by FACS 3 days later. (c) Data show representative FACS profiles after staining with antibody to CD11c. The numbers indicate cell frequency within the indicated gate and are expressed as mean±s.e.m.; n=5 mice per group. Two independent experiments were performed. (d, e) Bead+ DCs were sorted and 104 cells were incubated with 105 CFSE-labeled Thy1.1+/+ RAG-1−/− WT15 CD4+ T cells for 7 days. (d) Data show representative FACS profiles after gating on Thy1.1+ CD4+ T cells. The numbers indicate the frequency of cells that have undergone ⩾6 divisions. (e) Cellular supernatants were assessed for interleukin (IL)-13, IL-5 and interferon-γ (IFN-γ) content by multiplex analysis using Cytometric Bead Array (CBA). Data are expressed as mean±s.d. of triplicate wells from one representative experiment out of two. *P<0.05. (f, g) Postasthmatic langerin-GFP mice were exposed to LPS or PBS together with fluorescent latex beads and MLN cells were analyzed 3 days later. (f) Data show representative FACS profiles after staining with antibodies to CD11c and CD11b. The numbers indicate the mean frequency±s.d. of cells in the indicated gates. (g) Data show the mean number±s.e.m. of langerin− (dark bars) and langerin+ (empty bars) bead+ DCs in PBS- and LPS-exposed mice (left panel). Data show the MFI±s.e.m. upon staining with antibodies to CD80 and CD86 after gating on langerin− (dark bars) or langerin+ (empty bars) bead+ DCs (right panels). n=10 mice per group pooled from two experiments. *P<0.05; **P<0.01; NS, not significant.
Previous studies have shown that lung-derived dendritic cells (DCs) continuously migrate from the lung to the MLN in the steady state, and that DC migration is greatly enhanced upon LPS intranasal (i.n.) treatment.17 Upon OVA administration in the absence of inflammatory stimulus, migratory lung DCs capture the antigen and migrate to the MLNs through a CCR7 (chemokine (C-C motif) receptor 7)-dependent mechanism where they promote the proliferation of both CD4+ and CD8+ T cells and induce T-cell tolerance.18, 19, 20 To confirm that LPS promoted DC migration in postasthmatic mice and to further investigate whether these migratory DCs presented LACK to T cells, fluorescent latex beads were administered together with LPS or PBS to postasthmatic mice and the frequency of bead+ DCs in MLNs was measured. Bead+ DCs were readily detected in the MLNs and their frequency increased from 2.7±0.1% in PBS-exposed mice to 6.1±0.1% in LPS-exposed animals (Figure 3c). Bead+ DCs were next purified from the MLNs of LPS- or PBS-exposed mice, and equal numbers of these cells were incubated with CFSE-labeled WT15 TCR transgenic T cells. Although 22% of T cells had undergone through ⩾6 divisions after 7 days when incubated with DCs from PBS-exposed mice, this frequency increased to 51% when T cells were incubated with bead+ DCs from LPS-exposed mice (Figure 3d). Furthermore, bead+ DCs from LPS-exposed mice induced LACK-specific T cells to secrete twofold more interleukin (IL)-5 and IL-13 and two- to three-fold less interferon-γ (IFN-γ) than those activated by bead+ DCs from PBS-exposed mice (Figure 3e). To further investigate the phenotype of migrating bead+ DCs, we performed additional experiments using langerin-GFP mice that express the enhanced green fluorescent protein (GFP) under the control of the langerin (CD207) gene.21 Fluorescent latex beads were administered together with LPS or PBS to postasthmatic langerin-GFP mice and MLN cells were analyzed by fluorescence-activated cell sorting (FACS) 3 days later. As observed in non-transgenic BALB/c mice, the number of bead+ DCs in langerin-GFP mice was much higher in animals treated with LPS when compared with those treated with PBS. Among bead+ DCs, 54±5% were langerin+ in LPS-exposed mice when compared with 33±3% in PBS-exposed animals (Figure 3f). In both PBS- and LPS-exposed mice, langerin+ bead+ DCs expressed lower levels of CD11b than langerin− bead+ DCs (Figure 3f). When compared with langerin+ bead+ DCs purified from PBS-exposed mice, those harvested from LPS-exposed mice expressed higher levels of both CD80 and CD86 (Figure 3g). Langerin− bead+ DCs from LPS-exposed mice also expressed higher levels of CD80 than those from PBS-exposed animals.
As LPS enhanced DC migration from the lungs to the MLNs, we sought to characterize lung DCs in postasthmatic mice. Four CD11c+ cell populations were identified on the basis of autofluorescence, CD11b, and CD11c expression (Figure 4c). Autofluorescent CD11b+ CD11c+ cells (R3 gate) expressed major histocompatibility complex (MHC) class II molecules, F4/80, and low levels of CD80 and CD86, and were heterogeneous for DEC-205 expression. Autofluorescent CD11b− CD11c+ cells (R4 gate) expressed F4/80, very low or undetectable levels of MHC class II and co-stimulatory molecules, but high levels of DEC-205. Non-autofluorescent CD11b+ CD11cmed cells (R5 gate) expressed F4/80, MHC class II, CD40, CD80, CD86, and CD103, but not langerin, that is, CD207. A small proportion of these cells also expressed DEC-205 and Ly6C. The small proportion of CD11b+ CD11cmed cells expressing Ly6C suggested that this gate contained not only resident lung CD11b+ DCs but also monocyte-derived DCs that rapidly upregulated CD11c and retained expression of Ly6C as a remnant of their monocytic origin. Non-autofluorescent CD11b− CD11c+ cells (R6 gate) expressed MHC class II molecules, CD40, CD80 and CD86, CD103, DEC-205, and CD207. A fraction of these cells expressed F4/80. These four cell populations expressed neither B220 (data not shown) nor CD8. Based on autofluorescence, cell surface phenotype, and published studies,22, 23, 24 we identified cells in R3 as CD11b+ lung macrophages (LMs), cells in R4 as conventional LMs, cells in R5 as cells containing mainly resident lung CD11b+ DCs, and cells in R6 as a population containing mainly langerin+ DCs.

Surface phenotype of lung dendritic cells (DCs) in postasthmatic mice. Lung cells were prepared from postasthmatic mice 21 days following LACK (Leishmania analog of the receptors of activated C kinase) challenge, enriched from CD11c+ cells using magnetic beads, and analyzed by fluorescence-activated cell sorting (FACS). Data show representative FACS profiles upon staining with antibodies to CD11c and CD11b (upper panels). The gates corresponding to autofluorescent (R1) and non-autofluorescent (R2) cells are indicated, as well as those corresponding to autofluorescent CD11b+ (R3), autofluorescent CD11b− (R4), non-autofluorescent CD11b+ (R5), and non-autofluorescent CD11b− (R6) cells. Data also show representative FACS profiles upon staining with the indicated antibody (empty histograms) or isotype control (gray histograms) after gating on the indicated populations (lower panels).
Conventional LMs, CD11b+ LMs, CD11b+ Ly6C− resident lung DCs, and langerin+ DCs were purified from PBS- and LPS-exposed postasthmatic mice and tested for their ability to stimulate WT15 TCR transgenic T cells. In contrast to LMs and CD11b+ LMs, both langerin+ DCs and CD11b+ DCs induced LACK-specific T cells to proliferate (Figure 5a). In both LPS- and PBS-exposed mice, langerin+ DCs were more efficient than CD11b+ DC at stimulating T-cell proliferation. LPS had only a minor effect on the intrinsic capacity of each lung DC subset to induce T-cell proliferation. Furthermore, although CD4+ T cells produced significant amounts of IFN-γ, little IL-13, and no detectable IL-5 or IL-10 when incubated with langerin+ DCs from PBS-exposed mice, they produced large amounts of IL-5, IL-10, and IL-13 and very low IFN-γ when incubated with lung langerin+ DCs from LPS-exposed mice (Figure 5b and data not shown). In contrast, CD4+ T cells produced very low amounts of IFN-γ, no IL-5, no IL-10 (data not shown), and very low amounts of IL-13 when incubated with lung CD11b+ DCs from either LPS- or PBS- exposed mice. IL-4 levels were below the detection level in all samples.

Role of langerin+ and CD11b+ dendritic cells (DCs) in postasthmatic mice in the steady state and upon lipopolysaccharide (LPS) exposure. (a) Postasthmatic langerin-GFP mice were exposed to LPS or phosphate-buffered saline (PBS) and lung cells were prepared 3 days later. Lung macrophages (LMs), CD11b+ LMs, CD11b+ DCs, and langerin+ (GFP+) DCs were sorted and 105 cells were incubated for 7 days with 105 carboxyfluorescein diacetate succinimidyl ester (CFSE)-labeled Thy1.1+/+ WT15 CD4+ T cells. (a) Data show representative fluorescence-activated cell sorting (FACS) profiles after gating on Thy1.1+ CD4+ T cells. (b) Cellular supernatants were analyzed for interferon-γ (IFN-γ), interleukin (IL)-5, and IL-13 content by Cytometric Bead Array (CBA). Data show mean values of triplicates±s.d. of one representative experiment (out of two). (c, d) Langerin-DTR mice were sensitized and challenged with LACK (Leishmania analog of the receptors of activated C kinase), treated with PBS or diphtheria toxin (DT) intranasally (i.n.) 16 days later. Mice were injected with CFSE-labeled WT15 Thy1.1+/+ CD4+ T cells 3 days later, and further exposed to PBS or LPS. (c) Mediastinal lymph node (MLN) and (d) blood cells were analyzed by FACS 72 h following LPS or PBS exposure. (c) Data show representative FACS profiles after gating on Thy1.1+ CD4+ T cells. The numbers indicate the frequency of cells that have undergone ⩾5 divisions. Data are expressed as mean±s.e.m.; n=7 mice per group. Three independent experiments were performed. (d) Blood cells were depleted of erythrocytes. The bars indicate the frequency of eosinophils in DT- and PBS-treated mice upon exposure to PBS or LPS. Data are expressed as mean±s.e.m.; n=7 mice per group pooled from two experiments.
We next investigated whether exposure to LPS modified the surface phenotype of lung DCs or their ability to induce cytokine secretion by effector Th2 cells. Both langerin+ and CD11b+ DCs were purified from the lungs of LPS- or PBS-exposed postasthmatic mice and either analyzed by FACS or incubated with LACK-specific WT15 Th2 cells. Although the phenotype of CD11b+ DCs remained unchanged upon LPS exposure, langerin+ DCs upregulated OX40 ligand and to a smaller extent CD80 (Supplementary Figure S1 online). Whether they were purified from PBS- or LPS-exposed mice, both langerin+ and CD11b+ DC subsets induced Th2 effector cells to secrete cytokines (Supplementary Figure S2 online). It is noteworthy that langerin+ DCs purified from LPS-exposed mice were less potent at inducing IL-10 secretion by Th2 cells when compared with those purified from PBS-exposed mice.
As LPS induced langerin+ DCs to promote Th2 differentiation in vitro and as eosinophil mobilization is dependent on the Th2 cytokine IL-5, we investigated the effect of langerin+ DC depletion on T-cell activation and eosinophil appearance. To this aim, we used langerin-DTR mice that express the human diphtheria toxin receptor (DTR) under the langerin promoter.21 When DT was administered i.n. to these mice, there was a strong reduction in the number of CD11b− langerin+ DCs in lungs whereas CD11b+ DCs and alveolar macrophages were not affected (Supplementary Figure S3 online). Postasthmatic langerin-DTR mice were treated or not with DT, injected with CFSE-labeled WT15 TCR transgenic T cells, and exposed to LPS or PBS. In agreement with the results obtained in non-transgenic postasthmatic mice, the frequency of LACK-specific T cells that had gone through ⩾5 divisions after 3 days increased from 22.0±2.0% in PBS-exposed mice to 56.7±4.4% in LPS-exposed mice (Figure 5c, left panels). In striking contrast, T-cell proliferation was not observed in DT-treated langerin-DTR mice irrespective of whether they were exposed to PBS or LPS (Figure 5c, right panels).
Most importantly, the increased frequency of eosinophils that was induced by LPS in postasthmatic mice was abolished upon DT treatment (Figure 5d). Taken together, our data demonstrate that langerin+ DCs are solely responsible for the presentation of LACK that occurs in the MLNs of postasthmatic mice, both in the steady state and upon exposure to a low dose of LPS. Furthermore, langerin+ DCs are responsible for the reactivation of LACK-specific T cells and for the subsequent recruitment of eosinophils that occurs upon LPS exposure of postasthmatic mice.
Discussion
In this paper, we have shown that postasthmatic mice that have previously experienced an acute episode of allergic asthma exhibited allergen-loaded DCs in their lungs. In a previous report, we found that bulk populations prepared from the BALF of postasthmatic mice, but not those prepared from the lung or the MLNs, stimulated LACK-specific T cells.16 In apparent contrast with this previous study, we have now found that purified DCs from both the lung and the MLNs of postasthmatic mice stimulated LACK-specific T cells. Our failure to detect antigen presentation in the lung and the draining LNs in our previous study was likely because of the fact that bulk populations, and not purified populations, were tested. Therefore, infrequent populations of antigen-presenting cells could have been diluted out with other cells, precluding their detection. Our present findings confirm the presence of antigenic reservoirs in the lung as previously reported,25 and further identify these reservoirs as particular DC subpopulations. Indeed, two types of lung DCs induced LACK-specific T-cell proliferation in vitro without the addition of exogenous antigen, langerin+ DCs and CD11b+ DCs, with the former being much more potent than the latter. In contrast to langerin+ DCs, CD11b+ DCs were not capable of inducing naive T cells to secrete large amounts of IFN-γ, IL-5, IL-13, or IL-10 in vitro. However, these two DC subsets were equally competent at inducing Th2 cytokine release from already differentiated Th2 cells. Interestingly, Th2 cells produced less IL-10 and slightly more IL-13 upon incubation with DCs from LPS-exposed mice than upon incubation with DCs from PBS-exposed mice. In addition, we have found that OX40 ligand was upregulated on the cell surface of langerin+ DCs upon LPS exposure. As suggested by other studies,26, 27, 28 this molecule together with other cell surface molecules might be responsible for the increased ability of langerin+ DCs to trigger inflammatory Th2 differentiation.
Our results obtained with langerin-DTR mice suggested that only the airway langerin+ DCs presented LACK to the CD4+ T cells. This result is reminiscent with the data obtained by GeurtsvanKessel et al.29 that recently found that CD11b+ DC cells purified from the MLNs of influenza-infected mice did not present antigen to naive CD4+ or CD8+ T cells despite containing viral antigens. Therefore, one can question the role of the lung CD11b+ DCs. Indeed, CD11b+ DCs have been found to be interacting with leukocytes in the peribronchial and perivascular infiltrated areas,30 where they massively produced inflammatory chemokines,31, 32 and recent experiments demonstrated that selective depletion of CD11b+ myeloid cells in lungs abrogated the production of the Th2-cell-attracting inflammatory chemokines, CCL17, CCL22, CCL11, and Th2 lymphocyte, and eosinophil recruitment into the airways of sensitized and challenged mice.32 Therefore, CD11b+ DC cells may attract effector CD4+ T cells to the lungs.
In contrast, we have shown here that langerin+ DCs, previously described to be localized at the basolateral side of bronchial epithelial cells,30, 33 continuously migrate from the lungs to the MLNs where they prime and induce the proliferation of allergen-specific CD4+ T cells, therefore suggesting that langerin+ DCs are mucosal DCs that sampled the antigen from the airways and migrate to the LNs. Upon LPS exposure, langerin+ DCs migrate to the MLNs at a faster rate, reactivate allergic-specific memory CD4+ T cells, and instruct naive CD4+ T cells to differentiate into Th2 cells. This cascade of events leads to the reactivation of the disease as evidenced by the recruitment of eosinophils to the airways and by AHR. Although lung langerin+ DCs were identified few years ago,23 no study has assessed their role in allergic airway inflammation. A recent study has elegantly demonstrated that the clearance of the influenza virus is dependent on langerin+ lung DCs, and that depletion of these cells results in reduced number of virus-specific CD8+ T cells in the MLNs.29 However, the ability of langerin+ DCs to induce CD4+ T-cell differentiation was not addressed in this report.
del Rio et al.19 have monitored the presence of OVA-loaded DCs in the MLNs of C57BL/6 following the intratracheal injection of fluorochrome-conjugated OVA. The ability of these DCs to prime OVA-specific CD4+ and CD8+ TCR transgenic T cells was also investigated. Although OVA was found in both CD103+ and CD103− LN DCs, CD103− but not CD103+ DCs could prime CD4+ T cells. As langerin+ DCs also express CD103, this latter result is in apparent contrast with our finding that lung langerin+ DCs are very potent at inducing CD4+ T-cell proliferation. Several hypotheses could explain the apparent discrepancy between our data and those reported by del Rio et al.19 First, these authors reported that CD103+ DCs expressed langerin mRNA but could not rule out that a small fraction of these cells did not express langerin. Indeed, although all lung langerin+ DCs were CD103+, we found that 8–10% of these cells did not express langerin in langerin-GFP reporter mice (Supplementary Figure S4 online). Second, our experiments were performed with lung DCs, whereas del Rio et al.19 have used CD103+ DCs from the draining LNs. Third, our experiments were performed using DCs purified from postasthmatic mice, whereas del Rio et al.19 have used naive animals. Last, we have delivered 40 μg of LACK and analyzed the mice 3 weeks later, whereas del Rio et al.19 have delivered 60 μg of OVA and analyzed the mice 24 h later.
Bedoret et al.34 have described a population of interstitial macrophages that prevent sensitization to harmless antigens in the presence of LPS. Investigating the role of these F4/80+ CD11c− cells in our model could be of interest, although technically difficult. In very preliminary experiments, we have tried to deplete alveolar macrophages (including interstitial macrophages) in postasthmatic mice using chlorydrate liposomes. This treatment did not alter the proliferation of adoptively transferred antigen-specific T cells, suggesting that macrophages did not have a role in the regulation of T-cell proliferation, at least in the steady state.
Here, we have demonstrated that langerin+ DCs are able to prime naive CD4+ T cells both in vitro and in vivo. Most importantly, whereas langerin+ DCs from PBS-exposed postasthmatic mice induce a Th1 differentiation, they promote Th2 responses in postasthmatic mice exposed to LPS, therefore providing a possible explanation for the subclinical worsening of asthmatic patients exposed to a low dose of LPS. It remains to be determined whether LPS acts directly on langerin+ DCs, or indirectly through other cell types such as epithelial cells, as recently demonstrated for the house dust mite allergen.14 Although our studies shed light into the role of langerin+ DCs during LPS reactivation, their role during sensitization and the acute asthma responses remains to be elucidated.
Methods
Mice. BALB/c ByJ mice were purchased from The Centre d’Elevage Janvier (France) and housed under specific pathogen-free conditions in our animal facility. OVA-specific DO11.10 and LACK-specific WT15 TCR transgenic BALB/c mice were previously described.35 In this study, WT15 and DO11.10 transgenic mice were further crossed onto RAG-1−/− Thy1.1+/+ BALB/c mice and onto RAG-1−/− Thy1.1−/− BALB/c, respectively. Langerin-DTR and langerin-GFP21 were previously described. In this study, langerin-DTR and langerin-GFP mice on the C57BL/6 background were crossed once onto BALB/c mice to generate F1 mice.
Asthma provocation and LPS exposure. Mice were sensitized intraperitoneally with 10 μg of LACK in 2 mg aluminum hydroxide (Pierce, Perbio Science, Brebieres, France) in PBS at days −17 and −10. Sensitized mice were anesthetized at day 0 with 0.1% ketamine (Imalgene; Merial, Lyon, France), 0.1% xylazyne (Rompun; Bayer), and received 40 μg of LACK i.n. At day 21, mice were anesthetized as indicated above and received 500 ng LPS from Escherichia coli 011:B4 (Invivogen, Toulouse, France) or PBS i.n. In the indicated experiments, postasthmatic mice were injected with anti-CD4 monoclonal antibodies (250 μg, GK1.5 clone) twice at days 15 and 18. When indicated, PBS and LPS shots included 1 μl of phycoerythrin fluorescent 0.5 μm latex beads (Sigma, Lyon, France). Mice were killed 3 or 7 days later for BALF and WT15T cell proliferation analyses.
AHR. Invasive measurements of dynamic lung resistance and compliance were performed 5 days after challenge using a Flexivent apparatus (SCIREQ, Emka Technologies, France) as previously described.36 For noninvasive measurements, mice were analyzed at the indicated days using whole-body plethysmography.37
DT treatment of langerin-DTR mice. DT was administered once i.n. at a dose of 20 ng g–1, 16 days after LACK challenge. PBS or LPS administration to DT-treated mice was done 5 days afterward, totaling 21 days after LACK challenge.
Tissue processing. Lungs, MLNs, or spleens were cut to small pieces in Hanks’ buffered salt solution containing 400 U type I collagenase and 1 mg ml–1 DNAse I and digested for 30 min at 37 °C. Cells were strained through a 70-μm cell strainer. Erythrocytes were lysed with ACK lysis buffer.
Cell enrichment and sorting. CD11c+ enrichment of lung cells was performed using anti-CD11c+ magnetic beads (Miltenyi, Paris, France). For cell sorting, CD11c+-enriched populations were labeled with antibodies to CD11c and CD11b and sorted on a FACSVantage cell sorter, BD Biosciences, Le Pont De Claix, France. In some experiments, postasthmatic langerin-GFP mice were exposed to LPS or PBS at 21 days after LACK challenge. After 3 days, CD11c+ were enriched from lungs, stained with antibodies to CD11c and CD11b, and sorted after gating on CD11c+GFP+ or CD11c+CD11b+ non-autofluorescent cells. CD4+ T-cell enrichment was performed using anti-CD4+ magnetic beads (Miltenyi).
FACS. For differential BALF cell counts, cells were stained with monoclonal antibodies to CCR3 (R&D Systems, Lille, France), Gr-1, TCR-β, and CD45R (BD Biosciences). Eosinophils were defined as CCR3+ TCR-β−CD45R−Gr-1med and lymphocytes as TCR-β+CD45R+. For characterization of CD11c+-enriched lung cells, anti-CD11c, -CD11b, -Ly6C, -IA/IE, -F4/80, -CD45R, -CD8α, -CD40, -CD80, -CD86, -CD103, -CD207, -OX40 ligand, and -DEC205 monoclonal antibodies (BD Biosciences) were used. Cells were analyzed using FACSCalibur, or LSR II Fortessa cytometers and CellQuestPro or DIVA softwares (BD Bioscience).
Cell transfer. CD4+ spleen T cells were prepared from naive WT15 TCR transgenic mice, stained with CFSE,38 and 2 × 106 WT15 cells were injected intraperitoneally. Donor cells were identified by CFSE-labeling and/or Thy1.1 expression.
Cell culture. For WT15 TCR transgenic CD4+ T-cell proliferation in vitro assays, WT15 CD4+ T cells were CFSE labeled and 105 T cells were incubated with 105 CD11b+ or 105 langerin+ DCs in 96-well flat-bottom plates in complete RPMI. T-cell proliferation was determined by CFSE dilution after 3 or 7 days. For 7-day-long experiments, fresh media was replenished after 3 days. Alternatively, 104 bead+ DCs from PBS- or LPS-exposed mice were used. All assays were performed in triplicates.
Multiplex cytokine analysis. After 7 days in culture, 50 ng ml–1 phorbol myristate acetate (Sigma) was added and supernatants were recovered after 6 h. Detection of IFN-γ, IL-5, IL-13, IL-4, and IL-10 was carried out by Cytometric Bead Array using a FACSArray (BD Bioscience).
LACK preparation. LACK recombinant protein was produced in E. coli, purified as described,39 and detoxified using an Endotrap column (Profos, Regensburg, Germany). Endotoxin levels assessed by LAL assay (Pierce) were below 5 ng mg–1 of protein.
LACK-specific IgE. Serum LACK-specific IgE was quantified as described.36
Statistical analysis. Analysis of variance for repeated measures was used to determine the levels of difference between groups of mice for plethysmography measurements. Comparisons for all pairs were performed using unpaired two-tailed Student's t-test. Significance levels were set at a P-value of 0.05.
References
Wills-Karp, M. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu. Rev. Immunol. 17, 255–281 (1999).
Cookson, W. The alliance of genes and environment in asthma and allergy. Nature 402, B5–B11 (1999).
Gereda, J.E. et al. Relation between house-dust endotoxin exposure, type 1 T-cell development, and allergen sensitisation in infants at high risk of asthma. Lancet 355, 1680–1683 (2000).
Braun, M.C., He, J., Wu, C.Y. & Kelsall, B.L. Cholera toxin suppresses interleukin (IL)-12 production and IL-12 receptor beta1 and beta2 chain expression [In Process Citation]. J. Exp. Med. 189, 541–552 (1999).
Thorne, P.S. et al. Endotoxin exposure is a risk factor for asthma: the national survey of endotoxin in United States housing. Am. J. Respir. Crit. Care. Med. 172, 1371–1377 (2005).
Michel, O. et al. Domestic endotoxin exposure and clinical severity of asthma. Clin. Exp. Allergy 21, 441–448 (1991).
Michel, O. et al. Severity of asthma is related to endotoxin in house dust. Am. J. Respir. Crit. Care. Med. 154, 1641–1646 (1996).
Michel, O., Duchateau, J. & Sergysels, R. Effect of inhaled endotoxin on bronchial reactivity in asthmatic and normal subjects. J. Appl. Physiol. 66, 1059–1064 (1989).
Michel, O. et al. Inflammatory response to acute inhalation of endotoxin in asthmatic patients. Am. Rev. Respir. Dis. 146, 352–357 (1992).
Eisenbarth, S.C. et al. Lipopolysaccharide-enhanced, Toll-like receptor 4-dependent T helper cell type 2 responses to inhaled antigen. J. Exp. Med. 196, 1645–1651 (2002).
Duan, W., So, T. & Croft, M. Antagonism of airway tolerance by endotoxin/lipopolysaccharide through promoting OX40L and suppressing antigen-specific Foxp3+ T regulatory cells. J. Immunol. 181, 8650–8659 (2008).
Murakami, D. et al. Lipopolysaccharide inhalation exacerbates allergic airway inflammation by activating mast cells and promoting Th2 responses. Clin. Exp. Allergy 37, 339–347 (2007).
Stephens, R. & Chaplin, D.D. IgE cross-linking or lipopolysaccharide treatment induces recruitment of Th2 cells to the lung in the absence of specific antigen. J. Immunol. 169, 5468–5476 (2002).
Hammad, H. et al. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nat. Med. 15, 410–416 (2009).
Trompette, A. et al. Allergenicity resulting from functional mimicry of a Toll-like receptor complex protein. Nature 457, 585–588 (2009).
Julia, V. et al. A restricted subset of dendritic cells captures airborne antigens and remains able to activate specific T cells long after antigen exposure. Immunity 16, 271–283 (2002).
Jakubzick, C. et al. Lymph-migrating, tissue-derived dendritic cells are minor constituents within steady-state lymph nodes. J. Exp. Med. 205, 2839–2850 (2008).
Vermaelen, K.Y., Carro-Muino, I., Lambrecht, B.N. & Pauwels, R. A. Specific migratory dendritic cells rapidly transport antigen from the airways to the thoracic lymph nodes. J. Exp. Med. 193, 51–60 (2001).
del Rio, M.L., Rodriguez-Barbosa, J.I., Kremmer, E. & Forster, R. CD103- and CD103+ bronchial lymph node dendritic cells are specialized in presenting and cross-presenting innocuous antigen to CD4+ and CD8+ T cells. J. Immunol. 178, 6861–6866 (2007).
Hintzen, G. et al. Induction of tolerance to innocuous inhaled antigen relies on a CCR7-dependent dendritic cell-mediated antigen transport to the bronchial lymph node. J. Immunol. 177, 7346–7354 (2006).
Kissenpfennig, A. et al. Dynamics and function of Langerhans cells in vivo: dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells. Immunity 22, 643–654 (2005).
Kirby, A.C., Raynes, J.G. & Kaye, P.M. CD11b regulates recruitment of alveolar macrophages but not pulmonary dendritic cells after pneumococcal challenge. J. Infect. Dis. 193, 205–213 (2006).
Sung, S.S. et al. A major lung CD103 (alphaE)-beta7 integrin-positive epithelial dendritic cell population expressing Langerin and tight junction proteins. J. Immunol. 176, 2161–2172 (2006).
Vermaelen, K. & Pauwels, R. Accurate and simple discrimination of mouse pulmonary dendritic cell and macrophage populations by flow cytometry: methodology and new insights. Cytometry 61A, 170–177 (2004).
Jelley-Gibbs, D.M et al. Persistent depots of influenza antigen fail to induce a cytotoxic CD8 T cell response. J. Immunol. 178, 7563–7570 (2007).
Akiba, H. et al. Critical contribution of OX40 ligand to T helper cell type 2 differentiation in experimental leishmaniasis. J. Exp. Med. 191, 375–380 (2000).
Ito, T. et al. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J. Exp. Med. 202, 1213–1223 (2005).
Seshasayee, D. et al. In vivo blockade of OX40 ligand inhibits thymic stromal lymphopoietin driven atopic inflammation. J. Clin. Invest. 117, 3868–3878 (2007).
GeurtsvanKessel, C.H. et al. Clearance of influenza virus from the lung depends on migratory langerin+CD11b- but not plasmacytoid dendritic cells. J. Exp. Med. 205, 1621–1634 (2008).
GeurtsvanKessel, C.H. & Lambrecht, B.N. Division of labor between dendritic cell subsets of the lung. Mucosal. Immunol. 1, 442–450 (2008).
Beaty, S.R., Rose, C.E. Jr & Sung, S.S. Diverse and potent chemokine production by lung CD11bhigh dendritic cells in homeostasis and in allergic lung inflammation. J. Immunol. 178, 1882–1895 (2007).
Medoff, B.D et al. CD11b+ myeloid cells are the key mediators of Th2 cell homing into the airway in allergic inflammation. J. Immunol. 182, 623–635 (2009).
Sung, S.J., Taketomi, E.A., Smith, A.M., Platts, M.T. & Fu, S.M. Efficient presentation of house dust mite allergen Der p 2 by monocyte- derived dendritic cells and the role of beta 2 integrins. Scand. J. Immunol. 49, 96–105 (1999).
Bedoret, D. et al. Lung interstitial macrophages alter dendritic cell functions to prevent airway allergy in mice. J. Clin. Invest. 119, 3723–3738 (2009).
Wang, Q. et al. CD4 promotes breadth in the TCR repertoire. J. Immunol. 167, 4311–4320 (2001).
Verhasselt, V. et al. Breast milk-mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat. Med. 14, 170–175 (2008).
Hamelmann, E. et al. Noninvasive measurement of airway responsiveness in allergic mice using barometric plethysmography. Am. J. Respir. Crit. Care. Med. 156, 766–775 (1997).
Quah, B.J., Warren, H.S. & Parish, C.R Monitoring lymphocyte proliferation in vitro and in vivo with the intracellular fluorescent dye carboxyfluorescein diacetate succinimidyl ester. Nat. Protoc. 2, 2049–2056 (2007).
Mougneau, E. et al. Expression cloning of a Leishmania major protective T-cell antigen. Science 268, 563–566 (1995).
Acknowledgements
We thank Frank Aguila for his help with figures, and Nicolas Guy, Véronique Thieffin, and Alain Barbot for their excellent animal care. This work was supported by grants from the Agence Nationale de la Recherche (ANR) (to V.J.) and from the Fondation pour la Recherche Médicale (FRM) (to V.J.). A.O.S was supported by a grant from the FRM.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Rights and permissions
About this article
Cite this article
Ortiz-Stern, A., Kanda, A., Mionnet, C. et al. Langerin+ dendritic cells are responsible for LPS-induced reactivation of allergen-specific Th2 responses in postasthmatic mice. Mucosal Immunol 4, 343–353 (2011). https://doi.org/10.1038/mi.2010.73
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2010.73
This article is cited by
-
The kinase p38α functions in dendritic cells to regulate Th2-cell differentiation and allergic inflammation
Cellular & Molecular Immunology (2022)
-
Eosinophil recruitment is dynamically regulated by interplay among lung dendritic cell subsets after allergen challenge
Nature Communications (2018)
-
Potential autocrine regulation of interleukin-33/ST2 signaling of dendritic cells in allergic inflammation
Mucosal Immunology (2013)
-
Pulmonary CD103+ dendritic cells prime Th2 responses to inhaled allergens
Mucosal Immunology (2012)

{kind=link}
{kind=link}
{kind=link}