
Abstract
Variants in the ABCA4 gene are the most common cause of juvenile-onset blindness affecting close to 1 in 10 000 people worldwide. Disease severity varies largely according to genotype, which can be specific to ethnic and racial groups. Here we investigate the spectrum of ABCA4 variation and its phenotypic expression in 38 patients of South Asian descent, notably from India, Pakistan, Bangladesh and Sri Lanka. Sequencing of all exons and flanking intronic sequences of ABCA4 revealed disease-causing variants in all patients: 3 in 2.6%, 2 in 81.6% and 1 in 15.8%. Altogether, 36 distinct variants were identified, including 9 previously not described. The most frequent variant c.5882G>A, p.(G1961E) was found in half the patients, the highest ever reported in a single study cohort. The South Asian founder variant c.859-9T>C was identified along with other founder variants ascribed to Danish, Chinese, Mexican and African patients. Patients carrying c.5882G>A, p.(G1961E) exhibited a consistently confined disease phenotype, normal quantitative autofluorescence (qAF) levels and preserved full-field ERG (ffERG) while c.859-9T>C resulted in widespread disease, significantly elevated qAF and reduced to non-detectable ffERG. South Asian patients present with a relatively unique ABCA4 profile comprised of various ethnic founder variants resulting in two or three major retinal phenotypes.
Similar content being viewed by others
Introduction
The ABCA4-associated disease spectrum encompasses large, heterogeneous groups of recessive retinal dystrophies including Stargardt disease1 (STGD1; MIM 248200), cone-rod dystrophy (CRD)2, 3 and retinitis pigmentosa-like phenotypes.2, 4, 5 Genotypic heterogeneity also exists within these groups as currently over 800 ABCA4 (MIM 601691, NM_000350.2) variants have been identified.6 Although precise genotype–phenotype correlations amongst even the most frequent variants are scarcely understood, the ethnic and geographic origin of several variants have been described. Founder variants broadly attributed to the Western European population include c.2588G>C, p.[(G863A,G863del)];7 although more specifically, variants associated with specific ethnicities include the c.[1622T>C;3113C>T], p.[L541P;A1038V] complex allele in individuals of predominantly German descent,3, 8 c.3386G>T, p.(R1129L) in individuals of Spanish descent9 and c.2894A>G, p.(N965S) in individuals of Danish descent.10 Populations outside of Europe are comparatively less well-characterized; however, another founder variant, c.5318C>T, p.(A1773V), was recently identified in Mexico11 and the c.6320G>A, p.(R2107H) variant was found to be significantly over-represented in a large cohort of African-American patients diagnosed with STGD1.12 Rigorous analyses of disease phenotypes corresponding to these ethnicity-specific variants are limited with the exception of p.[L541P;A1038V], which was shown to impart an accelerated onset of retinopathy in patients13 and rapid accumulation of bisretinoid lipofuscin in the RPE in the homozygous knock-in mouse model (AbcaPV/PV).14 Increased lipofuscin accumulation in patients with p.[L541P;A1038V] has also been demonstrated by quantitative autofluorescence (qAF) imaging in patients,15 though only when p.[L541P;A1038V] is not in combination with the c.5882G>A, p.(G1961E) variant, which itself contrastingly results in a less pronounced accumulation of lipofuscin and the absence of a dark choroid on fluorescein angiography.15, 16 In reporting the c.6320G>A, p.(R2107H) variant in Africa Americans, we also noted that patients in this cohort experienced a significantly later onset and milder severity of disease symptoms as compared to affected Caucasian patients.12
The proper management of patients harboring genetically well-characterized disease alleles in ABCA4 is inadequate without a comprehensive understanding of their corresponding disease phenotypes. Recently, we reported an intronic variant, c.859-9T>C, that is highly frequent in Asian Indian patients (ethnic descent from Pakistan, India and Bangladesh) but rare in the ethnically matched general population.17 This study describes the phenotypic spectrum and genotypic composition of South Asian patients diagnosed with ABCA4-associated disease.
Materials and methods
Patients and clinical evaluation
All study subjects were consented before participating in the study under the protocols #AAAI9906 approved by the Institutional Review Board at Columbia University and #20130770 by the Western Institutional Review Board at the Chicago Lighthouse. The study adhered to tenets established in the Declaration of Helsinki. Complete ophthalmic examinations were provided by a retinal specialist (SHT, GF, SB), including slit-lamp and dilated fundus examinations. Vision was assessed by the measurement of best-corrected visual acuity (BCVA; Snellen or Feinbloom charts), while further clinical assessments were made from color fundus photographs, 488 and 787 nm excitation autofluorescence images and spectral domain-optical coherence tomography scans and full-field electroretinogram results.
Clinical data acquisition and analysis
Spectral domain-optical coherence tomography (SD-OCT) scans and corresponding infrared reflectance fundus images were acquired using a Spectralis HRA+OCT (or HRA+OCT; Heidelberg Engineering, Heidelberg, Germany). Fundus autofluorescence images were obtained using a confocal scanning laser ophthalmoscope (cSLO, Heidelberg Retina Angiograph 2, Heidelberg Engineering, Dossenheim, Germany) Fundus AF images were acquired by illuminating the fundus with an argon laser source (488 nm excitation) and viewing the resultant fluorescence through a band pass filter with a short wavelength cutoff at 495 nm. Color fundus photos were obtained with a FF 450plus Fundus Camera (Carl Zeiss Meditec AG, Jena, Germany) and CR-1 Mark II Fundus Camera (Canon, Tokyo, Japan).
Full-field electroretinograms (ffERG), available in 12 patients, were recorded using the Diagnosys Espion Electrophysiology System (Diagnosys LLC, Littleton, MA, USA). Prior to acquisition, pupils were maximally dilated and measured before ffERG recording using tropicamide (1%) and phenylephrine hydrochloride (2.5%); and the corneas were anesthetized with proparacaine 0.5%. Silver impregnated fiber electrodes (DTL; Diagnosys LLC, Littleton, MA) and Burian-Allen contact lens were used with a ground electrode on the forehead. Normative amplitude and implicit time ranges for each stimulus were provided by the Diagnosys system software. All procedures were performed using extended testing protocols outlined by the International Society for Clinical Electrophysiology of Vision standard.18
Quantitative autofluorescence (qAF) was performed and analyzed in 25 eyes of 14 patients (Table 1). Protocols for the acquisition of AF images that meet the quality standards necessary for quantification have been previously described.15, 19 Fundus AF images (30°; 488 nm excitation) were acquired using a confocal scanning laser ophthalmoscope (Spectralis HRA+OCT; Heidelberg Engineering, Heidelberg, Germany) modified by the insertion of an internal fluorescent reference to account for variations in laser power and detector gain. The barrier filter in the device transmitted light from 500 to 680 nm. Prior to acquisition, the fundus was exposed to the AF light for 20 to 30 s to bleach rhodopsin, while at the same time, focus and alignment were refined to produce a maximum and uniform signal over the entire field.
Fundus AF images were analyzed with a dedicated image analysis software written in IGOR (WaveMetrics, Lake Oswego, OR, USA) to determine qAF. The software recorded the mean GLs of the internal reference and of eight circularly arranged segments positioned at an eccentricity of approximately 7° to 9°. Segments were scaled to the horizontal distance between the fovea and the temporal edge of the optic disc. Control values used in this study consisted of previously published data from 277 healthy subjects (374 eyes; age range, 5–60 years) without a family history of retinal dystrophy.20
Sequencing and variant analyses
All 50 exons and exon–intron boundaries of the ABCA4 gene (MIM 601691, NM_000350.2) were amplified using Illumina TruSeq Custom Amplicon protocol (Illumina, San Diego, CA, USA), followed by sequencing on Illumina MiSeq platform. The next-generation sequencing reads were analyzed and compared to the ABCA4 reference sequence NG 009073.1, using the variant discovery software NextGENe (SoftGenetics LLC, State College, PA, USA). All detected possibly disease-associated variants were confirmed by Sanger sequencing. Nucleotide positions and protein translation correspond to CCDS747.1 and NP 000341.2, respectively. Nucleotide numbering uses the A of the ATG translation initiation start site as nucleotide 1. The allele frequencies of all variants were compared to the Exome Aggregation Consortium (ExAC) database (Cambridge, MA, USA; http://exac.broadinstitute.org; accessed September 2016). All ABCA4 variants and patients reported in this manuscript were submitted to the Leiden Open Variation Database 3.0 (http://www.lovd.nl/ABCA4). Segregation of the new variants with the disease was analyzed in families if family members were available (Table 1). The possible effect of all ABCA4 variants was assessed using the combination of following prediction programs (Table 2): Polyphen-2,21 Align-GVGD,22 SIFT,23 MutationTaster,24 SpliceSiteFinder,25 MaxEntScan,26 NNSPLICE,27 GeneSplicer,28 and Human Splicing Finder.29 All of these algorithms except for Polyphen-2 were accessed via Alamut software version 2.2 (Interactive Biosoftware, Rouen, France; http://www.interactive-biosoftware.com), using automated computation of this software version. Polyphen-2 results were retrieved from the single entry Web form (http://genetics.bwh.harvard.edu/pph2/) with the HumDiv-model and version 2.2.2 of the software. Predictions are included as supporting data to the conclusions that are based on different frequencies of the variants between the patients and general population.
Results
All patients included in the study (n=38) are of South Asian descent by both, or either maternal or paternal lineage. Thirty-three of the patients were bi-parentally Indian, (n=27, 71%), Pakistani (n=4, 11%), Sri Lankan (n=1) and Bangladeshi (n=1). The remaining five patients were Indian by one parent and either African, Chinese, Italian or Moroccan by the other. The cohort also consists of two familial pairs, P25-P21 (parent–child) and P37–P38 (siblings). Demographic and clinical characteristics are summarized in Table 1. Mean cohort age at clinical presentation was 35 years but ranged widely between 9 to 65 years. Disease onset also varied within the cohort with 16% reporting visual symptoms in the first decade, 39% in the second, 18% in the third, 13% in the fourth and 11% in the fifth and one patient (P30) in the sixth. Measured BCVAs ranged from 20/20 to HM (Feinbloom BCVA was measured in 5 patients, which ranged from 10/63-2 to 10/160).
Analysis of ABCA4 variants in South Asian patients
Following direct sequencing of ABCA4, disease-causing alleles were found in all patients: two in 81%, three in 3% and one in 16%. Nine unrelated individuals were homozygous. A total of 36 distinct ABCA4 variants were identified in the cohort (Table 2). All variants were mostly predicted deleterious with the exception of one missense variant, c.4217A>G, p.(H1406R), which had predicted minimal biochemical (Align GVGD) or functional consequence (Polyphen-2 and MutationTaster). In total, 11 variants were predicted to negatively affect exon splicing resulting in the creation, deletion or reduction of donor and acceptor sites. Half of the patients harbored the c.5882G>A, p.(G1961E) allele, which is consistent with the population frequency of the allele (~1.5%) in the South Asians. The intronic variant, c.859-9T>C, previously associated with patients of Asian Indian descent was seen in five patients and is predicted to affect splicing. The nine novel variants identified were c.66G>A, c.571-2_ 575delAGTTCGC, c.3407G>T (p.(G1136V)), c.5243G>A (p.(G1748E)), c.5313-2_ 5316delAGATGG, c.6098T>A (p.(L2033H)), c.6191C>T (p.(A2064V)), c.6317G>A (p.(R2106H)) and c.6479+1G>A (Table 2).
Retinal phenotypes
Clinically, all patients exhibited phenotypes consistent with the described spectrum for ABCA4-associated disease from mild to very severe. Classically associated features were observed in autofluorescence imaging including central lesions of discrete atrophy or hypoAF mottling accompanied by bright pisciform flecks arranged in a confluent pattern across the posterior pole and a notable sparing of the peripapillary region. The most severe, end-stage phenotypes, characterized by widespread RPE wipeout across the posterior pole, were also observed in both older (P20, age 62 years) and younger patients (P7, age 34 years).
Genotype–phenotype correlations
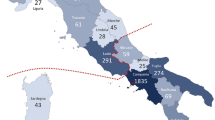
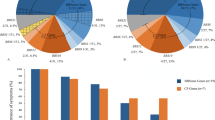
All patients harboring the c.5882G>A, p.(G1961E) variant (n=19) (age range, 9–51 years) presented with bull’s eye lesions or discrete geographic atrophy and very few, locally confined flecks, which never extended beyond the macula (Figure 1). The five patients (P4, P6, P16, P30 and P31) with only bull’s eye lesions and no visible flecks were also in the p.(G1961E) group. Patients harboring the c.859-9T>C exhibited moderately severe to very advanced phenotypes (Figure 2). P10 (aged 33 years), for instance, presented with a bleached-white appearance, multiple regions of atrophy across the posterior pole on autofluorescence imaging and an arc of densely accumulated bone-spicule pigmentation in the mid-periphery along the superior vascular arcades. This patient harbored 3 ABCA4 variants, including a splicing-affecting variant c.66G>A, which was found to be in phase with c.859-9T>C, and the stop mutation c.6658C>T, p.(Q2220*) on the second allele. Differences in quantitative autofluorescence (qAF) levels analyzed in 14 patients further distinguished genotypic groups. Overall, both elevated and normal qAF values were measured in affected patients. The majority of p.(G1961E) eyes fell within the 95% confidence interval for healthy eyes while P14 and P18 who were homozygous for c.859-9T>C were significantly elevated with respect to the other genotypes. Color maps of qAF illustrate the difference in autofluorescence in the macula between age-matched healthy control eye, a c.859-9T>C (P18) and c.5882G>A, p.(G1961E) (P21) eye (Figure 3). Scotopic, maximal, 30 Hz flicker and single flash photopic ffERG recordings were variable across the cohort (Figure 4). Severe attenuation of both the rod and cone systems were observed in, P7, P20, and notably P10 and P9 of the c.859-9T>C group while photopic and, especially, rod responses were generally unaffected in p.(G1961E) patients.

Retinal phenotype on color photographs, autofluorescence imaging and spectral domain-optical coherence tomography in South Asian patients harboring the c.5882G>A, p.(G1961E) allele of ABCA4. In total, 18 of 36 unrelated cases within the cohort harbored the p.(G1961E) variant resulting phenotype characterized by central bull’s eye lesions and no isolated flecks outside of the lesion (a, b), Confined flecks lesions within the macula (c, d) and early discrete areas of geographic atrophy with flecks distributed around the lesion (e, f).

Moderate to severe phenotypes were associated with the c.859-9T>C founder allele of ABCA4. Color photography (a) and autofluorescence imaging (b) in the most advanced case who was compound heterozygous for c.859-9T>C and c.66G>A, p.(K22=) (P10) exhibited severe atrophy of the posterior pole and a confluent pattern of bone-spicule pigment migration along the superior arcades. Also presented are two homozygous c.859-9T>C cases with central atrophy extra-macular flecks (c, d) and progressively advanced lesions with densely accumulated flecks (e, f).

Quantitative autofluorescence (qAF) in South Asian patients with ABCA4 disease. A qAF value was calculated by averaging the values of 8 scaled segments in the macula. Analysis segments and color-coded maps corresponding to qAF-unit scale (0–1200) in an age- and ethnicity-matched healthy control (a) and P18 (c.859-9T>C homozygous) (b) and P21(p.[(G1961E; R2149*)]) (c). qAF versus age plot of 14 patients (25 eyes) grouped according to genotypes with the mean (solid black line) and 95% confidence intervals (dotted black lines) of 277 healthy subjects (374 eyes) (d).

Full-field electroretinogram results in 12 South Asian patients with ABCA4 disease. Scotopic, maximal, 30 Hz flicker and photopic scans of the right eye of 12 patients with segregated plots for c.5882G>A, p.(G1961E) and the founder variant c.859-9T>C. Peak amplitude and implicit time ranges for healthy eyes are delineated by the dashed line boxes.
Discussion
The proportion of inherited retinal disorders in the Indian subcontinent has yielded diverse findings that consist predominantly of retinitis pigmentosa (TULP1, NR2E3, MFRP,30 EYS,31 FAM161A,32 AIPL1, RPE65, GUCY2D, CRB1, RDH12, IQCB1 and SPATA733 and to a lesser extent, STGD1,34, 35 among others. Although the frequency of ABCA4 disease alleles in this population is unknown, two reports have described novel variants segregating in several affected families34, 35 along with the c.859-9T>C variant identified by our group.17 The aim of the current study is to provide a comprehensive genetic and clinical profile of ABCA4-associated diseases in this population.
Overall, the clinical spectrum of this cohort exhibited overlapping characteristics with patients of other ethnicities but also varied in accordance with an individual patient’s respective genotype. The detection rate of variants in this cohort (at least two disease-causing variants in 84%) was higher than what has been previously reported from our group in other ethnicities.12, 17 Nine unrelated cases were found to be homozygous, which was not unexpected due to the reportedly high incidence of consanguinity resulting in retinal disease in this population.36, 37 The most severe variant combinations were found in P7 (homozygous nonsense mutations) and P10 (3 total: two intronic/splice and one nonsense mutation), which expectedly produced an aggressive, early-onset phenotype. The pattern of bone-spicule pigment migration in P10 is a reported feature of severe ABCA4 phenotypes associated with splice site variants2, 4 and is similar to an ethnic Indian patient homozygous for the null variant c.6088C>T described by Singh et al.34
Interestingly, several variants of different geographical and ethnic provenance were found in our cohort. The c.2894A>G, p.(N965S) variant, homozygous in the severely affected P20 of bi-parental Indian descent, is the most frequent missense and founder disease-associated ABCA4 variant10 in the Danish population (16.2% of disease-associated alleles) resulting in moderate to severe (generalized choriocapillaris dystrophy38) phenotypes. This variant is also one of the most frequent missense alleles (3.1%) in Chinese patients with cone-rod dystrophy, where an independent occurrence of this allele from that in Europe was reported by haplotype analysis.39 Patient 22, who is also of bi-parental Indian descent, harbored the c.5318C>T, p.(A1773V) missense variant which was identified in 17% of disease-associated alleles in a Mexican cohort.11 Haplotype analyses in four patients from the same geographical region harboring this allele confirmed a common origin of this variant in Mexico. The high proportion of racial admixture in the Mexican population may make it difficult to determine whether this variant resulted from multiple or a single occurrence within a specific population. The missense variant c.2966T>C, p.(V989A), which occurs at an allele frequency of 6.82% in African-American patients,12 compared to 0.20% in European patients, was found in P1, P37 and P38 who, unsurprisingly, were of both Indian and African descent. Parental testing confirmed the segregation of p.(V989A) from the Moroccan parent of P37 and P38 (siblings).
The high allele frequency of the c.5822G>A, p.(G1961E) in our cohort (23.6%) in both the compound heterozygous and homozygous state as well as in a complex allele in cis with the c.6445C>T, p.(R2149*) stop-gain variant was initially an unexpected finding. It is well established that p.(G1961E) variant is the most common disease allele (~10%) in documented cases in Europeans, where the population frequency is 0.47%. However, the frequency of the p.(G1961E) allele in the South Asian population is ~1.5% according to Exome Aggregation Consortium database (ExAC; http://exac.broadinstitute.org; accessed September 2016). Therefore, the finding of this variant in half of the patients is not entirely surprising, but rather expected based on the population frequency, which is high for a fully penetrant variant. Patients compound heterozygous for p.(G1961E) have been associated with confinement of disease changes to the central macula, preservation of generalized cone and rod function on ffERG, absence of the ‘dark choroid’ sign on fluorescein angiography and lower qAF levels relative to other ABCA4 genotypes.16, 40, 41, 42 This characterization was reflected by all p.(G1961E) patients in our cohort, which posits mild effects that are inherent to this particular variant and not ethnic background of the individual.
Retinal imaging and ffERG findings revealed both apparently mild and advanced phenotypes associated with the South Asian c.859-9T>C founder variant in patients making it difficult to elucidate its effects without functional studies. In addition, the second variant in the two advanced cases were often categorically severe including intronic variants within a canonical splice sites. Two unrelated patients harboring c.859-9T>C, P14 (compound heterozygous) and P18 (homozygous) did not exhibit end-stage phenotypes; however, analysis of qAF revealed lipofuscin levels far above 95% confidence interval for healthy eyes and above all other analyzed affected eyes when corrected for age. The fundus autofluorescence (488 nm excitation) signal originates predominantly from RPE lipofuscin, which accumulates due to ABCA4 dysfunction resulting in cell death through a number of mechanisms.43 Although ABCA4-associated disease as a group has been shown to collectively exhibit increased qAF levels, more severe phenotypes such as those associated with c.[1622T>C;3113C>T], p.[L541P/A1038V] have been shown to correspond to higher qAF relative to other affected eyes.15 The c.859-9T>C variant may thus impart a phenotype which begins with significantly more rapid lipofuscin accumulation, followed by a progression to generalized posterior pole atrophy consistent with non-detectable ffERG responses.
In summary, the clinical and genetic spectrum of an ethnically South Asian cohort of ABCA4-associated disease patients was described. Several missense variants of different geographical and ethnic origin found in the cohort. The most frequent variant was c.5882G>A, p.(G1961E), which was found in half the cohort results in comparatively mild phenotypes. The c.859-9T>C founder variant in this population is likely associated with an early, rapid accumulation of lipofuscin (significantly elevated qAF) relative to other ABCA4 genotypes followed by widespread atrophy of the macula and posterior pole within the following two decades.
References
Allikmets R, Singh N, Sun H et al: A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet 1997; 15: 236–246.
Cremers FP, van de Pol DJ, van Driel M et al: Autosomal recessive retinitis pigmentosa and cone-rod dystrophy caused by splice site mutations in the Stargardt's disease gene ABCR. Hum Mol Genet 1998; 7: 355–362.
Maugeri A, Klevering BJ, Rohrschneider K et al: Mutations in the ABCA4 (ABCR) gene are the major cause of autosomal recessive cone-rod dystrophy. Am J Hum Genet 2000; 67: 960–966.
Martinez-Mir A, Paloma E, Allikmets R et al: Retinitis pigmentosa caused by a homozygous mutation in the Stargardt disease gene ABCR. Nat Genet 1998; 18: 11–12.
Shroyer NF, Lewis RA, Yatsenko AN, Lupski JR : Null missense ABCR (ABCA4) mutations in a family with stargardt disease and retinitis pigmentosa. Invest Ophthalmol Vis Sci 2001; 42: 2757–2761.
Allikmets R : Retinal Degenerations: Biology, Diagnostics and Therapeutics. Totowa, NJ, USA: Humana Press, 2007.
Maugeri A, van Driel MA, van de Pol DJ et al: The 2588G—>C mutation in the ABCR gene is a mild frequent founder mutation in the Western European population and allows the classification of ABCR mutations in patients with Stargardt disease. Am J Hum Genet 1999; 64: 1024–1035.
Rivera A, White K, Stohr H et al: A comprehensive survey of sequence variation in the ABCA4 (ABCR) gene in Stargardt disease and age-related macular degeneration. Am J Hum Genet 2000; 67: 800–813.
Valverde D, Riveiro-Alvarez R, Bernal S et al: Microarray-based mutation analysis of the ABCA4 gene in Spanish patients with Stargardt disease: evidence of a prevalent mutated allele. Mol Vis 2006; 12: 902–908.
Rosenberg T, Klie F, Garred P, Schwartz M : N965S is a common ABCA4 variant in Stargardt-related retinopathies in the Danish population. Mol Vis 2007; 13: 1962–1969.
Chacon-Camacho OF, Granillo-Alvarez M, Ayala-Ramirez R, Zenteno JC : ABCA4 mutational spectrum in Mexican patients with Stargardt disease: Identification of 12 novel mutations and evidence of a founder effect for the common p.A1773V mutation. Exp Eye Res 2013; 109: 77–82.
Zernant J, Collison FT, Lee W et al: Genetic and clinical analysis of ABCA4-associated disease in African American patients. Hum Mutat 2014; 35: 1187–1194.
Cideciyan AV, Swider M, Aleman TS et al: ABCA4 disease progression and a proposed strategy for gene therapy. Hum Mol Genet 2009; 18: 931–941.
Zhang N, Tsybovsky Y, Kolesnikov AV et al: Protein misfolding and the pathogenesis of ABCA4-associated retinal degenerations. Hum Mol Genet 2015; 24: 3220–3237.
Burke TR, Duncker T, Woods RL et al: Quantitative fundus autofluorescence in recessive Stargardt disease. Invest Ophthalmol Vis Sci 2014; 55: 2841–2852.
Fishman GA, Stone EM, Grover S, Derlacki DJ, Haines HL, Hockey RR : Variation of clinical expression in patients with Stargardt dystrophy and sequence variations in the ABCR gene. Arch Ophthalmol 1999; 117: 504–510.
Zernant J, Xie YA, Ayuso C et al: Analysis of the ABCA4 genomic locus in Stargardt disease. Hum Mol Genet 2014; 23: 6797–6806.
McCulloch DL, Marmor MF, Brigell MG et al: ISCEV Standard for full-field clinical electroretinography (2015 update). Doc Ophthalmol 2015; 130: 1–12.
Delori F, Greenberg JP, Woods RL et al: Quantitative measurements of autofluorescence with the scanning laser ophthalmoscope. Invest Ophthalmol Vis Sci 2011; 52: 9379–9390.
Greenberg JP, Duncker T, Woods RL, Smith RT, Sparrow JR, Delori FC : Quantitative fundus autofluorescence in healthy eyes. Invest Ophthalmol Vis Sci 2013; 54: 5684–5693.
Adzhubei IA, Schmidt S, Peshkin L et al: A method and server for predicting damaging missense mutations. Nat Methods 2010; 7: 248–249.
Tavtigian SV, Deffenbaugh AM, Yin L et al: Comprehensive statistical study of 452 BRCA1 missense substitutions with classification of eight recurrent substitutions as neutral. J Med Genet 2006; 43: 295–305.
Ng PC, Henikoff S : Predicting deleterious amino acid substitutions. Genome Res 2001; 11: 863–874.
Schwarz JM, Rodelsperger C, Schuelke M, Seelow D : MutationTaster evaluates disease-causing potential of sequence alterations. Nat Methods 2010; 7: 575–576.
Zhang MQ : Statistical features of human exons and their flanking regions. Hum Mol Genet 1998; 7: 919–932.
Yeo G, Burge CB : Maximum entropy modeling of short sequence motifs with applications to RNA splicing signals. J Comput Biol 2004; 11: 377–394.
Reese MG, Eeckman FH, Kulp D, Haussler D : Improved splice site detection in Genie. J Comput Biol 1997; 4: 311–323.
Pertea M, Lin X, Salzberg SL : GeneSplicer: a new computational method for splice site prediction. Nucleic Acids Res 2001; 29: 1185–1190.
Desmet FO, Hamroun D, Lalande M, Collod-Beroud G, Claustres M, Beroud C : Human Splicing Finder: an online bioinformatics tool to predict splicing signals. Nucleic Acids Res 2009; 37: e67.
Kannabiran C, Singh H, Sahini N, Jalali S, Mohan G : Mutations in TULP1, NR2E3, and MFRP genes in Indian families with autosomal recessive retinitis pigmentosa. Mol Vis 2012; 18: 1165–1174.
Di Y, Huang L, Sundaresan P et al: Whole-exome sequencing analysis identifies mutations in the EYS gene in retinitis pigmentosa in the Indian population. Sci Rep 2016; 6: 19432.
Zhou Y, Saikia BB, Jiang Z et al: Whole-exome sequencing reveals a novel frameshift mutation in the FAM161A gene causing autosomal recessive retinitis pigmentosa in the Indian population. J Hum Genet 2015; 60: 625–630.
Srilekha S, Arokiasamy T, Srikrupa NN et al: Homozygosity mapping in leber congenital amaurosis and autosomal recessive retinitis pigmentosa in South Indian families. PLoS One 2015; 10: e0131679.
Singh HP, Jalali S, Hejtmancik JF, Kannabiran C : Homozygous null mutations in the ABCA4 gene in two families with autosomal recessive retinal dystrophy. Am J Ophthalmol 2006; 141: 906–913.
Battu R, Verma A, Hariharan R et al: Identification of novel mutations in ABCA4 gene: clinical and genetic analysis of Indian patients with Stargardt disease. Biomed Res Int 2015; 2015: 940864.
Bittles A : Consanguinity and its relevance to clinical genetics. Clin Genet 2001; 60: 89–98.
Vinchurkar MS, Sathye SM, Dikshit M : Retinitis pigmentosa genetics: a study in Indian population. Indian J Ophthalmol 1996; 44: 77–82.
Bertelsen M, Zernant J, Larsen M, Duno M, Allikmets R, Rosenberg T : Generalized choriocapillaris dystrophy, a distinct phenotype in the spectrum of ABCA4-associated retinopathies. Invest Ophthalmol Vis Sci 2014; 55: 2766–2776.
Jiang F, Pan Z, Xu K et al: Screening of ABCA4 gene in a Chinese cohort with Stargardt disease or cone-rod dystrophy with a report on 85 novel mutations. Invest Ophthalmol Vis Sci 2016; 57: 145–152.
Simonelli F, Testa F, Zernant J et al: Genotype-phenotype correlation in Italian families with Stargardt disease. Ophthalmic Res 2005; 37: 159–167.
Cella W, Greenstein VC, Zernant-Rajang J et al: G1961E mutant allele in the Stargardt disease gene ABCA4 causes bull's eye maculopathy. Exp Eye Res 2009; 89: 16–24.
Allikmets R : Simple and complex ABCR: genetic predisposition to retinal disease. Am J Hum Genet 2000; 67: 793–799.
Sparrow JR, Gregory-Roberts E, Yamamoto K et al: The bisretinoids of retinal pigment epithelium. Prog Retin Eye Res 2012; 31: 121–135.
Acknowledgements
The authors thank Professor François C. Delori of Schepens Eye Research Institute, Harvard Medical School for contributing to the development of the quantitative autofluorescence (qAF) acquisition and analysis method used in this study. This work was supported, in part, by grants from the National Eye Institute/NIH EY021163, EY019861 and EY024091, EY019007 (Core Support for Vision Research), OPOS Stiftung zugunsten Wahrnehmungsbehinderten, St Gallen, Switzerland and Alfred-Vogt-Stifung, St Gallen, Switzerland, Pangere Family Foundation, Pangere Center, Chicago Lighthouse, Robert L. Burch III Fund, Columbia University, NY, New York; New York Community Trust – Fredrick J. and Theresa Dow Wallace Fund, Columbia University, NY, New York and unrestricted funds from Research to Prevent Blindness (New York, NY) to the Department of Ophthalmology, Columbia University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Lee, W., Schuerch, K., Zernant, J. et al. Genotypic spectrum and phenotype correlations of ABCA4-associated disease in patients of south Asian descent. Eur J Hum Genet 25, 735–743 (2017). https://doi.org/10.1038/ejhg.2017.13
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2017.13
