
Abstract
In recent years, an increasing number of studies incorporated biogeography with phylogenetic analyses to reveal the origin and evolutionary history of specific floras. In this study, we constructed the mega-phylogeny of the floras of three representative regions across Yunnan, southwestern China. We analyzed the phylogenetic structure and beta diversity based on the presence/absence of species (genus or family) data to investigate the phylogenetic patterns of regional floras. We found conspicuous divergence at the genus and species level in the pattern of phylogenetic structures, which most likely related to historical biogeography. The flora of southern Yunnan was shaped by the strike-slip extrusion of Indochina and the regional climatic stability, while the flora of northwestern Yunnan was shaped by the uplift of the Himalaya-Tibetan Plateau and the oscillations of the glacial-interglacial periods. The flora of central Yunnan had nearly equal proportions of the northern and southern floras that may be derived from a common Tertiary tropical or subtropical flora. Geological events fit well with the floristic and phylogenetic patterns across Yunnan. This study highlighted the importance of linking phylogenetic analyses to biogeographic interpretations to improve our understanding of the origin, evolution and divergence of regional floras.
Similar content being viewed by others

Introduction
It is widely accepted that geological history, evolutionary events and processes, and current climatic conditions are significant factors in the formation of contemporary biogeographic patterns1,2. A time-calibrated phylogeny contains information about the phylogenetic relationships among species and the timing of evolutionary processes, such as speciation, divergence, and extinction, and can therefore be used to extract information about the evolutionary history of biological communities or floras at different temporal and spatial scales3. Recently, the term ‘phylofloristics’ was introduced to describe an analytical approach that relates the compositional similarity between entire floras to spatial and environmental distances in order to phylogenetically investigate a floristic assembly4. Phylogenetic measures provide an estimate of how much of the evolutionary history is represented in a particular region, providing a novel research and analytical approach for biogeography and making it possible to infer the relative influence of various biogeographical events in the historical period on the phylogenetic patterns of floras4,5,6. In this study, we applied this concept to investigate the contemporary pattern of floras and the associated geological events during the collision between India and Eurasia.
The collision between India and Eurasia, which began in the early Cenozoic, may be the largest active orogenic event on earth7,8. This long-lasting and ongoing tectonic process triggered associated geological events ranging from the uplift of the Himalaya-Tibetan plateau to the lateral extrusion of the continental landmass9,10. The associated orogenic and environmental effects (e.g., geomorphology and climate change), especially the later large-scale uplifts on or around the Himalaya-Tibetan Plateau11 and the strike-slip extrusion of Indochina, were reported to have driven and influenced genetic discontinuities, speciation, and the evolution of numerous plant and animal groups, as well as generating the contemporary biotic distribution pattern7,12,13. The species-based study on the biogeography of southern, central and northwestern Yunnan found that these three floras might be derived from a common flora, but the geological history of each region has influenced its flora, and they have remained divergent since the late Tertiary14,15,16. However, this biogeographical hypothesis lacks the support of phylogenetic evidence at a regional scale. Phylofloristics give us new insight to use phylogenetic information to demonstrate that the multistage uplift of the Himalayas and the strike-slip extrusion of Indochina resulted in the contemporary patterns of biogeography or initiated the biogeographical divergence of floras at a regional scale.
In this study, we investigated the floristic phylogenetic patterns of three representative regions in southern, central, and northwestern Yunnan. The present study aimed to examine the significance of key geological events, particularly the plate collision-induced uplift of the Himalaya-Tibetan Plateau and the strike-slip extrusion of Indochina, on contemporary patterns of biogeography using a comprehensive method combining floristic geography with phylogenetic information. Several factors made these regions ideal for this research. First, the Yunnan region is located at a sutural zone between Laurasia and Gondwana in geological history17,18, and an evident biogeographical divergence is believed to have occurred between these three regional floristic compositions in response to the uplift of Himalaya and extrusion of the Indochina block14,15,16. Second, floristic inventories of angiosperms in these three areas are well documented15,19,20,21. Third, the biogeography and divergence of these three floras were investigated on the basis of analyses of floristic geography14,15,16 without incorporating the phylogenetic patterns of these floras, which would allow the deduction of the influence of geological history on the floristic assembly with an evolutionary perspective.
We constructed mega-phylogenies of the regional floras and then analyzed the phylogenetic assemblage structure in each flora, as well as the phylogenetic beta diversity between floras from three taxonomical hierarchies (family, genus and species). These phylogenetic analyses were also conducted among different geographical elements (e.g., pantropic or tropical Asia genus, north temperate or temperate Asia genus) for more detailed phylogenetic investigations of these floras. Specifically, we addressed the following questions: (i) What is the pattern of the phylogenetic structure in three floras across Yunnan (i.e., southern, central and northwestern Yunnan)? (ii) What is the phylogenetic beta diversity between these floras? and (iii) Do the results integrated with phylogenetic information agree with previous biogeographic hypotheses in terms of the biogeographical divergence of the flora across Yunnan initiated by the uplift of Himalaya and extrusion of Indochina? Based on the biogeographic history described above and predicted patterns for community phylogenetic structure, we proposed three hypotheses. (i) The assemblages of flora in northwestern Yunnan are clustered, especially to temperate floristic elements, due to the quick speciation and divergence of abundant closely related taxa with the uplift of Himalaya. (ii) In contrast, the southern flora is more phylogenetically overdispersed, especially to tropical floristic elements, in response to the collision between India and Asia. This geological event resulted in southern Yunnan as a sutural zone, where distantly related taxa from Indo-Malesian flora (from Gondwana) and East Asia flora (from Laurasian) came together. (iii) The phylogenetic similarity between central and northwestern flora is nearly identical to the similarity between central and southern flora, which could derive from a common flora in geological time.
Results
Patterns of phylogenetic structure
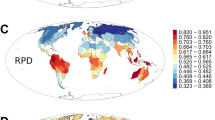
At the family level, the patterns of phylogenetic structure in these three regions were less divergent (Figures S1 and S2) but distinctly differed at the genus and species level. The genera and species of southern Yunnan exhibited a significant pattern of phylogenetic overdispersion across the entire tree (NRI), while genera and species in central and northwestern Yunnan were clustered (Fig. 1, ALL TAXA and Fig. 2). Genera that had a tropical distribution in southern Yunnan, which made up the majority of the genera (76.7% of the total), also showed significant overdispersion, particularly those with tropical Asia and tropical Asia to tropical Australia distributions. Interestingly, this pattern of overdispersion was also seen in the northwestern genera with a tropical distribution, which made up 43.5% of all genera (Fig. 1, ALL TRO-TA). However, northwestern genera with temperate distributions (45.8% of all genera) were highly clustered, especially those with old world temperate, temperate Asia and center Asia distributions (Figs 1 and 3, ALL TEM-CA). Moreover, with respect to the terminals (more recent divergences), the species and genera with a tropical distribution in southern Yunnan were clustered (Fig. 3, ALL TRO-TA and Fig. 2, NTI).

*The p-values corresponding to NRI less than 0.05. ALL TAXA, all taxa at the generic level; COSM, cosmopolitan; ALL TRO, all taxa with tropical distributions; PT, pantropic; TATAD, tropical Asia and tropical America disjointed; OWT, old world tropic; TATA, tropical Asia to tropical Australia; TATAF, tropical Asia to tropical Africa; TA, tropical Asia; ALL TEM, all taxa with temperate distributions; NT, north temperate; EANAD, east Asia and North America disjointed; OWTE, old world temperate; TEA, temperate Asia; MWACA, Mediterranean, west Asia to center Asia; CA, center Asia; EA, east Asia; and EC, endemic to China. The map was generated using ArcGIS 10.2.2. (http://www.esri.com).

*The p-values corresponding to NRI or NTI less than 0.05. NRI, the net relatedness index and NTI, the nearest taxon index. The map was generated using ArcGIS 10.2.2. (http://www.esri.com).

*The p-values corresponding to NTI less than 0.05. The map was generated using ArcGIS 10.2.2. (http://www.esri.com).
The p-values of all NRI and NTI measures and the taxonomic richness of these groups are available in the Supplementary Material (Table S1).
Phylogenetic beta diversity
The floras of the three regions at the family level were very similar (over 0.837), followed by those at the genus level (0.624–0.747), but fairly dissimilar at the species level (0.324–0.530) (Table 1). The phylogenetic similarities between the floras of central and southern Yunnan were similar to those between central and northwestern Yunnan at all taxonomic levels and geographic elements, implying that central Yunnan had nearly equal proportions of the southern and northwestern floras. Moreover, we found that the phylogenetic similarity scores of families or genera with tropical distributions were both notably higher than between temperate families or genera, which indicated a close phylogenetic relationship among tropical elements of these three floras. Additionally, higher positive values of S.E.S. Dnn in the tropical families and genera than temperate families and genera suggested that tropical and temperate taxa had smaller phylogenetic beta diversity than expected, and tropical taxa had a lower diversity than temperate taxa among these regions (Table 2).
Discussion
In this study, we extended traditional species-centric floristic analyses and phylogenetic analyses of certain taxa or local floras to incorporate phylogenetic information and focus on regional floras. We used presence/absence floristic data of regions in southern, central, and northwestern Yunnan to analyze the phylogenetic structure and beta diversity of the three regional floras to quantify how closely related pairs of taxa were within or between floras. Our results showed a conspicuous divergence at the genus and species level in the pattern of phylogenetic structures among the three representative floras. We integrated these results with the influence of geological historical events on the biogeographical divergence of regional floras to assist with interpretation.
The pattern of phylogenetic structure in the flora of southern Yunnan was overdispersed. The assemblages of species (or genera) in southern Yunnan were more phylogenetically dispersed across the whole phylogeny (NRI), and the same was found for the tropical genera, especially with tropical Asia and tropical Asia to tropical Australia distributions (Fig. 1, ALL TAXA, ALL TRO, TATA, TA and Fig. 2, NRI). This pattern aligned with the geological and biogeological events, which was the collision between India and Asia that displaced Indochina southeastward along the Ailao Shan-Red River shear zone7,22. The land and tropical climate of southern Yunnan, located at a sutural zone between Gondwana and Laurasia17,18, were shaped during this geological event. Thus, the southern biome was conspicuously linked to the dispersal of distantly related species from Palaeotropic regions and South China. Our phylogenetic results illustrated that geological changes and the associated dispersal occurred in this region. This was generally compatible with the biogeographical hypothesis, which proposed that the flora of southern Yunnan evolved in concert with the extrusion of the Indochina block and was mainly influenced by tropical Asian elements since the later Tertiary16.
However, the NTI values indicated that, with respect to the more recent divergences, the species and tropical genera in southern Yunnan were clustered (Fig. 3, ALL TRO-TA and Fig. 2, NTI). This pattern may relate to the evolution and reproduction of species under a stable tropical climate. Phylogenetic clustering at a regional scale may result from the rapid speciation rates or slow extinction rates in evolutionary time23,24. Tropical regions were expected to have relatively low rates of extinction and high rates of speciation14,25,26. The different results of NRI and NTI were due to the different identification and measurements, which emphasized the importance of the simultaneous use of two indices providing diverse information from the whole phylogeny and the more recent divergences.
The floristic composition of northwestern Yunnan showed significant phylogenetic clustering. The species (or genera) in northwestern Yunnan were clustered across the entire phylogeny (NRI) and with respect to the more recent divergences (NTI), and the same was found for the temperate genera (Fig. 1, ALL TAXA, ALL TEM; Figs 2 and 3, ALL TAXA and ALL TEM). This pattern indicated the coexistence of substantial closely related species. This clustering was consistent with the biogeographical hypothesis, as the flora of northwestern Yunnan evolved with the uplift of the Himalayas and by the gradual proliferation of mainly cosmopolitan and north temperate floristic elements16. The plate collision-induced uplifting of the Himalayas-Tibetan Plateau began in the early Cenozoic and was relatively rapid and uniform, particularly during the Quaternary period27,28. Taxa on the Himalaya-Tibetan Plateau became isolated from peripheral regions with the appearance of massive mountains and deep valleys. The continued uplift finally cut off the genetic exchange between the Himalayan range and the interior Tibetan Plateau7, promoting local speciation and hybridization of many closely related species, such as those in the genera Saussurea12, Pedicularis13, and Meconopsis29. The patterns of phylogenetic structure we observed in northwestern Yunnan reflected these geological and evolutionary processes.
We also observed that tropical genera of northwestern Yunnan, particularly those with tropical Asia and tropical Asia to tropical Australia distributions, were phylogenetically overdispersed across the whole phylogeny (NRI) (Fig. 1, ALL TRO, TATA, TA). The severe environments during the Quaternary may have filtered out many closely related tropical elements, leading to this overdispersed pattern. Another reason for such a pattern was that northwestern Yunnan may have acted as a refuge during glacial periods by providing diverse habitats in low, warm areas for plants distantly related to the local flora. This was consistent with an investigation of the Qinghai-Tibetan Plateau30, which detected phylogenetic overdispersion at the junction of the eastern edge of the plateau due to shelters during glacial periods.
Lower phylogenetic beta diversity and closer phylogenetic relationships existed among the tropical elements in these three floras rather than among the temperate elements (Tables 1 and 2). In addition, the flora of central Yunnan had nearly equal proportions of the northern and southern floras, and the phylogenetic similarities of families among the three regions were very high, which may due to the possible common origin. However, the different geological and evolutionary histories after the Tertiary were most likely to have driven the biogeographical divergence, which explained the relatively low similarity of species in focused areas. The uplift of the mountains and the oscillations in the climate created severe living conditions during the Quaternary31 that acted as a barrier to dispersal, making it difficult for species from the lowlands to migrate to the northern area. Furthermore, niche conservatism suggested that few clades crossed ecophysiological barriers to harsher environments32,33 due to the conserved ecological traits34,35,36. This suggests that various tropical elements were present in northwestern Yunnan before the Quaternary period, which might explain such a close relationship among tropical elements. These results also supported the hypothesis that the floras of Yunnan were derived from a common Tertiary flora distributed in tropical or subtropical Asia16.
Taken together, the integration of floristic geography with phylogenetic information in our analyses provided clues that revealed the influence of geological events on the biogeographical divergence of regional floras, as well as evidences for biogeographical hypotheses about Yunnan. The uplift of the Himalayas and the extrusion of Indochina fit well with the phylogenetic patterns of regional floras across Yunnan. However, our results were no doubt coarser than would be found with a more refined supertree based on DNA sequences. The method used to build the phylogeny in this study was actually not ideal, especially when some genera or species were missing in the PhytoPhylo mega-phylogeny. In addition, we should be cautious about the limitations of using phylogenetic trees to date the timing of evolutionary processes because of the crucial role of fossils, especially when using these times to interpret the biological impacts of geological events37,38. Thus, it is essential to improve the method of constructing a robust supertree, the ability of acquiring considerable DNA sequences and the study of plant fossils, which will help us reveal the origin, evolution and divergence of regional floras more objectively and truly.
Methods
Study area
Yunnan province is located in southwestern China, between 21°09′–29°15′N and 97°32′–106°12′E. The present analyses utilized the floras of three representative regions in southern, central and northwestern Yunnan (Fig. 4). Southern Yunnan (21°09′–22°36′N and 99°58′–101°50′E), which includes Xishuangbanna Dai Autonomous Prefecture, has a tropical monsoon climate and a low mountain-basin topography, with altitudes ranging from 475 m to 2,430 m across an area of 19,690 km2. Its lowlands are often dominated by tropical rainforest39. Central Yunnan (23°53′–25°11′N and 100°32′–101°58′E), which includes 7 counties and incorporates the core regions of Mt. Wuliangshan and Mt. Ailaoshan, has a subtropical climate and a middle mountain-valley topography, with altitudes ranging from 422 m to 3,157 m (mainly 1,300 m to 2,200 m) across an area of 25,424 km2. It is mostly covered by subtropical, evergreen broad-leaf forests and secondary Pinus yunnanensis forests. Northwestern Yunnan (27°10′–28°27′N and 98°53′–99°42′E), which includes 3 counties located at the center of the Hengduan Mountains, has a temperate climate and an alpine-deep valley topography, with altitudes ranging from 1,900 m to 6,740 m across an area of 23,870 km2. Its main vegetation consists of temperate sclerophyllous oak forests and cold temperate coniferous forests16.

(a) Northwestern Yunnan; (b) central Yunnan; and (c) southern Yunnan. The map was generated using ArcGIS 10.2.2. (http://www.esri.com).
Floristic data
Comprehensive synonymized inventories of the total angiosperms in southern, central and northwestern Yunnan were obtained based on checklists from several monographs15,19,20,21, the Flora of Yunnan40 and a database of angiosperms from the herbarium of the Kunming Institute of Botany, Chinese Academy of Sciences. Only native species were used in the study. All intraspecific taxa (e.g., subspecies, variety and forma) were not included in the analyses. A total of 9,370 angiosperm species from 1,860 genera and 212 families were used in this study. We checked and standardized the spelling and nomenclature of species in the floras according to The Plant List version 1.1 (TPL, available at http://www.theplantlist.org), which is an international standard database for plant nomenclature. This process aimed to maximize the match between names in the local lists and those in the backbone phylogenetic hypothesis for construction of the phylogeny. All the names in the species list that were considered synonyms were replaced with their accepted names from the TPL. We assigned each species to a family using the R package ‘plantlist’41,42 following the TPL database, in which the circumscription of angiosperm families was generally consistent with APG III43.
Furthermore, to explore the phylogenetic information concerning taxa with different patterns of geographical distribution in the study areas, patterns of angiosperms distribution were quantified at the genus and family levels based on Wu44 and Wu et al.45,46. The distribution patterns of 6 families and 5 genera were unclear, so we excluded these taxa from thephylogenetic analysis of geographical elements. Across the three regions, a total of 13 types of geographical distribution at the family level and 15 types of geographical distribution at the generic level were included.
We built a presence/absence matrix at each level (i.e., species, genus and family) based on the presence or absence of each angiosperm in each region for the following calculation of phylogenetic structure and beta diversity.
Mega-phylogeny construction
We constructed phylogenies by grafting the families, genera and species included in the study onto a backbone phylogenetic hypothesis in the R package ‘S. PhyloMaker’47. The backbone of this supertree was the PhytoPhylo mega-phylogeny47, an updated and expanded version of Zanne et al.’s species-level phylogeny33, which included 30,771 seed plants and was time-calibrated for all branches using seven gene regions available in GenBankand fossil data. PhytoPhylo included all families of extant seed plants in the world48 and had five times more genera and over 55 times more species than the newest angiosperm supertrees (i.e., R20120829)47. Genera and species that were not found in the PhytoPhylo mega-phylogeny were handled by S. PhyloMaker in one of three ways: (1) adding genera or species as polytomies within their families or genera; (2) randomly adding genera or species within their families or genera; and (3) adding genera or species to their families or genera with the same approach used in the online software Phylomatic and BLADJ. Using these three approaches, three phylogenies were generated at each level of resolution (i.e., family, genus and species). The phylogenies with the first approach are available in the Supplementary Material (Figures S3, S4 and S5). However, we mainly reported results based on the first approach. The detailed results using the other approaches are available in the Supplementary Material (Tables S2 and S3).
Phylogenetic structure
The phylogenetic structure of each flora was quantified using two indices: the net relatedness index (NRI) and the nearest taxon index (NTI). In general, NRI is considered more sensitive to tree-wide patterns of phylogenetic clustering and overdispersion, while NTI is more sensitive to patterns of overdispersion and clustering closer to the tips of the phylogenetic tree49. NRI and NTI are defined as the measure of standardized effect size of mean pairwise distance (MPD) and mean nearest taxon distance (MNTD), respectively. These indices describe the difference in phylogenetic distance (i.e., MPD or MNTD) between observed and null communities generated with some randomization methods divided by the standard deviation of phylogenetic distance in the null communities50. They are computed as follows:


where MPDobserved (or MNTDobserved) is the observed MPD (or MNTD), mean (MPDnull) [or mean (MNTDnull)] is the average MPD (or MNTD) of randomly generated assemblages, and sd (MPDnull) [or sd (MNTDnull)] is the standard deviation of MPD (or MNTD) in the null assemblages. We randomly shuffled the tip labels of the constructed phylogeny 999 times to generate null communities. A positive NRI (or NTI) value indicated a smaller phylogenetic distance among co-occurring taxa than expected (i.e., phylogenetic clustering). Conversely, a negative NRI (or NTI) value indicated a greater phylogenetic distance among co-occurring taxa than expected (i.e., phylogenetic overdispersion).
In this study, we used all the species in the three floras as a species pool and each study area as a plant community at the regional scale.
Phylogenetic beta diversity
Phylogenetic beta diversity provides insights into the phylogenetic relationships between communities. Previous studies demonstrated that Dpw (mean pairwise phylogenetic distance) was unsuitable for large-scale studies, because it did not vary significantly along environmental and distance gradients51. Thus, the present analyses used PhyloSor (phylogenetic Sørensen index) and Dnn (nearest neighbor phylogenetic distance) to evaluate the phylogenetic relationships between floras. PhyloSor is defined as the ratio of branch length between shared taxa to the total branch length of all taxa in two ecological communities52. A larger PhyloSor value indicates closer community relationships51. Dnn is defined as the mean phylogenetic distance between a species in community A and its most-related species in community B. We used a null model analysis to test whether regional floras were more or less phylogenetically similar than expected by chance53. Negative values indicated a higher than expected phylogenetic distance, and positive values indicated a lower than expected phylogenetic distance. Specifically, the PhyloSor and the standardized effect size (S.E.S.) of Dnn can be quantified as follows:


where BLij is the branch length between taxa shared by two communities, and BLi (or BLj) is the branch length between all taxa of community i (or j). Dnnobserved is the observed dissimilarity between floras. Dnnnull is the mean of Dnn of the null distributions generated by randomly shuffling the tip labels of the phylogeny 999 times. sd (Dnnnull) is the standard deviation of Dnn in the null distributions.
Analyses of phylogenetic structure and beta diversity were performed using the R packages ‘picante’49 and ‘vegan’54. The results of the phylogenetic structure were mapped with ArcGIS 10.2.2.
Additional Information
How to cite this article: Liu, S. et al. A Phylogenetic Perspective on Biogeographical Divergence of the Flora in Yunnan, Southwestern China. Sci. Rep. 7, 43032; doi: 10.1038/srep43032 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Ricklefs, R. E. Community diversity: Relative roles of local and regional processes. Science 235, 167–171 (1987).
Hawkins, B. A. & Porter, E. E. Relative influence of current and historical factors on mammal and bird diversity patterns in deglaciated north America. Glob. Ecol. Biogeogr. 12, 475–481 (2003).
Pennington, R. T., Richardson, J. E. & Lavin, M. Insights into the historical construction of species-rich biomes from dated plant phylogenies, neutral ecological theory and phylogenetic community structure. New Phytol. 172, 605–616 (2006).
Swenson, N. G. & Umaña, M. N. Phylofloristics: An example from the Lesser Antilles. J. Plant Ecol. 7, 166–175 (2014).
Kooyman, R., Rossetto, M., Cornwell, W. & Westoby, M. Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests. Global Ecol. Biogeogr. 20, 707–716 (2011).
Thornhill, A. H. et al. Continental-scale spatial phylogenetics of Australian angiosperms provides insights into ecology, evolution and conservation. J. Biogeogr. 43, 2085–2098 (2016).
Che, J. et al. Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia. Pro. Natl. Acad. Sci. USA 107, 13765–13770 (2010).
Jain, A. K. When dis India-Asia collide and make the Himalaya? Curr. Sci. 106, 254–266 (2014).
Raymo, M. E. & Ruddiman, W. F. Tectonic forcing of late Cenozoic climate. Nature 359, 117–122 (1992).
Aikman, A. B., Harrison, T. M. & Lin, D. Evidence for early (>44 Ma) Himalayan crustal thickening, Tethyan Himalaya, southeastern Tibet. Earth Planet. Sc. Lett. 274, 14–23 (2008).
An, Z., Kutzbach, J., Prell, W. & Porter, S. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since late Miocene time. Nature 411, 62–66 (2001).
Wang, Y. J., Susanna, A., Raab-Straube, E. V., Milne, R. & Liu, J. Q. Island-like radiation of Saussurea (Asteraceae: Cardueae) trigged by uplifts of the Qinghai-Tibetan Plateau. Bot. J. Linnean Soc. 97, 893–903 (2009).
Yang, F. S., Li, Y. F., Ding, X. & Wang, X. Q. Extensive population expansion of Pedicularis longiflora (Orobanchaceae) on the Qinghai-Tibetan Plateau and its correlation with the Quaternary climate change. Mol. Ecol. 17, 5135–5145 (2008).
Zhu, H. The tropical flora of southern Yunnan, China, and its biogeographic affinities. Ann. Mo. Bot. Gard. 95, 661–680 (2008).
Zhu, H. Biogeography of Shangri-la flora in southwestern China. Phytotaxa 203, 231–244 (2015).
Zhu, H. Biogeographical divergence of the flora of Yunnan, southwestern China initiated by the uplift of Himalaya and extrusion of Indochina block. Plos One 7, e45601 (2012).
Jin, X. C. Permo-Carboniferous sequences of Gondwana affinity in southwest China and their paleogeographic implications. J. Asian Earth Sci. 20, 633–646 (2002).
Metcalfe, I. Palaeozoic and Mesozoic tectonic evolution and palaeogeography of East Asian crustal fragments: The Korean peninsula in context. Gondwana Res. 9, 24–46 (2006).
Peng, H. The seed plants from Mt. Wuliang in south-central Yunnan, China (Yunnan Science and Technology Press, 1998).
Zhu, H. & Yan, L. C. List of seed plants in the Ailao Mts. of Yunnan province, China (Yunnan Science and Technology Press, 2009).
Zhu, H. & Yan, L. C. Native seed plants in Xishuangbanna of Yunnan (Science Press, 2012).
Tapponnier, P. et al. The Ailao Shan/Red River metamorphic belt: Tertiary left-lateral shear between Indochina and South China. Nature 343, 431–437 (1990).
Cardillo, M. Phylogenetic structure of mammal assemblages at large geographical scales: Linking phylogenetic community ecology with macroecology. Phil. Trans. R. Soc. B. 366, 2545–2553 (2011).
Wiens, J. J. & Graham, C. H. Niche conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 36, 519–539 (2005).
Kerkhoff, A. J., Moriarty, P. E. & Weiser, M. D. The latitudinal species richness gradient in New World woody angiosperms is consistent with the tropical conservatism hypothesis. Proc. Natl. Acad. Sci. USA 111, 8125–8130 (2014).
Matthew R. H., Thomas J. B., Christopher K. W. & Anthony R. I. Phylogenetic measures of biodiversity. Am. Nat. 169, E68–E83 (2007).
An, Y. Genozoic tectonic evolution of Asia: Apreliminary synthesis. Tectonophysics 488, 293–325 (2010).
Harrison, T. M., Copeland, P., Kidd, W. S. F. & Yin, A. Raising Tibet. Science 255, 1663–1670 (1992).
Yang, F. S., Qin, A. L., Li, Y. F. & Wang, X. Q. Great genetic differentiation among populations of Meconopsis integrifolia and its implication for plant speciation in the Qinghai-Tibetan Plateau. Plos One 7, e37196 (2012).
Yan, Y. J., Yang, X. & Tang, Z. Y. Patterns of species diversity and phylogenetic structure of vascular plants on the Qinghai-Tibetan Plateau. Ecol. Evol. 3, 4584–4595 (2013).
Liu, D. S., Zhang, X. S., Xiong, S. F. & Qin, X. G. Qinghai-Xizang Plateau glacial environment and global cooling. Quat. Sci. 19, 385–396 (1999).
Ricklefs, R. E. Evolutionary diversification and the origin of the diversity-environment relationship. Ecology 87, S3–S13 (2006).
Zanne, A. E. et al. Three keys to the radiation of angiosperms into freezing environments. Nature 506, 89–92 (2014).
Ackerly, D. D. Community assembly, niche conservatism, and adaptive evolution in changing environments. Int. J. Plant Sci. 164, S165–S184 (2003).
Donoghue, M. J. A phylogenetic perspective on the distribution of plant diversity. Proc. Natl. Acad. Sci. USA 105, 11549–11555 (2008).
Wiens, J. J. et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 13, 1310–1324 (2010).
Sauquet, H. et al. Testing the impact of calibration on molecular divergence times using a fossil-rich group: The case of Nothofagus (Fagales). Syst. Biol. 61, 289–313 (2012).
Wilf, P. & Escapa, I. H. Green Web or megabiased clock? Plant fossils from Gondwanan Patagonia speak on evolutionary radiations. New Phytol. 207, 283–290 (2015).
Zhu, H. Ecological and biogeographical studies on the tropical rain forest of south Yunnan, SW China with a special reference to its relation with rain forests of tropical Asia. J. Biogeogr. 24, 647–662 (1997).
Wu, Z. Y. Flora Yunnanica Vol. 1–21 (Science Press, 1977–2006).
R Development Core Team. R: A language and environment for statistical computing. Available at: http://www.R-project.org/. (Date of access: 08/11/2015) (2012).
Zhang, J. L. Plantlist: Looking up families of higher plants. (2015) Available at: http://R-Forge.R-project.org/projects/plantlist/. (Date of access: 20/03/2016).
The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linnean Soc. 161, 105–121 (2009).
Wu, Z. Y. The areal-types of Chinese genera of seed plants. Acta Bot. Yunnan. Supp. IV, 1–139 (1991).
Wu, Z. Y., Zhou, Z. K., Li, D. Z., Peng, H. & Sun, H. The areal-types of the world families of seed plants. Acta Bot. Yunnan. 25, 245–257 (2003).
Wu, Z. Y., Zhou, Z. K., Sun, H., Li, D. Z. & Peng, H. The areal-types of seed plants and their origin and differentiation (Yunnan Science and Technology Press, 2006).
Qian, H. & Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant Ecol. 9, 233–239 (2015).
Qian, H. & Zhang, J. Using an updated time-calibrated family-level phylogeny of seed plants to test for non-random patterns of life forms across the phylogeny. J. Syst. Evol. 52, 423–430 (2014).
Kembel, S. W. et al. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26, 1463–1464 (2010).
Webb, C. O., Ackerly, D. D., McPeek, M. A. & Donoghue, M. J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 33, 475–505 (2002)
Feng, G. et al. Comparison of phylobetadiversity indices based on community data from Gutianshan forest plot. Chin. Sci. Bull. 57, 623–630 (2011).
Bryant, J. A. et al. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 105, 11505–11511 (2008).
Webb, C. O., Ackerly, D. D. & Kembel, S. W. Phylocom: Software for the analysis of phylogenetic community structure andtrait evolution. Bioinformatics 24, 2098 (2008).
Oksanen, J. et al. Vegan: Community ecology package. Available at: http://CRAN.R-project.org/package=vegan. (Date of access: 02/11/2016) (2012).
Acknowledgements
This research was supported by the HPC platform, the public technology service center of Xishuangbanna tropical botanical garden (XTBG), CAS, China, the National Natural Science Foundation of China, No. 41471051, 41071040, 31170195, 31400362 and 31670442, the West Light Foundation of Chinese Academy of Sciences and the Chinese Academy of Sciences Youth Innovation Promotion Association (2016352). We also appreciate the reviewers for their helpful comments and suggestions to improve this manuscript.
Author information
Authors and Affiliations
Contributions
S.L., H.Z. and J.Y. conceived and designed the experiments. H.Z. provided the data. S.L. analyzed the data and wrote the first draft of the manuscript. S.L. and J.Y. contributed substantially to revisions.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Liu, S., Zhu, H. & Yang, J. A Phylogenetic Perspective on Biogeographical Divergence of the Flora in Yunnan, Southwestern China. Sci Rep 7, 43032 (2017). https://doi.org/10.1038/srep43032
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep43032
This article is cited by
-
Those Nematode-Trapping Fungi That are not Everywhere: Hints Towards Soil Microbial Biogeography
Journal of Microbiology (2023)
-
Spatial phylogenetic patterns and conservation of threatened woody species in a transition zone of southwest China
Biodiversity and Conservation (2022)
-
Genetic structure and differentiation in Dendrocalamus sinicus (Poaceae: Bambusoideae) populations provide insight into evolutionary history and speciation of woody bamboos
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
