
Abstract
HIV-specific CD8+ T cells demonstrate an exhausted phenotype associated with increased expression of inhibitory receptors, decreased functional capacity, and a skewed transcriptional profile, which are only partially restored by antiretroviral treatment (ART). Expression levels of the inhibitory receptor, T cell immunoglobulin and ITIM domain (TIGIT), the co-stimulatory receptor CD226 and their ligand PVR are altered in viral infections and cancer. However, the extent to which the TIGIT/CD226/PVR-axis is affected by HIV-infection has not been characterized. Here, we report that TIGIT expression increased over time despite early initiation of ART. HIV-specific CD8+ T cells were almost exclusively TIGIT+, had an inverse expression of the transcription factors T-bet and Eomes and co-expressed PD-1, CD160 and 2B4. HIV-specific TIGIThi cells were negatively correlated with polyfunctionality and displayed a diminished expression of CD226. Furthermore, expression of PVR was increased on CD4+ T cells, especially T follicular helper (Tfh) cells, in HIV-infected lymph nodes. These results depict a skewing of the TIGIT/CD226 axis from CD226 co-stimulation towards TIGIT-mediated inhibition of CD8+ T cells, despite early ART. These findings highlight the importance of the TIGIT/CD226/PVR axis as an immune checkpoint barrier that could hinder future “cure” strategies requiring potent HIV-specific CD8+ T cells.
Similar content being viewed by others


Introduction
During chronic HIV-1 infection CD8+ T cells gradually lose their cytotoxic function, production of antiviral cytokines and their capacity to proliferate1,2,3,4 (reviewed in ref. 5). In addition, these cells accumulate cell surface markers associated with immune dysfunction, e.g., inhibitory receptors such as PD-1, CD160, 2B4, TIM-3, and LAG-36,7,8,9,10,11,12,13,14. This pattern persists even after years of successful antiretroviral treatment (ART)9,10. The function of the CD8+ T cells is partly regulated by the T-box transcription factors T-bet and Eomesodermin (Eomes) that control effector function and memory differentiation15,16. HIV-specific cells display a specific pattern of T-box transcriptions factors (T-betdimEomeshi) that is coupled to the increased expression of inhibitory receptors9. Together, these processes result in an exhausted population of HIV-specific CD8+ T cells that accumulates over time and impairs optimal control of the infection, even after successful ART.
Recently, T cell immunoglobulin and immunoreceptor tyrosine-based inhibitory motif (ITIM) domain (TIGIT) was identified as an inhibitory receptor expressed by human T cells and natural killer (NK) cells17,18,19. TIGIT inhibits T cell and NK cell function through several mechanisms including signaling through its ITIM domain18 induction of immunoregulatory dendritic cells19 and by blocking cytotoxicity18,20. Furthermore, TIGIT is associated with a downregulation of the co-stimulatory receptor CD226 and molecules involved in TCR signaling17,21.
TIGIT expression is increased on CD8+ T cells in HIV-infection and in cancer21,22 and co-blockade of TIGIT and the PD-1/PD-L1 pathway was recently shown to in part restore the proliferative capacity and/or cytokine production of CD8+ T cells in some HIV-infected subjects23 and cancer patients21,22 in vitro, making these pathways promising candidates for restoration of CD8+ T cell responses. Differences in response to inhibitory receptor blockade treatment have also been shown in murine studies and in human T cells from HIV-infected subjects, where not all individuals responded equally to treatment24,25. Therefore, the potential usefulness of HIV-specific CD8+ T cells to eliminate target cells in immune based curative strategies requires further investigation with a particular emphasis of the T cell exhaustion process, including the role of TIGIT. Recently, CD4+ T cells expressing TIGIT, alone or together with PD-1 and/or LAG-3 were shown to be enriched for persistent HIV during ART, suggesting that inhibitory receptor blockade treatment may also have an effect on latently infected CD4+ T cells26.
TIGIT shares its ligands, poliovirus receptor (PVR, CD155) and Nectin-2 (PVRL2, CD112), with the co-stimulatory receptor CD226 (also known as DNAM-1)19. Expression of CD226 is downregulated during chronic HIV infection27. Additionally, TIGIT is able to suppress CD226 expression by directly hindering the dimerization of CD22621. Hence, TIGIT and CD226 are emerging as an inhibitory/stimulatory receptor pair of importance for CD8+ T cell function, similar to CTLA-4 and CD28 (reviewed in ref. 28). However, the link between TIGIT and CD226 in the context of HIV infection has not been evaluated, nor has the relationship of TIGIT to the inhibitory receptors CD160 and 2B4 or transcription factors involved in T cell differentiation. Additionally, expression of the TIGIT/CD226 ligand PVR is upregulated on cancer cells29,30 and HIV proteins are able to regulate PVR expression on CD4+ T cells infected with HIV in vitro31,32. However, the expression of PVR on ex vivo CD4+ T cells from HIV-infected subjects has not been described.
Here, we show that TIGIT and CD226 are differentially expressed on HIV-specific CD8+ T cells. Strikingly, elevation of TIGIT expression levels was detected in longitudinal samples from HIV infected subjects treated from early infection. Increased expression of TIGIT during HIV-1 infection was coupled to a transitional T-betdimEomeshi transcriptional phenotype and decreased functional capacity of HIV-specific CD8+ T cells. Furthermore, increased expression of the TIGIT/CD226 ligand PVR on CD4+ T cells in HIV-infected subjects was observed, especially on T follicular helper cells (Tfh), which are a major compartment of productive and latent HIV-infection33,34. Overall, these results highlight the important role of the TIGIT/CD226/PVR axis in T cell exhaustion and control of HIV-infection.
Materials and Methods
Human subjects and ethical statement
Blood samples from 30 treatment-naïve HIV-positive subjects, 20 HIV-positive subjects on long-term ART and 26 HIV-negative healthy controls were collected at the HIV clinics at Karolinska University Hospital in Huddinge and Venhälsan at Stockholm South General Hospital (Table 1). Cryopreserved peripheral blood mononuclear cells (PBMCs) from subjects with acute HIV-infection (n = 12) and elite controller subjects (n = 14) were acquired from the OPTIONS35 and SCOPE36 cohorts, respectively, at University of California San Francisco, USA (Table 1). Samples from subjects with acute HIV-infection were collected within 24–43 days (median 26.5) after the estimated infection date and all subjects initiated ART during acute HIV-infection. Of the 12 individuals with acute HIV-infection, 10 were followed longitudinally with samples collected at baseline (median 24 days), 6 months post-ART initiation (median 5.5 months) and 1.5–12 years (median 3.2 years) after the estimated infection date. Lymph nodes and matched blood samples were collected from 8 HIV-positive individuals at the Centre for Infectious Diseases Research, National Institute of Respiratory Diseases, Mexico City, Mexico. As controls, lymph nodes and blood were collected from HIV-negative subjects at the Division of Transplantation, Department of Surgery, Perelman School of Medicine, University of Pennsylvania, Philadelphia, USA. Lymph node mononuclear cells (LNMCs) were isolated through mechanical disruption of lymph nodes, either manually or according to the manufacturer’s instructions for the gentleMACS tissue dissociator (Miltenyi Biotec).
The Regional Ethical Council, Stockholm, Sweden (2014/842-32, 2009/1592-32) approved the study. The institutional review board of the University of California, San Francisco, California, USA approved the use of samples from OPTIONS and SCOPE. The Institutional Review Board at the University of Pennsylvania, Philadelphia, USA, approved collection of the lymph node and blood samples from HIV-negative subjects and the Institutional Review Board at the Centre for Infectious Diseases Research, Mexico City, Mexico approved collection of blood and lymph node samples from HIV infected subjects. Written informed consent was documented from all study subjects in accordance with the Declaration of Helsinki. All methods utilized for this study were performed in accordance with the relevant guidelines and regulations.
Antibody reagents
The following antibodies were used for extra- or intracellular staining of PBMCs; CCR7 PECy7(Clone 3D12), CD3 V500 (Clone UCHT1), CD3 APC-H7 (Clone SK7) CD4 A700 (Clone RPA-T4), CD11c BV711 (Clone B-ly6), CD14 V500 (Clone M5E2), CD16 APC-H7 (Clone 3G8), CD19 V500 (Clone HIB19), CD19 BV711 (Clone SJ25C1), CD21 PECF594 (Clone B-ly4), CD35 BV421 (Clone E11), CD38 BV711 (Clone HIT2), CD45RO BV650 (Clone UCHL1), CD56 AF488 (Clone B159), CD107a PE-CF594 (Clone H4A3), CD123 PE-Cy7 (Clone 7G3), CD160 A488 (Clone BY55), CD226 FITC or BV711 (Clone DX11), GrzB A700 (Clone GB11), HLA-DR BV605 (Clone G46-6), IFNg A700 (Clone B27), TNF PE-Cy7 or PeCF594 (Clone MAb11) were purchased from BD Bioscience. CCR7 APC-Cy7 (Clone G043H7), CD3 BV570 (Clone UCHT1), CD4 BV785 (Clone OKT4), CD8 BV605 (Clone RPA-T8), CD14 BV570 (Clone M5E2), CD19 BV785 (Clone HIB19), CD27 BV785 (Clone O323), IFN-γ APC-Cy7 (Clone 4 S.B3), PD-1 PE-Cy7 or BV421 or BV785 (Clone EH12.2H7), Perforin BV421 (Clone B-D48) PVRL2 APC (Clone TX31) and T-bet BV605 or T-bet PE (Clone 4B10) were purchased from Biolegend. CD8 PE-Cy5.5 (Clone RPA-T8), CD38 APC (Clone HIT2), Eomes eFluor660 (Clone WD1928), CXCR5 PEeFluor610 (Clone MU5UBEE), PVR PE (Clone 2H7CD155), TIGIT PE, PE-Cy7 or PerCPeFluorF710 (Clone MBSA43) were purchased from eBioscience. CD8 Qdot565 (Clone 3B5), Granzyme B PE-Cy5.5 (Clone GB11) and CD4 PE-Cy5.5 (Clone S3.5) were purchased from LifeTechnologies/Invitrogen. 2B4 PE-Cy5 (Clone C1.7) and CD45RO ECD (Clone UCHL1) were purchased from Beckman Coulter. LIVE/DEAD® Fixable Aqua Dead Cell Stain Kit from Life Technologies was used to stain for dead cells. For tetramer analysis, the HLA-A*0201 restricted tetramers HIV Gag SLYNTVATL (SL9) conjugated to PE and CMV pp65 NLVPMVATV (NV9) conjugated to PE (MBL) were used.
Cell preparation and antigens
Density gradient centrifugation with Hypaque-Ficoll (GE Healthcare), was used to isolate PBMCs from whole blood. The isolated cells were cryopreserved in FBS (LifeTechnologies) containing 10% DMSO. Cryopreserved PBMCs were thawed and washed twice in R10 media (RPMI-1640 Medium AQ Media (Sigma Aldricht) containing 10% FBS, 50 IU/mL penicillin, 50 μg/mL streptomycin and 10 mM HEPES (LifeTechnologies)). Cells were rested for 6 hours at 37 °C. Peptide pools (15-mers overlapping by 11 amino acids were used at a concentration of 1 μg/mL to detect HIV Gag p55 (JPT Technologies) and HCMV pp65 (JPT Technologies) specific CD8+ T cell responses.
Stimulation and extra- and intracellular staining of cells for flow cytometry
Stimulation and intracellular staining were performed as described previously9. Briefly, cells (1–1.5 × 106/well) were stimulated with overlapping HIV Gag-p55 and HCMV pp65 peptides or medium alone (negative controls) in the presence of anti-CD107a antibody, monensin (0.7 mg/mL, BD Bioscience) and Brefeldin A (5 μg/mL, BD Bioscience). After 10 hours cells were washed with PBS:EDTA and stained with LIVE/DEAD Aqua amine dye solution containing the extracellular antibodies. The cells were incubated for 30 minutes at room temperature; alternatively, for staining with anti-CCR7 antibodies, cells were incubated at 37 °C for 10 minutes followed by 20 minutes in room temperature. For extracellular staining only, cells were subsequently washed and fixed with PBS + 1% paraformaldehyde. For the intracellular staining, cells were fixated and permeabilized using the FoxP3 transcription factor buffer kit (eBioscience) according to the manufacturer’s protocol, followed by staining with intracellular markers and incubation for 60 minutes at room temperature. The cells were washed and resuspended in PBS + 1% paraformaldehyde.
Flow cytometry analysis
PBMCs and LMNCs were analyzed on a modified 4-laser LSR Fortessa (BD Biosciences). Antibody capture beads (BD Bioscience) were used for single-stain compensation controls. Flow cytometry data were analyzed with FlowJo 8.8.7 (Treestar) and when applicable, fluorescence minus one (FMO) gating strategies were used to set the manual gates9,37,38. A virus-specific response was considered positive when ≥0.05% of the cells produced IFN-γ after background reduction and if the response was larger than twice the negative background signal9,37,38.
Statistics
The matlab based tool, CYT, was used to generate and visualize the viSNE maps39. This analysis was performed to enable the visualization of complex expression patterns of cell surface markers on CD8+ T cell subsets. Comparisons between two groups of individuals were performed with the Mann-Whitney U test, and the Wilcoxon matched-pairs rank test was used to compare paired samples. The non-parametric Spearman rank test was used for correlation analysis. One-way ANOVA with repeated measures, followed by Kruskal-Wallis non-parametric Dunn’s multiple comparison tests were used to compare three or more groups simultaneously. The statistical tests were performed in GraphPad Prism 5.0. Permutation tests were performed in SPICE version 5.300340. In the figures statistically significant values are indicated by one to three stars; *P < 0.05, **P < 0.01 and ***P < 0.001).
Results
TIGIT is expressed on effector/memory CD8+ T cell populations in healthy and HIV-infected individuals
TIGIT is expressed on human T cells and NK cells18,19. On CD8+ T cells, it is mainly expressed on intermediate/transitional and effector cells23. Here, we first set out to analyze the expression pattern of TIGIT in relation to its complementary co-stimulatory receptor CD226 and the inhibitory receptor PD-1 on different CD8+ T cell populations (gating strategy shown in Fig. 1a). The expression patterns of TIGIT, CD226 and PD-1 on naïve and memory CD8+ T cell populations (defined by the markers CD45RO, CCR7 and CD27) were visualized using viSNE maps in healthy (Supplementary Fig. S1) and HIV infected individuals (Fig. 1b–d). In all groups, TIGIT was expressed on all memory CD8+ T cell subsets but not on naïve CD8+ T cells, whereas PD-1 was more narrowly expressed on memory subpopulations within the central, effector, and transitional memory cells (Fig. 1d, and Supplementary Fig. S1). In contrast, CD226 had a broad range of expression, being highly expressed on all CD8+ T cell subsets including the naïve cells (Fig. 1c). The vast majority of cells expressing PD-1 were TIGIT+, however, not all TIGIT+ cells were PD-1+ demonstrating that TIGIT is more broadly expressed on CD8+ memory T cells than PD-1.

(a) Gating strategy for TIGIT expression on CD8+ T cells in a representative subject. (b) viSNE map of naïve/memory phenotype distribution (based on CD45RO, CCR7 and CD27) in HIV+ subjects. Colors depict memory phenotypes. (c) Distribution of cells based on treatment status, HIV+ART− (green, n = 6) and HIV+ART+ (orange, n = 6) based on memory populations shown in (b). (d) Distribution of TIGIT, PD-1 and CD226 based on memory populations shown in (b). Colors depict intensity of protein expression.
Elevated levels of TIGIT+ CD8+ T cells despite early initiation of ART
TIGIT expression was recently found to be increased on CD8+ T cells during chronic HIV-infection23. Here, we extended these studies and evaluated the longitudinal impact of ART on the expression levels of TIGIT. First, TIGIT expression was measured on total CD8+ T cells by multi-color flow cytometry in four groups of HIV-1-infected individuals; chronically infected treatment-naïve subjects, chronically infected successfully long-term treated subjects, elite controllers, and subjects with acute HIV-infection (Table 1, Fig. 2a). As compared to HIV-uninfected adults, the frequency of TIGIT+ CD8+ T cells was increased in chronic treatment-naïve and long-term treated subjects but not in elite controllers nor in individuals with acute infection. Similar patterns were observed when excluding the naïve CD8+ T cells from the analysis and when measuring the expression of PD-1, CD160 and 2B4 on total CD8+ T cells (Supplementary Fig. S2). However, treatment-naïve subjects had significantly higher TIGIT mean fluorescence intensity (MFI) compared to the long-term treated subjects, the other groups of HIV-positive subjects, and healthy controls (Fig. 2a and Supplementary Fig. S2).

Expression of TIGIT on CD8+ T cells in HIV-positive subjects before and after ART (a) Frequency and MFI of TIGIT on CD8+ T cells in HIV+ treatment naïve (HIV+ART−, n = 30), long term treated (HIV+ART+, n = 20), elite controller (EC, n = 14), acutely infected (AI, n = 12) subjects and healthy controls (HC, n = 26). (b) Frequency and MFI of TIGIT on total CD8+ T cells over time in subjects followed from acute HIV infection (n = 12). Blue arrows indicate ART. (c) Frequency of TIGIT+ cells and TIGIT MFI on CD8+ T cells in subjects followed from acute HIV infection to chronic infection (n = 12). Circles represent acute HIV infection, squares represent the timepoint 6 months after ART initiation and triangles represent the timepoint 1–12 years after ART initiation. One-way ANOVA followed by Kruskall-Wallis test and Dunn’s multiple comparisons test were used to compare between ≥3 groups. The Wilcoxon matched-pairs signed rank test was used to compare paired samples. The Spearman non-parametric test was used for correlation analysis. *P < 0.05, **P < 0.01 and ***P < 0.001.
We next investigated if early initiation of ART during acute HIV-infection would affect TIGIT expression levels. Longitudinal samples were collected (within median 26.5 days after the estimated infection date) from 10 study subjects initiated on ART during acute infection from two or three time-points up to 12 years (Table 1). Despite early initiation of ART, both the frequency of TIGIT+ cells and TIGIT MFI increased in the subsequent samples and was significantly higher in the samples obtained after one year on treatment (Fig. 2b and c). After one year, the frequency of TIGIT+ CD8+ T cells and TIGIT MFI on CD8+ T cells did not differ from long-term treated subjects in whom ART was initiated during chronic HIV-infection (Supplementary Fig. S2).
TIGIT correlates with immunological markers of T cell pathology
In addition to TIGIT, a number of other inhibitory receptors linked to immune dysfunction, such as PD-1, CD160 and 2B46,7,9,10,12,13, are expressed on HIV-specific CD8+ T cells during the course of infection. However, since the link between TIGIT, PD-1, CD160 and 2B4 has not been investigated, we measured expression patterns of these markers on CD8+ T cells (Fig. 3a). Expression of TIGIT in combinations with PD-1, CD160 and 2B4 was significantly increased in the HIV-positive subject groups compared to healthy controls (Fig. 3b). Significant differences were seen between all groups except the chronically infected successfully long-term treated subjects and the elite controllers (Fig. 3b). The frequency of TIGIT+ cells was positively correlated with single- and triple expression of PD-1, CD160 and 2B4 (Supplementary Fig. S3) on CD8+ T cells.

Expression of TIGIT, PD-1, CD160, 2B4 and T-box transcription factors on CD8+ T cells in HIV-positive subjects (a) Gating procedure for co-expression of TIGIT versus PD-1, CD160 and 2B4 on CD8+ T cells in a representative HIV+ subject (HIV+) and a healthy control (HC) subject. (b) SPICE analysis of combinations of TIGIT, PD-1, CD160 and 2B4 on total CD8+ T cells in HIV-positive treatment naïve (HIV+ART−, red, n = 30), long-term treated (HIV+ART−, blue, n = 20), elite controller (EC, orange, n = 14) subjects and healthy controls (HC, black, n = 26). (c) Expression of T-betdimEomeshi (red) and T-bethiEomesdim (blue) shown for total CD8+ T cells. Gating of T-bet and Eomes populations was performed according to previously used gating strategies9. (d) TIGIT expression on T-betdimEomeshi and T-bethiEomesdim populations on total CD8+ T cells in HIV+ART−, HIV+ART+ and EC subjects (n = 64). Permutation test was performed between the pie charts. Bar charts show median and IQR. The Wilcoxon matched-pairs signed rank test was used to compare paired samples. ***P < 0.001.
T cell memory formation and functionality are controlled at the transcriptional level, and the T-box transcription factors T-bet and Eomes play an important role in these processes15,16. We, and others have recently shown that dysfunctional CD8+ T cells expressing high levels of PD-1, CD160 and 2B49,16 have a specific T-betdimEomeshi expression profile, while T-bethiEomesdim cells define a less exhausted phenotype. Accordingly, we found that TIGIT MFI and the frequency of TIGIT+ cells were significantly higher in the T-betdimEomeshi population compared to the T-bethiEomesdim cells (Fig. 3c,d and Supplementary Fig. S4). Furthermore, the frequency of TIGIT+ CD8+ T cells correlated positively with the frequency of CD8+ T-betdimEomeshi cells in chronically infected HIV-positive subjects (Supplementary Fig. S4). These data suggest that TIGIT is expressed on a highly exhausted population of CD8+ T cells.
Subsequently we investigated the relationship between TIGIT and clinical laboratory markers (viral load, CD4 count, CD4% and the CD4/CD8 ratio) and level of immune activation (CD38+HLA-DR+). Neither the frequency of TIGIT+ cells or the TIGIT MFI correlated with viral load in the chronically infected treatment-naïve subjects (excluding the elite controllers) (Supplementary Fig. S3). However, both the frequency of TIGIT+ cells and TIGIT MFI on CD8+ T cells were inversely correlated with CD4 count, the frequency of CD4+ T cells and the CD4/CD8 ratio (Supplementary Fig. S3). Furthermore, the frequency of CD38+HLA-DR+ cells was positively correlated to both the MFI and frequency of TIGIT+ cells (Supplementary Fig. S3). These results are in agreement with our previous findings in which the frequencies of CD4+ T cells and the CD4/CD8 ratio correlated strongly with PD-1 expression as well as immune activation37,41. These findings demonstrate that antiretroviral treatment initiated in chronic infection decreases TIGIT expression on TIGIT+ cells even though the frequency of TIGIT+ cells does not change significantly, which is in agreement with previous data for other inhibitory receptors6,7,8,9,10.
Differential expression of TIGIT and CD226 on HIV-specific CD8+ T cells
We next compared TIGIT expression on HIV-specific CD8+ T cells to CMV-specific cells that retain their effector capacity during chronic infection42,43. The vast majority of all HIV-specific CD8+ T cells expressed TIGIT, whereas CMV-specific cells displayed a significantly lower frequency of TIGIT+ cells and lower TIGIT MFI (Fig. 4a). The difference was most pronounced in the treatment naive and elite controller groups (Fig. 4a). The higher TIGIT MFI on HIV-specific cells was confirmed by tetramer staining identifying HLA-A2-restricted HIV-SL9-specific and CMV-NV9-specific CD8+ T cells in HIV+ subjects (n = 7) (Supplementary Fig. S4). Next, the co-expression pattern of TIGIT, PD-1, CD160 and 2B4 was measured on virus-specific CD8+ T cells. SPICE analysis revealed that treatment naïve subjects had a higher frequency of HIV-specific cells expressing all four inhibitory receptors compared to elite controllers (Fig. 4b). Three out of the four most common combinations contained TIGIT together with PD-1 (TIGIT+PD-1+CD160+2B4+, TIGIT+PD-1+CD160−2B4+, TIGIT+PD-1+CD160−2B4−) (Fig. 4b). In contrast, CMV-specific cells from the same subjects were more likely not to express inhibitory receptors on their surface and very few expressed all four inhibitory receptors (Fig. 4c).

(a) Frequency and mean fluorescence intensity of TIGIT on HIV-specific (filled circles) and CMV-specific (open circles) CD8+ T cells in HIV-positive treatment-naive (HIV+ART−, red, n = 27), long-term treated (HIV+ART+, blue, n = 8) and elite controller (EC, orange, n = 11) subjects. (b,c) SPICE analysis of inhibitory receptor co-expression on HIV-specific CD8+ T cells (b) and CMV-specific CD8+ T cells (c) in HIV+ART− (n = 27), HIV+ART+ (n = 8) and EC (n = 11) subjects. (d) Frequency of CD226+ cells of HIV-specific (filled circles) and CMV-specific (open circles) CD8+ T cells in HIV+ART− (n = 15), HIV+ART+ (n = 10) and EC (n = 10) subjects. (e) Gating strategy for TIGIT hi/dim/neg and CD226 hi/dim/neg on total CD8+ T cells and an overlay of HIV-specific (red) and CMV-specific (blue) cells from a representative treatment naïve HIV infected subject. (f) Frequency of TIGIThi and TIGITdim HIV-specific (closed circles) and CMV-specific (open circles) CD8+ T cells that are CD226hi, CD226dim and CD226neg in chronically HIV infected subjects (n = 46). The Wilcoxon matched-pairs signed rank test was used to compare paired samples. The Mann-Whitney test was used to compare two groups. Permutation test was performed between the pie charts. Median and IQR are shown for all bars. *P < 0.05, **P < 0.01 and ***P < 0.001.
Expression of TIGIT’s complementary stimulatory receptor CD226 is decreased on HIV- and melanoma-specific CD8+ T cells and cells with a high expression of PD-122,27. However, the co-expression pattern of TIGIT and CD226 has not been investigated in HIV-infection. Here, we show that the frequency of CD226+ cells and CD226 MFI was lower on HIV-specific cells as compared to CMV-specific cells, especially in the treatment naïve and elite controller subjects (Fig. 4d and Supplementary Fig. S4). In the long-term treated subjects the frequency of CD226+ cells was partially restored, but remained lower than on CMV-specific cells from the same subjects (Fig. 4d). Next, the CD8+ T cells were divided into subpopulations (hi/dim/neg) based on intensity of TIGIT and CD226 expression (Fig. 4e). HIV-specific cells were predominantly TIGIThiCD226neg/dim, whereas CMV-specific cells were TIGITdimCD226dim (Fig. 4f).
We next determined whether the TIGIT+ HIV-specific cells were activated (Supplementary Fig. S4). A positive correlation was found between the number of TIGIT+ cells and the number of highly activated (CD38+HLA-DR+) cells (Supplementary Fig. S4). These results show that the balance between CD226 and TIGIT expression is disrupted on HIV-specific cells and that HIV-specific cells expressing TIGIT are linked to a phenotype associated with severe dysfunction9.
TIGIThi HIV-specific CD8+ T cells have a decreased functional capacity
As TIGIT MFI on HIV-specific cells was markedly higher in chronic untreated HIV-1 infection and discriminated HIV-specific from CMV-specific cells, we next investigated if the intensity of TIGIT expression on CD8+ T cells (Fig. 5a) was associated with their functional capacity. TIGIThi cells were more frequent in treatment-naïve subjects compared to long-term treated subjects and elite controllers (Fig. 5b and Supplementary Fig. S5). In contrast, only a small fraction of the CMV-specific cells were TIGIThi (Supplementary Fig. S5). Next, the functional capacity (expression of IFN-γ, TNF, granzyme B and CD107a) of TIGIThi HIV-specific CD8+ T cells was measured. The frequency of TIGIThi HIV-specific cells (measured by expression of IFN-γ) was inversely correlated with the frequency of polyfunctional CD107a+GrzB+IFN-γ+TNF+ cells as well as single expression of IFN-γ (Fig. 5c). A high frequency of polyfunctional CD8+ T cells was only detected in elite controllers and long-term treated subjects. On the contrary, single expression of CD107a correlated positively with the frequency of TIGIThi cells (Fig. 5c) and was primarily detected in the treatment-naïve subjects. In CMV-specific CD8+ T cells, the frequency of TIGIThi cells correlated inversely with the frequency of polyfunctional cells; however, no correlation was found for TIGIThi cells versus the frequency of cells expressing a single function (IFN-γ, TNF or CD107a) (Supplementary Fig. S5). This suggests that TIGIThi cells represent a dysfunctional population of HIV-specific CD8+ T cells that is most common in treatment naïve HIV-positive subjects. Furthermore we found that TIGIThi cells had a higher frequency of T-betdimEomeshi cells as compared to TIGITdim cells (Fig. 5d). A similar observation has been made for PD-1hi cells that are more likely to be Eomeshi whereas PD-1dim cells are T-bethi 16, suggesting that TIGIThi cells (similarly to PD-1hi cells) represent a more transcriptionally exhausted phenotype.

Functional capacity of HIV-specific CD8+TIGIThi cells (a) Gating procedure for TIGIThi, TIGITdim and TIGITneg cells on CD8+ T cells in a representative treatment naïve HIV infected subject. (b) Frequency of TIGIThi cells of HIV-specific CD8+ T cells in HIV-positive treatment-naïve (HIV+ART−, n = 28), HIV+ long-term treated (HIV+ART+, n = 18) and elite controller (EC, n = 11) subjects. (c) Correlation between the frequency of TIGIThi cells and the frequency of polyfunctional CD8+ T cells or single expression of IFN-γ and CD107a on HIV-specific CD8+ T cells in HIV+ART− (red, n = 28), HIV+ART+ (blue, n = 18) and EC (orange, n = 11) subjects. (d) Frequency of T-betdimEomeshi cells in the HIV-specific TIGIThi and TIGITdim population in HIV+ART− (n = 28), HIV+ART+ (n = 18) and EC (n = 11) subjects. One-way ANOVA followed by Kruskall-Wallis test and Dunn’s multiple comparisons test was used to compare between ≥3 groups. The Spearman non-parametric test was used for correlation analysis. The Wilcoxon matched-pairs signed rank test was used to compare paired samples. *P < 0.05 and ***P < 0.001.
Expression of the TIGIT ligand PVR is increased on CD4+ T cells from HIV-infected lymph nodes
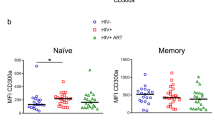
To exert its inhibitory function, TIGIT must bind to its ligand PVR on antigen presenting cells. However, the expression of PVR in HIV-positive subjects has not been investigated ex vivo. For that purpose, we measured PVR expression on CD4+ T cells from peripheral blood and lymph nodes in treatment naïve HIV-positive subjects (n = 10) and healthy controls (n = 8) (Fig. 6a and b). Expression of PVR on CD4+ T cells was significantly higher in both lymph node and peripheral blood in HIV-positive individuals, compared to healthy controls (Fig. 6b). Further, since numbers of T follicular helper cells (Tfh) cells are expanded and represent a major reservoir of HIV33,34, we next determined PVR expression on these cells in HIV-positive subjects. Tfh cells were defined by an expression profile of CD4+PD-1hiCXCR5+ (Fig. 6c) and were found mainly in the lymph nodes. In HIV-infected subjects, Tfh cells had a higher frequency of PVR+ cells and a higher PVR MFI compared to memory CD4+ T cells in the same subjects (Fig. 6d). The increased expression of PVR could be a result of an increased activation of CD4+ T cells. Indeed, the frequency of CD4+ T cell activation and PVR expression correlated positively on Tfh and CD4+ memory T cells (Fig. 6e) and Tfh cells displayed a more activated phenotype as compared to CD4+ memory cells (Fig. 6f). These data demonstrate that PVR is increased on CD4+ T cells and is highly elevated on Tfh cells in HIV-infected subjects. Although PVRL2 (CD112) was also measured in these samples, no significant differences were found (data not shown).

(a) Gating strategy for PVR+ cells of total CD4+ T cells from a representative HIV+ and healthy control (HC) subject. (b) Frequency of PVR+ cells of CD4+ T cells in HIV+ (n = 10) and HC (n = 7) subjects in lymph node and peripheral blood. (c) Gating strategy for PVR expression on T follicular helper (Tfh) cells and histogram showing PVR expression of Tfh cells (red) and memory CD4+ T cells (blue). (d) Frequency of PVR+ cells of Tfh cells versus memory CD4+ T cells in lymph nodes of HIV infected subjects (n = 10). (e) Correlation analysis of the frequency of CD38+HLA-DR+ cells versus frequency of PVR expression on Tfh cells (red) and CD4+ memory T cells (blue). (f) Comparison between the frequency of CD38+HLA-DR+ cells of Tfh versus CD4+ memory cells in the lymph nodes from HIV infected subjects (n = 10). The Mann-Whitney test was used for comparisons between two groups. The Wilcoxon matched-pairs signed rank test was used to compare paired samples. The Spearman non-parametric test was used for correlation analysis. **P < 0.01 and ***P < 0.001.
Discussion
HIV-1 infection is characterized by severe perturbation of the cellular immune system, a process referred to as immune exhaustion. This process has previously been linked to the co-expression of several inhibitory receptors, including PD-1, CD160, 2B4, TIM-3 and LAG-3 on CD8+ T cells5,9,10, poor functionality44,45 and an altered transcriptional profile9. A recently described inhibitory receptor, TIGIT, known to inhibit the function of T cells and NK cells17,18,19, is expressed on the majority of HIV-specific CD8+ T cells23 and tumor-infiltrating lymphocytes21,22. Here, in a set of longitudinal samples, we report an increased TIGIT expression over time despite early intervention with ART during acute HIV infection. The expression of TIGIT was closely linked to expression of the inhibitory receptors PD-1, CD160 and 2B4 and to a T-betdimEomeshi transcriptional profile. Additionally, TIGIThi HIV-specific CD8+ T cells were predominantly CD226dim/neg and associated with a poor functional capacity. Furthermore, we demonstrate that expression of the TIGIT/CD226 ligand, PVR was increased on Tfh cells in HIV-positive subjects, which represent a major reservoir for HIV-1, adding to the number of obstacles one must overcome to clear HIV-1 infected cells by immune-based curative strategies.
First, in agreement with recent report by Chew et al.23, we show an increased expression of TIGIT on bulk and memory CD8+ T cells in HIV-positive subjects. We further extended these recent data by demonstrating in longitudinal samples from subjects followed from acute to chronic HIV-infection, that ART did not prevent the increase of TIGIT expression, nor did it significantly decrease the frequency of TIGIT+ cells in subjects treated from chronic infection. A similar pattern was previously shown for PD-1 expression, where a decrease in viral load after treatment was more strongly related to the decreased MFI of PD-1 than the frequency of PD-1+ cells6,8,9,10. Furthermore, this process might be influenced by the establishment of the viral reservoir within lymphoid tissues, i.e. infection of the Tfh cells34 and the suggested ongoing viral replication despite successful ART46. However, it remains to be identified whether the elevated TIGIT expression levels in individuals treated from acute infection might be associated with persistent RNA replication within the viral reservoirs.
We next demonstrated that the inhibitory receptors PD-1, CD160 and 2B4, expressed on exhausted CD8+ T cells in HIV-infection9,10, were linked to TIGIT expression on bulk and HIV-specific CD8+ T cells. Our data further show that a high level of viral replication is linked to the markedly higher frequency of CD8+ T cells expressing TIGIT, PD-1, CD160 and 2B4, seen in untreated subjects as compared to subjects with undetectable viral load (long-term treated subjects and elite controllers). Importantly, long-term ART did not restore the levels of inhibitory receptor expression to levels seen in healthy subjects, as previously shown for PD-1, CD160 and 2B49, suggesting that a decrease in viral load alone does not fully restore CD8+ T cells in HIV-infection.
Previous studies on PD-1+ cells have shown different functional properties and differential expression of T-bet and Eomes depending on the intensity of PD-1 expression16. Additionally, TIGIT can repress T-bet expression in human CD4+ T cells47 and our group recently linked cells with a T-betdimEomeshi transcriptional profile to a population of highly exhausted CD8+ T cells in chronic HIV infection9. The results show that exhausted T cells during HIV-infection lose their expression of T-bet, which is linked to the functional impairment of the CD8+ T cells and thus an inability to clear infected cells. Here, we accordingly link the T-betdimEomeshi cells phenotype to an increased expression of TIGIT in HIV-infection. However it remains to be determined whether T-bet and Eomes are able to directly regulate TIGIT expression on CD8+ T cells.
TIGIT expression is increased on HIV-specific CD8+ T cells and CD8+ tumor-infiltrating lymphocytes21,22,23, and a decreased CD226 expression on PD-1+CD8+ T cells is associated with an increase in viral load in chronically HIV infected subjects27. Activation of CD8+ T cells by non-professional antigen-presenting cells, such as CD4+ T cells, requires expression of CD22648, and is likely of importance for HIV-specific CD8+ T cell recognition of infected CD4+ T cells. We here demonstrate that HIV-specific TIGIThi CD8+ T cells were mainly CD226dim/neg, likely due to the ability of TIGIT to disrupt CD226 dimerization21. These data suggest that the role of CD226 as a co-stimulatory molecule is limited in HIV-infection. Interestingly, co-blockade of TIGIT and PD-1 on CD8+ T cells was recently shown to be less effective on tumor infiltrating lymphocytes as compared to circulating melanoma-specific CD8+ T cells in patients with metastatic melanoma. This might be related to the expression of CD226, which is expressed by most tumor-specific cells in the periphery but downregulated on tumor-infiltrating lymphocytes22. Therefore, the effect of TIGIT blockade is likely to be related to expression of CD226. The modest effect on HIV-specific CD8+ T cell function after co-blockade of TIGIT and PD- seen by Chew et al.23 might therefore be explained by the loss of CD226, however this remains to be determined. The expression pattern of TIGIThiCD226neg/dim on HIV-specific cells shown here likely represents a way for the immune system to balance the antiviral response in order to limit immune damage by increasing inhibition and decreasing activation. However, in terms of restoring CD8+ T cells for immune based cure strategies49, this process becomes troublesome, as the CD8+ T cell responses are dampened.
Additionally the frequency of HIV-specific TIGIThi CD8+ T cells was negatively associated with polyfunctionality measured as co-expression of IFN-γ, TNF, granzyme B and CD107a, as well as single expression of IFN-γ. Interestingly, the frequency of HIV-specific TIGIThi CD8+ T cells was positively correlated with the number of CD107a single positive cells. In our previous studies, single expression of CD107a was linked to a highly exhausted phenotype9. Thus, the TIGIThi CD8+ T cells have the capacity to degranulate, but most degranulating cells do not express granzyme B, suggesting that the cells may not be able to eliminate infected cells. However this remains to be determined.
TIGIT induced inhibition of degranulation requires TIGIT binding to its ligand PVR on target cells20 (e.g. HIV-infected CD4+ T cells). We found an increased expression of PVR on CD4+ T cells from lymph nodes and peripheral blood of HIV-infected individuals. Interestingly, the highest levels of PVR expression on CD4+ T cells was found on Tfh cells, providing evidence that PVR expression could be increased on HIV infected CD4+ T cells. Data from HIV- and SIV-infection show that HIV- and SIV-specific cells do not generally enter the B cell follicles where Tfh cells reside50,51. Our data suggest, that even if CD8+ T cells were to reach the B cell follicles, their cytolytic function would likely be limited due to the high expression of TIGIT and PVR and the loss of CD226 expression. This creates a barrier for HIV-specific CD8+ T cells to overcome in order to eliminate the HIV reservoir.
In summary, our findings show an increased expression of TIGIT on bulk and HIV-specific CD8+ T cells during HIV-infection coupled to increased expression of the inhibitory markers PD-1, CD160 and 2B4, a T-betdimEomeshi transcriptional profile and loss of functional capacity. Importantly, early initiation of ART did not prevent a steady longitudinal increased expression of TIGIT. In addition, expression of the TIGIT’s complementary co-stimulatory molecule CD226 was decreased on HIV-specific CD8+ T cells and expression of their shared ligand PVR was increased on CD4+ T cells, specifically on Tfh cells. Our results demonstrate a perturbation of the TIGIT/CD226/PVR axis linked to multiparametric T cell pathology in HIV infection despite ART. This outcome suggests that future immune-based curative strategies will need to reverse the imbalance of the TIGIT/CD226/PVR axis caused by HIV.
Additional Information
How to cite this article: Tauriainen, J. et al. Perturbed CD8+ T cell TIGIT/CD226/PVR axis despite early initiation of antiretroviral treatment in HIV infected individuals. Sci. Rep. 7, 40354; doi: 10.1038/srep40354 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Zajac, A. J. et al. Viral immune evasion due to persistence of activated T cells without effector function. J Exp Med 188, 2205–2213 (1998).
Goepfert, P. A. et al. A significant number of human immunodeficiency virus epitope-specific cytotoxic T lymphocytes detected by tetramer binding do not produce gamma interferon. J Virol 74, 10249–10255 (2000).
Kostense, S. et al. High viral burden in the presence of major HIV-specific CD8(+) T cell expansions: evidence for impaired CTL effector function. Eur J Immunol 31, 677–686, doi: 10.1002/1521-4141(200103)31:3 <677::AID-IMMU677>3.0.CO;2-M (2001).
Shankar, P. et al. Impaired function of circulating HIV-specific CD8(+) T cells in chronic human immunodeficiency virus infection. Blood 96, 3094–3101 (2000).
Wherry, E. J. T cell exhaustion. Nat Immunol 12, 492–499 (2011).
Day, C. L. et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443, 350–354, doi: 10.1038/nature05115 (2006).
Petrovas, C. et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J Exp Med 203, 2281–2292, doi: 10.1084/jem.20061496 (2006).
Trautmann, L. et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat Med 12, 1198–1202, doi: 10.1038/nm1482 (2006).
Buggert, M. et al. T-bet and Eomes are differentially linked to the exhausted phenotype of CD8+ T cells in HIV infection. PLoS Pathog 10, e1004251, doi: 10.1371/journal.ppat.1004251 (2014).
Yamamoto, T. et al. Surface expression patterns of negative regulatory molecules identify determinants of virus-specific CD8+ T-cell exhaustion in HIV infection. Blood 117, 4805–4815, doi: 10.1182/blood-2010-11-317297 (2011).
Peretz, Y. et al. CD160 and PD-1 co-expression on HIV-specific CD8 T cells defines a subset with advanced dysfunction. PLoS Pathog 8, e1002840, doi: 10.1371/journal.ppat.1002840 (2012).
Vigano, S. et al. CD160-associated CD8 T-cell functional impairment is independent of PD-1 expression. PLoS Pathog 10, e1004380, doi: 10.1371/journal.ppat.1004380 (2014).
Pombo, C., Wherry, E. J., Gostick, E., Price, D. A. & Betts, M. R. Elevated Expression of CD160 and 2B4 Defines a Cytolytic HIV-Specific CD8+ T-Cell Population in Elite Controllers. J Infect Dis 212, 1376–1386, doi: 10.1093/infdis/jiv226 (2015).
Jones, R. B. et al. Tim-3 expression defines a novel population of dysfunctional T cells with highly elevated frequencies in progressive HIV-1 infection. J Exp Med 205, 2763–2779, doi: 10.1084/jem.20081398 (2008).
Kao, C. et al. Transcription factor T-bet represses expression of the inhibitory receptor PD-1 and sustains virus-specific CD8+ T cell responses during chronic infection. Nat Immunol 12, 663–671, doi: 10.1038/ni.2046 (2011).
Paley, M. A. et al. Progenitor and terminal subsets of CD8+ T cells cooperate to contain chronic viral infection. Science 338, 1220–1225, doi: 10.1126/science.1229620 (2012).
Joller, N. et al. Cutting edge: TIGIT has T cell-intrinsic inhibitory functions. J Immunol 186, 1338–1342, doi: 10.4049/jimmunol.1003081 (2011).
Stanietsky, N. et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc Natl Acad Sci USA 106, 17858–17863, doi: 10.1073/pnas.0903474106 (2009).
Yu, X. et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat Immunol 10, 48–57, doi: 10.1038/ni.1674 (2009).
Liu, S. et al. Recruitment of Grb2 and SHIP1 by the ITT-like motif of TIGIT suppresses granule polarization and cytotoxicity of NK cells. Cell Death Differ 20, 456–464, doi: 10.1038/cdd.2012.141 (2013).
Johnston, R. J. et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function. Cancer Cell 26, 923–937, doi: 10.1016/j.ccell.2014.10.018 (2014).
Chauvin, J. M. et al. TIGIT and PD-1 impair tumor antigen-specific CD8(+) T cells in melanoma patients. J Clin Invest 125, 2046–2058, doi: 10.1172/JCI80445 (2015).
Chew, G. M. et al. TIGIT Marks Exhausted T Cells, Correlates with Disease Progression, and Serves as a Target for Immune Restoration in HIV and SIV Infection. PLoS Pathog 12, e1005349, doi: 10.1371/journal.ppat.1005349 (2016).
Blackburn, S. D., Shin, H., Freeman, G. J. & Wherry, E. J. Selective expansion of a subset of exhausted CD8 T cells by alphaPD-L1 blockade. Proc Natl Acad Sci USA 105, 15016–15021, doi: 10.1073/pnas.0801497105 (2008).
Peligero, C. et al. PD-L1 Blockade Differentially Impacts Regulatory T Cells from HIV-Infected Individuals Depending on Plasma Viremia. PLoS Pathog 11, e1005270, doi: 10.1371/journal.ppat.1005270 (2015).
Fromentin, R. et al. CD4+ T Cells Expressing PD-1, TIGIT and LAG-3 Contribute to HIV Persistence during ART. PLoS Pathog 12, e1005761, doi: 10.1371/journal.ppat.1005761 (2016).
Cella, M. et al. Loss of DNAM-1 contributes to CD8+ T-cell exhaustion in chronic HIV-1 infection. Eur J Immunol 40, 949–954, doi: 10.1002/eji.200940234 (2010).
Topalian, S. L., Drake, C. G. & Pardoll, D. M. Immune checkpoint blockade: a common denominator approach to cancer therapy. Cancer Cell 27, 450–461, doi: 10.1016/j.ccell.2015.03.001 (2015).
Masson, D. et al. Overexpression of the CD155 gene in human colorectal carcinoma. Gut 49, 236–240 (2001).
Merrill, M. K. et al. Poliovirus receptor CD155-targeted oncolysis of glioma. Neuro Oncol 6, 208–217, doi: 10.1215/S1152851703000577 (2004).
Matusali, G., Potesta, M., Santoni, A., Cerboni, C. & Doria, M. The human immunodeficiency virus type 1 Nef and Vpu proteins downregulate the natural killer cell-activating ligand PVR. J Virol 86, 4496–4504, doi: 10.1128/JVI.05788-11 (2012).
Vassena, L., Giuliani, E., Matusali, G., Cohen, E. A. & Doria, M. The human immunodeficiency virus type 1 Vpr protein upregulates PVR via activation of the ATR-mediated DNA damage response pathway. J Gen Virol 94, 2664–2669, doi: 10.1099/vir.0.055541-0 (2013).
Lindqvist, M. et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection. J Clin Invest 122, 3271–3280, doi: 10.1172/JCI64314 (2012).
Perreau, M. et al. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J Exp Med 210, 143–156, doi: 10.1084/jem.20121932 (2013).
Hecht, F. M. et al. Use of laboratory tests and clinical symptoms for identification of primary HIV infection. AIDS 16, 1119–1129 (2002).
Hunt, P. W. et al. A low T regulatory cell response may contribute to both viral control and generalized immune activation in HIV controllers. PLoS One 6, e15924, doi: 10.1371/journal.pone.0015924 (2011).
Buggert, M. et al. Multiparametric bioinformatics distinguish the CD4/CD8 ratio as a suitable laboratory predictor of combined T cell pathogenesis in HIV infection. J Immunol 192, 2099–2108, doi: 10.4049/jimmunol.1302596 (2014).
Buggert, M., Norstrom, M. M., Salemi, M., Hecht, F. M. & Karlsson, A. C. Functional avidity and IL-2/perforin production is linked to the emergence of mutations within HLA-B*5701-restricted epitopes and HIV-1 disease progression. J Immunol 192, 4685–4696, doi: 10.4049/jimmunol.1302253 (2014).
Amir el, A. D. et al. viSNE enables visualization of high dimensional single-cell data and reveals phenotypic heterogeneity of leukemia. Nat Biotechnol 31, 545–552, doi: 10.1038/nbt.2594 (2013).
Roederer, M., Nozzi, J. L. & Nason, M. C. SPICE: exploration and analysis of post-cytometric complex multivariate datasets. Cytometry A 79, 167–174, doi: 10.1002/cyto.a.21015 (2011).
Sainz, T. et al. The CD4/CD8 ratio as a marker T-cell activation, senescence and activation/exhaustion in treated HIV-infected children and young adults. AIDS 27, 1513–1516, doi: 10.1097/QAD.0b013e32835faa72 (2013).
Appay, V. et al. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat Med 8, 379–385, doi: 10.1038/nm0402-379 (2002).
Appay, V. et al. HIV-specific CD8(+) T cells produce antiviral cytokines but are impaired in cytolytic function. J Exp Med 192, 63–75 (2000).
Betts, M. R. et al. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8+ T cells. Blood 107, 4781–4789, doi: 10.1182/blood-2005-12-4818 (2006).
Almeida, J. R. et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J Exp Med 204, 2473–2485, doi: 10.1084/jem.20070784 (2007).
Lorenzo-Redondo, R. et al. Persistent HIV-1 replication maintains the tissue reservoir during therapy. Nature 530, 51–56, doi: 10.1038/nature16933 (2016).
Lozano, E., Dominguez-Villar, M., Kuchroo, V. & Hafler, D. A. The TIGIT/CD226 axis regulates human T cell function. J Immunol 188, 3869–3875, doi: 10.4049/jimmunol.1103627 (2012).
Gilfillan, S. et al. DNAM-1 promotes activation of cytotoxic lymphocytes by nonprofessional antigen-presenting cells and tumors. J Exp Med 205, 2965–2973, doi: 10.1084/jem.20081752 (2008).
Deeks, S. G. HIV: Shock and kill. Nature 487, 439–440, doi: 10.1038/487439a (2012).
Connick, E. et al. CTL fail to accumulate at sites of HIV-1 replication in lymphoid tissue. J Immunol 178, 6975–6983 (2007).
Fukazawa, Y. et al. B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers. Nat Med 21, 132–139, doi: 10.1038/nm.3781 (2015).
Acknowledgements
We thank the study subjects for their participation, Bo Hejdeman, Marja Ahlqvist, Lise-Lott Nije, Sofia Sandberg, Elisabet Storgärd, Katarina Gyllensten, Maarit Maliniemi and Berit Sundberg for assistance with sample collection. This work was supported by The Swedish Research Council (Grants K2010-56X-20345-04-3 and K2014-57X-22451-01-5), The Swedish Physicians Against AIDS Research Foundation (Fob2013-0008 and Fob2015-0012), Magnus Bergvalls foundation and Karolinska Institutet. The UCSF SCOPE cohort was supported by Delaney AIDS Research Enterprise (DARE; AI096109), the UCSF/Gladstone Institute of Virology & Immunology CFAR (P30 AI027763), and the CFAR Network of Integrated Systems (R24 AI067039).
Author information
Authors and Affiliations
Contributions
A.C.K., M.B. and J.T. designed the study. J.T. and L.S. performed the experiments. J.T. analyzed data, prepared the figures and performed statistical analysis with input from M.B. and A.C.K. Biostatistical analysis (viSNE) and figures were prepared by J.F. with input from O.L. Clinical data and samples were provided by A.N., A.S., G.R.-T., F.M.H. and S.G.D. J.T., M.B. and A.C.K. wrote the manuscript with input from H.G.L. and M.R.B. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Tauriainen, J., Scharf, L., Frederiksen, J. et al. Perturbed CD8+ T cell TIGIT/CD226/PVR axis despite early initiation of antiretroviral treatment in HIV infected individuals. Sci Rep 7, 40354 (2017). https://doi.org/10.1038/srep40354
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40354
This article is cited by
-
Defective HIV-1 genomes and their potential impact on HIV pathogenesis
Retrovirology (2022)
-
Immune mobilising T cell receptors redirect polyclonal CD8+ T cells in chronic HIV infection to form immunological synapses
Scientific Reports (2022)
-
HCV-specific CD4+ T cells of patients with acute and chronic HCV infection display high expression of TIGIT and other co-inhibitory molecules
Scientific Reports (2019)
-
Why and where an HIV cure is needed and how it might be achieved
Nature (2019)
-
T cell senescence and CAR-T cell exhaustion in hematological malignancies
Journal of Hematology & Oncology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
