
Abstract
Melatonin is a widely known hormone in animals. Since melatonin was discovered in plants, more and more studies highlight its involvement in a wide range of physiological processes including plant development and stress responses. Many advances have been made in the terms of melatonin-mediated abiotic stress resistance and innate immunity in plants, focusing on model plants such as rice and Arabidopsis. In this study, 7 melatonin synthesis genes were systematically analyzed in cassava. Quantitative real-time PCR showed that all these genes were commonly regulated by melatonin, flg22, Xanthomonas axonopodis pv manihotis (Xam) and hydrogen peroxide (H2O2). Transient expression in Nicotiana benthamiana revealed the subcellular locations and possible roles of these melatonin synthesis genes. Notably, we highlight novel roles of these genes in hypersensitive-like cell death, as confirmed by the results of several physiological parameters. Moreover, transient expression of these genes had significant effects on the transcripts of reactive oxygen species (ROS) accumulation and defense-related genes, and triggered the burst of callose depositions and papillae-associated plant defense, indicating the possible role of them in plant innate immunity. Taken together, this study reveals the comprehensive transcripts and putative roles of melatonin synthesis genes as well as melatonin in immune responses in cassava.
Similar content being viewed by others


Introduction
In 1958, N-acetyl-5-methoxytryptamine (melatonin) was first discovered in the pineal gland of cow, thereafter melatonin was widely identified in multiple animals1. In 1995, melatonin was identified by two research groups in plants2,3. So far, using gas chromatography/mass spectrometry (GC/MS) and radioimmunoassay, melatonin has been identified in more and more plant species with different levels, including multiple edible plants (banana, cucumber, apple, coffee, corn)4, lupin5, tomato6,7,8,9, rice9,10,11,12,13,14,15, sweet cherry16, Arabidopsis17,18, bermudagrass19, etc. Moreover, it was found that different plant species, plant organs, plant stages, plant location and treatments had significant effects on endogenous melatonin levels19,20,21,22. The wide distribution of melatonin in plants especially in popular beverages and crops makes people can daily take in melatonin from the related products, and this may be benefit to human for the well-known beneficial effects of melatonin on human health23,24,25.
To date, melatonin biosynthetic and metabolic pathway in plants have been revealed. Melatonin biosynthesis begins from tryptophan, the same as it does in animals19,20,21,22,23,24,25,26,27,28. Four sequential enzymatic steps are involved in melatonin biosynthesis in plants, including tryptophan decarboxylase (TDC), tryptamine 5-hydroxylase (T5H), serotonin N-acetyltransferase (SNAT) and N-aceylserotonin O-methyltransferase (ASMT)29,30,31,32,33,34. Moreover, melatonin 2-hydroxylase (M2H) is also responsible for melatonin metabolism, by converting melatonin to 2-hydroxymelatonin35. Genetic modulation by overexpression or knockdown of the transcripts of the above genes responsible for melatonin biosynthesis from animals and plants, including OsTDC, OsASMT, C. reinhardtii AANAT, ovine AANAT and HIOMT, found that the underlying gene expressions are directly associated with endogenous melatonin production19,20,21,22,23,24,25,26,27,34,36,37,38.
Through exogenous application of melatonin or using transgenic plants with affected melatonin levels, previous studies have shown that melatonin plays multiple roles in plants, including senescence39,40,41,42,43, root development44,45,46, floral transition47, fruit ripening48, postharvest physiological deterioration49,50, seed germination51, plant abiotic and biotic stresses52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71. In recent years, many advances have been made in the terms of melatonin-mediated abiotic stress resistance and innate immunity in plants, including salt, drought, cold, heat and oxidative stresses and biotic stress such as pathogen infection19,20,21,22,23,24,25,26,27.
Cassava (Manihot esculenta) can be cultivated under adverse environmental (drought and heat) and nutrient-limited conditions (low phosphorus and low nitrogen), together with its high photosynthetic efficiencies and starch enrichment, making it be considered as an energy crop and important tropical crop72,73,74,75. So far, only two manuscripts reported the in vivo role of melatonin in cassava49,50. Melatonin delays the postharvest physiological deterioration of cassava storage root, through modulation of ROS metabolism, starch metabolism, calcium signaling and mitogen-activated protein kinase (MAPK) cascades49,50. To extend our understanding of melatonin in cassava stress responses, it is essential to reveal the function of melatonin synthesis genes in cassava. In this study, 7 melatonin synthesis genes were cloned and functionally analyzed, especially their possible involvement in immune response. The results may help us in understanding the putative roles of these genes as well as melatonin in immune responses in cassava.
Results
Isolation and expression profiles of melatonin synthesis genes in cassava
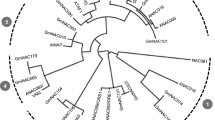
Based on previous studies29,30,31,32,33,34,76, the enzymes responsible for melatonin biosynthesis in plants have been largely revealed. Through genome-wide identification in cassava (Manihot esculenta) annotation database at Phytozome v10.3 (http://www.phytozome.net/cassava.php), 7 cassava genes were identified as candidate genes, including 2 MeTDCs, 1 MeT5H, 1 MeSNAT and 3 MeASMTs (Fig. 1). Moreover, the molecular weight (MW) and theoretical pI of these proteins were analyzed by ProtParam software (http://web.expasy.org/protparam). Then the detailed information of these genes were listed in Table 1, including the locus name, chromosome location, the lengths of peptide, CDS, cDNA and genome of these genes.

The melatonin synthesis genes responsible for melatonin synthesis in cassava.
Through quantitative real-time PCR, we found that the transcript levels of 7 melatonin synthesis genes (MeTDC1, MeTDC2, MeT5H, MeSNAT, MeASMT1, MeASMT2 and MeASMT3) were significantly affected after treatment with flg22, Xanthomonas axonopodis pv manihotis (Xam), melatonin or hydrogen peroxide (H2O2) for 1, 3 and 6 h (Fig. 2A–D). Notably, all these gene transcripts were commonly down-regulated by exogenous melatonin treatment, but were largely up-regulated by Xam and H2O2 at least in one time-point (Fig. 2B–D). However, the transcript levels of these genes were differentially regulated by flg22 treatment (Fig. 2A). Generally, the transcripts levels of these genes were consistent with endogenous melatonin levels in cassava response to flg22, Xam and H2O2 treatments (Supplemental Fig. S1). The common expression profile of 7 melatonin synthesis genes in response to flg22, Xam, melatonin or H2O2 treatments, indicates the possible role of them as well as melatonin in immune response and reactive oxygen species (ROS) signaling in cassava. Thus, melatonin synthesis pathways may play some roles in cassava immune response.

The expression profiles of melatonin synthesis genes in response to flg22 (A), Xam (B), melatonin (C) and H2O2 (D). For the assays, about 30-day-old cassava leaves were treated with water (mock), or 200 μM melatonin, or 10 μM flg22, or Xam infection, or 5 mM H2O2 for 0, 1, 3 and 6 h. The gene transcripts of all genes at 0 h of mock treatment were normalized as 1.0. Asterisk symbols (*) were shown as significant difference at p < 0.05.
Subcellular localization of melatonin synthesis genes in cassava
To investigate the subcellular location of 7 melatonin synthesis genes in cassava, the CDS of these genes were fused in the in-frame with green fluorescent protein (GFP) reporter gene and express in tobacco (N. benthamiana) leaves. After 2 days post infiltration (dpi), we found that the fusion proteins of GFP and 7 melatonin synthesis proteins (MeTDC1, MeTDC2, MeT5H, MeSNAT, MeASMT1, MeASMT2 and MeASMT3) exhibited obvious green signals in both nucleus and cell membrane in the leaf cells (Fig. 3).

Subcellular localization of melatonin synthesis genes in cassava.
Transient expression of GV3101 cell culture harbouring the 35S::GFP, 35S::GFP-MeTDC1, 35S::GFP-MeTDC2, 35S::GFP-MeSNAT, 35S::GFP-MeT5H, 35S::GFP-MeASMT1, 35S::GFP-MeASMT2 and 35S::GFP-MeASMT3 plasmids in tobacco leaves. Cell nuclei was stained by 1 μg/ml 4′,6-diamidino-2-phenylindole (DAPI). Bar = 25 μm.
Transient expression of melatonin synthesis genes triggers hypersensitive response-like cell death
Interestingly, we observed that the transient expression of all these genes resulted in obvious cell death and hypersentive response (HR) symptoms after infiltration in tobacco (N. benthamiana) leaves in comparison to the overexpression of GFP (control) (Fig. 4A). HR is a common type of programmed cell death (PCD) as well as a marker feature of plant immune response, which are largely related to ROS accumulation77,78. In this study, tobacco leaves expressing melatonin synthesis genes exhibited significantly higher levels of H2O2 and superoxide radical (O2•−) than those from leaves expressing GFP alone, as evidenced by DAB, NBT staining (Fig. 4B,C) and the quantifications of endogenous H2O2 and O2•− (Fig. 4D,E). Moreover, the electrolyte leakage (EL) of leaf discs from tobacco leaves expressing these genes was significantly higher than those from leaves expressing GFP (Fig. 4F), indicating the significant increase of ion leakage triggered by these genes transient expression. Consistently, malondialdehyde (MDA), which is a lipid peroxidation caused by intracellular ROS accumulation, displayed high level in tobacco leaves expressing these genes than GFP (Fig. 4G). These results indicate the in vivo roles of melatonin synthesis genes in HR, immune response and underlying ROS modulation.

Transient expression of melatonin synthesis genes in tobacco leaves on hypersensitive response-like cell death.
(A) Symptoms of cell death in leaves were visualized under UV light using the ChemiDoc Imaging System. (B,C) DAB staining for endogenous H2O2 level (B) and NBT staining for endogenous O2•− level (C) in tobacco leaf discs expressing different plasmids. (D–G) Quantification of H2O2 (D), O2•− (E), EL (F) and MDA (G) contents in leaf discs expressing different plasmids. Asterisk symbols (*) were shown as significant difference at p < 0.05.
Modulation of melatonin synthesis genes expression regulates melatonin content
To reveal the possible relationship between melatonin synthesis genes-regulated HR and endogenous melatonin level, we investigate the effect of these genes transient expression on melatonin level. Through quantification by ELISA, we found that tobacco leaves expressing melatonin synthesis genes exhibited significantly higher levels of endogenous melatonin than those from leaves expressing GFP alone (Fig. 5), suggesting the possible involvement of melatonin in these genes -mediated HR and immune response. Although MeT5H gene is missing at least 70 aa at the N-terminus when compared to other MeT5H homologs such as OsT5H55, it had significant effect on melatonin synthesis, indicating the difference of T5H in different plant species.

The endogenous melatonin levels induced by the expression of melatonin synthesis genes expression.
Asterisk symbols (*) were shown as significant difference at p < 0.05.
Modulation of melatonin synthesis genes expression triggers immune response
To further dissect the possible mechanisms of melatonin synthesis genes-mediated ROS accumulation and immune response, we analyzed the effects of these genes expression on the transcripts of several genes in ROS and defense pathways. Through quantitative real-time PCR, we found that these genes transient expression in tobacco leaves significantly increased the transcript levels of both ROS-related genes (superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX)) (Fig. 6) and defense-related genes (RbohA, RbohB, pathogensis-related gene 1 (PR1), PR2 and PR5) (Fig. 7). As common feature of defense response, ROS accumulation and PCD are involved in immune response. These results indicate the involvement of melatonin synthesis genes in both ROS and defense signalings.

Modulation of melatonin synthesis genes expression regulates the transcripts of ROS-related genes.
The gene expression was assayed at 3 dpi (A) and 6 dpi (B). The transcript levels of every gene in mock (GFP) transformed leaves were set as 1. Asterisk symbols (*) were shown as significant difference at p < 0.05.

Modulation of melatonin synthesis genes expression regulates the transcripts of several genes in plant immunity.
The gene expression was assayed at 3 dpi (A) and 6 dpi (B). The transcript levels of every gene in mock (GFP) transformed leaves were set as 1. Asterisk symbols (*) were shown as significant difference at p < 0.05.
Additionally, the effects of melatonin synthesis genes transient expression on callose depositions were also analyzed. As shown in Fig. 8, the tobacco leaves expressing melatonin synthesis genes exhibited significantly more callose depositions than those from leaves expressing GFP alone. This result indicates that melatonin synthesis genes might be involved in the modulation of callose-associated cell wall and papillae-associated plant defense.

The effects of melatonin synthesis genes transient expression on callose depositions.
White dots in the figures indicate callose depositions staining with aniline blue. Bar = 500 μm.
Discussion
In the long period of evolution, plants have developed complicated mechanisms to survive and thrive in response to various environmental stresses and microbial pathogens26. Briefly, different stress signals are first perceived by membrane receptors, thereafter are activated by several secondary messengers such as calcium, abscisic acid (ABA), nitric oxide (NO) and H2O2. Then the signal transduction by secondary messengers leads to the activation of protein kinases, transcription factors, stress-responsive genes and physiological responses, eventually resulting in protective responses18,19,26,66,67,68,69,70. The significant elevations of endogenous melatonin in plant early stress signaling indicated that melatonin may serve as an important early messenger in plant stress response20,21,22,23,24,25,26,27.
As reviewed by Zhang et al.26, most of previous studies attribute the protective role of melatonin in stress resistance to the alleviation of stress-triggered ROS production and the activation of antioxidants. As it does in animals, melatonin has been shown to detoxify the H2O2, O2•−, singlet oxygen (1O2), hydroxyl radical (•OH), peroxynitrite anion (ONOO−), and hypochlorous acid (HOCl) directly19,20,21,22,23,24,25,26,27. Additionally, melatonin also has indirect antioxidative actions through activating the activities of antioxidant enzymes including CAT, SOD, glutathione reductase (GR), and glutathione peroxidase (GPX), all of which remove toxic reactants metabolically19,20,21,22,23,24,25,26,27. These actions of melatonin including potent antioxidant and radical free scavenger may largely contribute to melatonin-conferred stress resistance in plants.
Recently, our studies together with other studies provided some clues for the molecular mechanisms of melatonin-mediated stress responses in plants18,19,26,62,63,64,65,66,67. ROS burst and associated changes such as the transcripts of defense genes play important roles in plant immune response, especially in plant-pathogen interaction77,78. However, the involvement of melatonin in hypersensitive-like cell death and underlying ROS accumulation remain unknown. Herein, the identification and functional analysis of melatonin synthesis genes in cassava provided direct link between melatonin and immune response, as well as the underlying mechanism of these genes in programmed-like cell death and ROS accumulation. ROS is important signal molecules in signal transduction, serving as second messengers, whereas ROS overproduction under stress conditions results in serious cell damage. Interestingly, melatonin also has dual roles in regulating ROS. On one hand, stress induced melatonin relieves oxidative stress damage by decreasing excess ROS18,19,26,62,63,64,65,66,67. On the other hand, melatonin induces the ROS level to activate the downstream responses in the early stress response. Thus, the dual roles of melatonin in regulating ROS further indicate the protective effect of melatonin in various stress response.
In this study, we successfully identified 7 melatonin synthesis genes (2 MeTDCs, 1 MeT5H, 1 MeSNAT and 3 MeASMTs) in cassava and cloned the coding sequences of them (Fig. 1 and Table 1). The responses of these genes under different treatments (flg22, Xam, melatonin and H2O2) were analyzed through quantitative real-time PCR, the common expression profile of these genes indicates the possible role of them as well as melatonin in immune response and ROS signaling in cassava (Fig. 2). As a rate limited enzyme of melatonin synthesis, gene expression of MeSANT was not up-regulated by the flg22 and H2O2. However, the endogenous melatonin levels were elevated under the treatments of flg22 and H2O2, as well as the response in hypersensitive-like cell death. On one hand, the transcript level is not always consistent with enzyme activity of MeSNAT, and the post-transcriptional regulation and post-translational regulation may also result in the issue. On the other hand, MeSNAT may only responsible for part melatonin synthesis in cassava under these stress conditions, and more other rate limiting enzymes of melatonin synthesis need to be further isolated. More importantly, we identified the novel role of transient expressing melatonin synthesis genes in hypersensitive-like cell death in leaves of N. benthamiana, depending on ROS accumulation and endogenous melatonin (Figs 4 and 5). SOD and CAT are major antioxidant metabolic enzymes by catalysing O2−. into H2O2 and O278,79. RbohA and RbohB are also important regulators of not only H2O2 accumulation, but also plant immunity. PR1, PR2 and PR5 are widely known marker genes of innate immune response66,79,80. Further investigation of gene expression indicated that transient expressing melatonin synthesis genes induced the transcription of both ROS- and defense-related marker genes (Figs 6 and 7), suggesting that these genes might exert its function through ROS accumulation and PCD as well as immune response. Additionally, the tobacco leaves expressing these genes triggered the burst of callose depositions (Fig. 8), suggesting that these genes might be involved in the modulation of callose-associated cell wall and papillae-associated plant defense.
Pathogen associated molecular patterns (PAMPs)-triggered immunity (PTI) can resist the incidence of most pathogenic microbes, and plays important roles in plant immune response77,78,79. In this study, melatonin synthesis genes expression plays common roles in some PTI responses, including the transcript profile in response to flg22 and Xam, ROS accumulation, hypersensitive-like cell death, defense-related gene expression and callose deposition. HR symptoms with accumulation of ROS usually occur in biotic or abiotic stresses in plants, but it also happen as a constitutive activated protective mechanism in recent studies79. The induction of PTI response as well as HR by melatonin synthesis genes indicates the basal immunity triggered by them.
Taken together, this study shows the comprehensive transcripts and the putative roles of melatonin related genes as well as melatonin in immune responses in cassava. The results may also provide important candidate genes for genetic breeding to improved disease resistance of cassava.
Methods
Plant materials and growth conditions
In this study, cassava plants of South China 124 (SC124) variety were grown in pots with soil and vermiculite (1:1) (pH 5.7) in the green house, which was controlled under 12 h light/28 °C and 12 h dark/26 °C cycles, at the irradiance of about 120–150 μmol quanta m−2 s−1. Hoagland’s solution was watered twice every week to keep the well growth of cassava plants.
RNA isolation and quantitative real-time PCR
Plant leaves were harvest for total RNA isolation using AxyPrepTM Multisource Total RNA Miniprep Kit (AYXGEN-09113KD1, Santa Clara, California, USA) according to the manufacturer’s instruction81. First-strand cDNA synthesis was performed using reverse transcriptase (Thermo-K1622, Waltham, Massachusetts, USA) as Shi et al. described previously19. Thereafter, the diluted cDNA and SYBR® Premix DimerEraserTM (TaKaRa Biotechnology-RR091A, Dalian city, Liaoning, China) were used for quantitative real-time PCR in LightCycler® 96 Real-Time PCR System (Roche, Basel, Switzerland). All gene transcripts were normalized to elongation factor 1α (EF1α) using comparative 2−ΔΔCT method. The primers used in this study were listed in Supplemental Table S1.
Plasmid construction and transient expression in tobacco leaves
The coding regions of MeTDC1, MeTDC2, MeSNAT, MeT5H, MeASMT1, MeASMT2 and MeASMT3 were amplified and cloned into downstream of GFP through restriction enzyme digestion and ligase in the pEGAD vector the control of CaMV 35S promoter82. The corresponding primers for gene clone were listed in Table S2. After sequencing for confirmation, the 35S::GFP-MeHSP90s plasmids were transformed into Agrobacterium tumefaciens strain GV3101 and used for the subcellular localization and overexpressing analysis.
The equal volumes of GV3101 cell culture harbouring the 35S::GFP, 35S::GFP-MeTDC1, 35S::GFP-MeTDC2, 35S::GFP-MeSNAT, 35S::GFP-MeT5H, 35S::GFP-MeASMT1, 35S::GFP-MeASMT2 and 35S::GFP-MeASMT3 plasmids and P19 were co-infected into 30-day-old tobacco leaves as previously described83. For each vector construct, 20 independent leaves of five independent tobacco (N. benthamiana) plants (four leaves per plant) were used for GFP and physiological assays. After two days, GFP signals in the transformed tobacco leaves were observed using a confocal laser-scanning microscope (TCS SP8, Leica, Heidelberg, Germany). Physiological assays were performed at different time points after infection.
Determination of physiological parameters
For the ROS staining, H2O2 and O2•− were stained in diaminobenzidine (DAB) solution (pH 3.8) and nitro blue tetrazolium (NBT) solution (pH 7.8) and further decolorized in 70% (v/v) ethanol as Shi et al. described19. Moreover, H2O2 and O2•− contents were quantified by examining the peroxide-titanium from the titanium sulphate regent and antibody-antigen-enzyme-antibody complex from the plant O2•− ELISA Kit as Shi et al. described19.
The EL was calculated as the relative value of initial conductivity (Ci) to the maximum conductivity (Cmax), which were assayed in the mixture of plant leaves-deionized water after 6 hr of shaken and after boiled and cooling, respectively.
The MDA was determined by examining the absorbance of the supernatant of leaves samples in the thiobarbituric acid (TBA) regent as Shi et al. described19.
Callose staining
Tobacco leaves were harvest after infiltration GV3101 cell culture harbouring the construction, cleared in alcoholic lactophenol solution and staining with 0.01% (w/v) aniline blue as Hauck et al. described84. Images were examined using a fluorescence microscope (DM6000B, Leica, Heidelberg, Germany), and the callose depositions were quantified using the ImageJ software.
Significant difference analysis
In this study, all experiments were repeated with three biological experiments. All data were shown as average value ± SD of three biological experiments. ANOVA and student’s t-test were performed to analyze the significant difference in comparison to mock treatment, and asterisk symbols (*) were shown as significant difference at p < 0.05.
Additional Information
How to cite this article: Wei, Y. et al. Comprehensive transcriptional and functional analyses of melatonin synthesis genes in cassava reveal their novel role in hypersensitive-like cell death. Sci. Rep. 6, 35029; doi: 10.1038/srep35029 (2016).
References
Lerner, A. B. et al. Isolation of melatonin, the pineal gland factor that lightening melanocytes. J. Am. Chem. Soc. 81, 2587 (1958).
Dubbels, R. et al. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 18, 28–31 (1995).
Hattori, A. et al. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Inter. 35, 627–634 (1995).
Manchester, L. C. et al. High levels of melatonin in the seeds of edible plants: the seed of edible plants: possible function in germ tissue protection. Life Sci. 67, 3023–3029 (2000).
Hernández-Ruiz, J., Cano, A. & Arna, M. B. Melatonin: a growth-stimulating compound present in lupin tissues. Planta. 220, 140–144 (2004).
Hernández-Ruiz, J., Cano, A. & Arna, M. B. Melatonin acts as a growth-stimulating compound in some monocot species. J. Pineal Res. 39, 137–142 (2005).
Okazaki, M. & Ezura, H. Profiling of melatonin in model tomato (Solanum lycopersicum L.) cultivar Micro-Tom. J. Pineal Res. 43, 338–343 (2009).
Okazaki, M. et al. Cloning and characterization of a Chlamydomonas reinhardtii cDNA arylalkylamine N -acetyltransferase and its use in the genetic engineering of melatonin content in the Micro-Tom tomato. J. Pineal Res. 43, 373–382 (2009).
Okazaki, M. et al. Lowering intercellular melatonin levels by transgenic analysis of indoleamine 2, 3-dioxygenase from rice in tomato plants. J. Pineal Res. 49, 239–247 (2010).
Kang, K. et al. Molecular cloning of a plant N-acetylserotonin methyltransferase and its expression characteristics in rice. J. Pineal Res. 50, 304–309 (2011).
Byeon, Y. et al. Light-regulated melatonin biosynthesis in rice during the senescence process in detached leaves. J. Pineal Res. 53, 107–111 (2012).
Byeon, Y. et al. Microarray analysis of genes differentially expressed in melatonin-rich transgenic rice expressing a sheep serotonin N-acetyltransferase. J. Pineal Res. 55, 357–363 (2013).
Byeon, Y. et al. Elevated production of melatonin in transgenic rice seeds expressing rice tryptophan decarboxylase. J. Pineal Res. 56, 275–282 (2014).
Byeon, Y. & Back, K. An increase in melatonin in transgenic rice causes pleiotropic phenotypes, including enhanced seedling growth, delayed flowering, and low grain yield. J. Pineal Res. 56, 408–414 (2014).
Byeon, Y. & Back, K. Melatonin synthesis in rice seedlings in vivo is enhanced at high temperatures and under dark conditions due to increased serotonin N-acetyltransferase and N-acetylserotonin methyltransferase activities. J. Pineal Res. 56, 189–195 (2014).
Zhao, Y. et al. Melatonin and its potential biological functions in the fruits of sweet cherry. J. Pineal Res. 55, 79–88 (2013).
Arnao, M. B. & Hernández-Ruiz, J. Melatonin: plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 19, 789–797 (2014).
Shi, H. & Chan, Z. The cysteine2/histidine2-type transcription factor ZINC FINGER OF ARABIDOPSIS THALIANA 6-activated C-REPEAT-BINDING FACTOR pathway is essential for melatonin-mediated freezing stress resistance in Arabidopsis. J. Pineal Res. 57, 185–191 (2014).
Shi, H. et al. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 66, 681–694 (2015).
Reiter, R. J., Tan, D. X. & Galano, A. Melatonin: exceeding expectations. Physiology (Bethesda) 29, 325–333 (2014).
Tan, D. X. et al. Fundamental issues related to the origin of melatonin and melatonin isomers during evolution: relation to their biological functions. Int. J. Mol. Sci. 15, 15858–15890 (2014).
Tan, D. X. et al. Melatonin as a potent and inducible endogenous antioxidant synthesis and metabolism. Molecules 20, 18886–18906 (2015).
Reiter, R. J. et al. Phytomelatonin: assisting plants to survive and thrive. Molecules 20, 7396–7437 (2015).
Arnao, M. B. & Hernández-Ruiz, J. Functions of melatonin in plants: a review. J. Pineal Res. 59, 133–150 (2015).
Hardeland, R. Melatonin in plants-diversity of levels and multiplicity of functions. Front Plant Sci. 7, 198 (2016).
Zhang, N. et al. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 66, 647–656 (2015).
Hardeland, R. Melatonin in plants and other phototrophs: advances and gaps concerning the diversity of functions. J. Exp. Bot. 66, 627–646 (2015).
Kolár, J. & Machácková, I. Melatonin in higher plants: occurrence and possible functions. J. Pineal Res. 39, 333–341 (2005).
Kang, S. et al. Characterization of rice tryptophan decarboxylases and their direct involvement in serotonin biosynthesis in transgenic rice. Planta 227, 263–272 (2007).
Posmyk, M. M. et al. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 45, 24–31 (2008).
Zuo, B. et al. Overexpression of MzASMT improves melatonin production and enhances drought tolerance in transgenic Arabidopsis thaliana plants. J. Pineal Res. 57, 408–417 (2014).
Kang, K. et al. Molecular cloning of rice serotonin N-acetyltransferase, the penultimate gene in plant melatonin biosynthesis. J. Pineal Res. 55, 7–13 (2013).
Park, S., Byeon, Y. & Back, K. Function analyses of three ASMT gene family members in rice plants. J. Pineal Res. 55, 409–415 (2013).
Park, S. et al. Melatonin-rich transgenic rice plants exhibit resistance to herbicide-induced oxidative stress. J. Pineal Res. 54, 258–263 (2013).
Byeon, Y. & Back, K. Molecular cloning of melatonin 2-hydroxylase responsible for 2-hydroxymelatonin production in rice (Oryza sativa). J. Pineal Res. 58, 343–351 (2015).
Kang, K. et al. Enhanced production of melatonin by ectopic overexpression of human serotonin N-acetyltransferase plays a role in cold resistance in transgenic rice seedlings. J. Pineal Res. 49, 176–182 (2010).
Wang, L. et al. Change in melatonin levels in transgenic ‘Micro-Tom’ tomato over-expressing ovine AANAT and ovine HIOMT genes. J. Pineal Res. 56, 126–133 (2014).
Lee, H. Y. et al. Arabidopsis serotonin N-acetyltransfase knockout mutant plants exhibit decreased melatonin and salicylic acid levels resulting in susceptibility to an avirulent pathogen. J. Pineal Res. 58, 291–299 (2015).
Wang, P. et al. Delayed senescence of apple leaves by exogenous melatonin treatment: toward regulating the ascorbate–glutathione cycle. J. Pineal Res. 53, 11–20 (2012).
Wang, P. et al. Delay in leaf senescence of Malus hupehensis by long-term melatonin application is associated with its regulation of metabolic status and protein degradation. J. Pineal Res. 55, 424–434 (2013).
Wang, P. et al. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 54, 292–302 (2013).
Kang, K. et al. Senescence-induced serotonin biosynthesis and its role in delaying senescence in rice leaves. Plant Physiol. 150, 1380–1393 (2009).
Shi, H. et al. INDOLE-3-ACETIC ACID INDUCIBLE 17 positively modulates natural leaf senescence through melatonin-mediated pathway in Arabidopsis. J. Pineal Res. 58, 26–33 (2015).
Park, S. & Back, K. W. Melatonin promotes seminal root elongation and root growth in transgenic rice after germination. J. Pineal Res. 53, 385–389 (2012).
Pelagio-Flores, R. et al. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 53, 279–288 (2012).
Zhang, N. et al. The RNA-seq approach to discriminate gene expression profiles in response to melatonin on cucumber lateral root formation. J. Pineal Res. 56, 39–50 (2014).
Shi, H. et al. Melatonin mediates the stabilization of DELLA proteins to repress the floral transition in Arabidopsis. J. Pineal Res. 60, 373–379 (2016).
Sun, Q. et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 66, 657–668 (2015).
Ma, Q. et al. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal Res. 60, 424–434 (2016).
Hu, W. et al. Comparative physiological and transcriptomic analyses reveal the actions of melatonin in the delay of postharvest physiological deterioration of cassava. Front Plant Sci. 7, 736 (2016).
Zhang, N. et al. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J. Pineal Res. 54, 15–23 (2013).
Meng, J. F. et al. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: antioxidant metabolites, leaf anatomy, and chloroplast morphology. J. Pineal Res. 57, 200–212 (2014).
Wei, W. et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 66, 695–707 (2014).
Zhang, H. J. et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 57, 269–279 (2014).
Liang, C. et al. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 59, 91–101 (2015).
Posmyk, M. M. et al. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress. J. Pineal Res. 46, 214–223 (2009).
Bajwa, V. S. et al. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 56, 238–245 (2014).
Turk, H. et al. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 74, 139–152 (2014).
Tiryaki, I. & Keles, H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J. Pineal Res. 52, 332–339 (2012).
Shi, H. et al. Comparative physiological and proteomic analyses reveal the actions of melatonin in the reduction of oxidative stress in Bermuda grass (Cynodon dactylon (L). Pers.). J. Pineal Res. 59, 120–131 (2015).
Byeon, Y. et al. Coordinated regulation of melatonin synthesis and degradation genes in rice leaves in response to cadmium treatment. J. Pineal Res. 58, 470–478 (2015).
Yin, L. et al. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J. Pineal Res. 54, 426–434 (2013).
Lee, H. Y., Byeon, Y. & Back, K. Melatonin as a signal molecule triggering defense responses against pathogen attack in Arabidopsis and tobacco. J. Pineal Res. 57, 262–268 (2014).
Zhao, H. et al. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis thaliana. J. Pineal Res. 59, 109–119 (2015).
Li, C. et al. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 66, 669–680 (2014).
Shi, H. et al. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 59, 102–108 (2015).
Shi, H. et al. Melatonin induces class A1 heat shock factors (HSFA1s) and their possible involvement of thermotolerance in Arabidopsis. J. Pineal Res. 58, 335–342 (2015).
Shi, H. et al. Melatonin induces the transcripts of CBF/DREBs and their involvement in abiotic and biotic stresses in Arabidopsis. J. Pineal Res. 59, 334–342 (2015).
Qian, Y. et al. Comparative metabolomic analysis highlights the involvement of sugars and glycerol in melatonin-mediated innate immunity against bacterial pathogen in Arabidopsis. Sci. Rep. 5, 15815 (2015).
Shi, H., Wei, Y. & He, C. Melatonin-induced CBF/DREB1s are essential for diurnal change of disease resistance and CCA1 expression in Arabidopsis. Plant Physiol. Biochem. 100, 150–155 (2016).
Lee, H. Y. & Back, K. Mitogen-activated protein kinase pathways are required for melatonin-mediated defense responses in plants. J. Pineal Res. 60, 327–335 (2016).
Wang, W. et al. Cassava genome from a wild ancestor to cultivated varieties. Nat. Commun. 5, 5110 (2014).
Zhao, P. et al. Analysis of different strategies adapted by two cassava cultivars in response to drought stress: ensuring survival or continuing growth. J. Exp. Bot. 66, 1477–1488 (2015).
Hu, W. et al. Genome-wide gene phylogeny of CIPK family in cassava and expression analysis of partial drought-induced genes. Front Plant Sci. 6, 914 (2016).
Hu, W. et al. Genome-wide characterization and analysis of bZIP transcription factor gene family related to abiotic stress in cassava. Sci. Rep. 6, 22783 (2016).
Tan, D. X. et al. On the significance of an alternate pathway of melatonin synthesis via 5-methoxytryptamine: comparisons across species. J. Pineal Res., doi: 10.1111/jpi.12336 (2016).
Lamb, C. & Dixon, R. A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 251–275 (1997).
Apel, K. & Hirt, H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 55, 373–399 (2004).
Kwon, S. I., Cho, H. J., Kim, S. R. & Park, O. K. The Rab GTPase RabG3b positively regulates autophagy and immunity-associated hypersensitive cell death in Arabidopsis. Plant Physiol. 161, 1722–1736 (2013).
Chen, B. et al. Identification, cloning and characterization of R2R3-MYB gene family in canola (Brassica napus L.) identify a novel member modulating ROS accumulation and hypersensitive-like cell death. DNA Res. 23, 101–114 (2016).
Wei, Y. et al. Genome-wide identification and expression analysis of the WRKY gene family in cassava. Front Plant Sci. 7, 25 (2016).
Cutler, S. R., Ehrhardt, D. W., Griffitts, J. S. & Somerville, C. R. Random GFP::cDNA fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. P. Natl. Acad. Sci. USA 97, 3718–3723 (2004).
Sparkes, I. A., Runions, J., Kearns, A. & Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 1, 2019–2025 (2006).
Hauck, P., Thilmony. R. & He, S. Y. A. Pseudomonas syringae type III effector suppresses cell wall-based extracellular defense in susceptible Arabidopsis plants. Proc. Natl. Acad. Sci. USA 100, 8577–8582 (2003).
Acknowledgements
This research was supported by the National Natural Science Foundation of China (No. 31570249), the scientific research foundation of higher education in Hainan province (the education curriculum reform program, title: Research on the overall optimization of the curriculum system and teaching content of Hainan province excellent course-gene engineering, No.Hnjg2016-10), a central financial support to enhance the comprehensive strength of the central and western colleges and universities, the startup funding and the scientific research foundation of Hainan University (No. kyqd1531) and the education curriculum reform program of Hainan University (No. hdjy1601) to Haitao Shi.
Author information
Authors and Affiliations
Contributions
H.S. conceived and directed this study, designed the experiments, wrote and revised the manuscript; Y.W. performed the experiments, analyzed the data, wrote and revised the manuscript; W.H. analyzed the data and revised the manuscript; Q.W. performed the fluorescence analysis and analyzed the data; W.L., C.W., H.Z., Y.Y. and X.L. performed the experiments; C.H. designed the experiments and revised the manuscript. All authors approved the manuscript and the version to be published.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wei, Y., Hu, W., Wang, Q. et al. Comprehensive transcriptional and functional analyses of melatonin synthesis genes in cassava reveal their novel role in hypersensitive-like cell death. Sci Rep 6, 35029 (2016). https://doi.org/10.1038/srep35029
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep35029
This article is cited by
-
Metabolite profiling and transcriptome analyses reveal novel regulatory mechanisms of melatonin biosynthesis in hickory
Horticulture Research (2021)
-
Novel Role of Ethanol in Delaying Postharvest Physiological Deterioration and Keeping Quality in Cassava
Food and Bioprocess Technology (2019)
-
Genome-wide analysis of autophagy-related genes in banana highlights MaATG8s in cell death and autophagy in immune response to Fusarium wilt
Plant Cell Reports (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
