
Abstract
Fishes have adapted a number of different behaviors to move out of the water, but none have been described as being able to walk on land with a tetrapod-like gait. Here we show that the blind cavefish Cryptotora thamicola walks and climbs waterfalls with a salamander-like diagonal-couplets lateral sequence gait and has evolved a robust pelvic girdle that shares morphological features associated with terrestrial vertebrates. In all other fishes, the pelvic bones are suspended in a muscular sling or loosely attached to the pectoral girdle anteriorly. In contrast, the pelvic girdle of Cryptotora is a large, broad puboischiadic plate that is joined to the iliac process of a hypertrophied sacral rib; fusion of these bones in tetrapods creates an acetabulum. The vertebral column in the sacral area has large anterior and posterior zygapophyses, transverse processes and broad neural spines, all of which are associated with terrestrial organisms. The diagonal-couplet lateral sequence gait was accomplished by rotation of the pectoral and pelvic girdles creating a standing wave of the axial body. These findings are significant because they represent the first example of behavioural and morphological adaptation in an extant fish that converges on the tetrapodal walking behaviour and morphology.
Similar content being viewed by others

Introduction
The vertebrate transition from finned to limbed appendages in the Devonian period resulted in the earliest records of quadrupedal walking on terra firma. However, fishes, which include more than 35,000 species, have evolved terrestrial locomotion, in one form or another, multiple times. Several extant fishes have a number of morphological and behavioural traits that facilitate moving out of water to escape predation, find food or new habitats, or lay eggs1,2,3. The most simplistic and least anatomically derived method to move across a dry horizontal surface is to undulate or flip the body by modifying the same motor programs that facilitate swimming and escape responses in water, as is seen in eels4, killifishes2,5 and sticklebacks6. Some fishes are able to walk underwater using their pectoral fins and pelvic fins (e.g. frogfish7); however, in submerged walking the bodyweight is supported by the fluid around the organism. Polypterus8, mudskippers9,10 and walking catfish11 move on land via crutching or lateral pushing by the posterior body and tail to rotate forward over their pectoral fins. Lungfish can walk on a horizontal surface using primarily pelvic driven alternating fin movements12. Fishes known for vertical climbing, such as Hawaiian waterfall climbing gobies, use either intermittent, rapid axial undulation (Awaous guamensis and Lentipes concolor) or oral and pelvic suction (Sicyopterus stimpsoni)13. Notably, none of these fishes walk with a diagonal-couplets lateral sequence gait on land, which has been described as a purely tetrapodal innovation.
The blind cavefish Cryptotora thamicola (Cypriniformes: Balitoridae; Fig. 1) can walk up rocks in fast-flowing water and on wet surfaces in air. This fish is a troglobitic hillstream loach endemic to the Tham Maelana and Tham Susa karst formation in northern Thailand. Cryptotora are found only in rapids and not in lentic pools14. They are commonly observed climbing steep rock surfaces in fast-flowing waterfalls created by basalt or andesite intrusions (Supplementary Video 1)14. While it is anecdotally known that these fish can walk, the rare and protected status of these fish has limited research into the functional morphology of their walking behaviour. Here we show that the blind cavefish Cryptotora thamicola convergently evolved a diagonal-couplets lateral sequence gait method of walking and shares morphological features typically attributed to the evolution of terrestriality in early tetrapods.

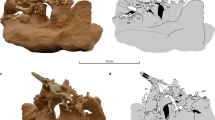
Cryptotora thamicola.
Dorsal view, resting on the bottom of glass tank. Photo by D.S.
Results and Discussion
Pelvic girdle morphology
We found that Cryptotora had a pelvic girdle that is connected to the axial skeleton via fusion with a hypertrophied rib of the seventh vertebra (Fig. 2A–C, Supplementary Video 2). The basipterygium, or puboischiadic plate (pu, tan, Fig. 2C), was broad in both anteroposterior and lateral directions and more heavily ossified than is typical of fishes (Fig. 3), with a large concavity for ventral attachment of muscles to the large processes at the base of the fin rays (pf, blue, Fig. 2B,C). The heads of the fin rays articulated with the lateral edges of the ischium.

Computed microtomography scan (voxel size = 8 μm) of Cryptotora thamicola (47 mm total length).
(A) transverse view of pelvic girdle, (B) dorsal (left) and ventral (right) view pelvic girdle (cranial to top), (C) anterolateral view of pelvic girdle. il, iliac region (dark purple); im, intermuscular bone (light purple); is, ischial region of puboischiadic plate (tan); pf, pelvic fin (blue); pu, pubic region of puboischiadic plate (tan); sr, sacral ribs (dark purple). Vertebrae numbers 6,7,8 designate position from skull, (D) close-up image of acetabular symphysis, (E) close up image of obturator foramen, of.

Computed microtomography scan (voxel size = 7 μm) of goldfish Carassius auratus pelvis (38 mm total length).
(A) Transverse view of puboischiadic plate (basipterygium, tan), pelvic fins (blue) and axial skeleton (grey), (B) dorsal (left) and ventral (right) view pelvis and axial skeleton (cranial to top), (C) anterolateral view of pelvis and axial skeleton. (D) dorsal view of pelvis and fins.
The ribs of Cryptotora were broader and extended more laterally than is typical of other fishes; the common goldfish (Carassius auratus), also of the order Cypriniformes, is shown for comparison (Figs 2 and 3). Notably, the goldfish pelvis did not articulating with the ribs, but was held in a muscular sling; this is typically the case for all other fishes in which the pelvis is not fused to the pectoral girdle anteriorly. In Cryptotora, the first seven ribs were broad and flat with an arch reflecting the dorsoventral compression of the fish body. The seventh rib (sr, purple, Fig. 2C) was more heavily calcified than more anterior ribs, which were only calcified around their edges. In addition, the seventh rib had a large, broad, flared process similar in appearance to the iliac crest which supports attachment of large hip flexors in tetrapods. The distal end of the seventh rib (originating on the seventh vertebrae) formed a ventrally oriented iliac process that passed through an arch formed by the pubis and ischium to fuse on the anterolateral aspect of the puboischiadic plate (Fig. 2C). The area of this arch where the puboischiadic plate and iliac process of the sacral rib united was the acetabular symphysis. (Fig. 2D). While extant fishes lack femora, they have historically had the posterior ridge of the pelvis, where the pelvic fins articulate, identified as the area of the acetabulum15,16. Exctinct fishes Gooloogongia, Eusthenopteron and Panderichthyes posessed more cup-like acetabula in a similar posterior position, despite lacking limbs16,17. In contrast, Cryptotora had an acetabular symphysis separate from and anterior to the pelvic fin articulation with the puboischiadic plate. Also visible posterior to the acetabular symphysis was the obturator foramen, an opening formed between the ischium and pubis through which blood vessels and nerves may pass (Fig. 2E). The eighth rib did not originate off the vertebral centra as the seventh rib did, but instead originated off a transverse process of the eighth vertebra and extended laterally and hooked anteriorly, which may provide additional area for muscle attachment on the dorsal aspect of the fin.
Based on their positions, we presume that the dorsolateral projections from the vertebral centra are intermuscular bones (im, light purple, Fig. 2C) - intramembranous ossifications which form in myosepta as a result of mechanical stress18. The intermuscular bone associated with the seventh vertebrae was different than the rest, in that it was attached to the dorsal aspect of the sacral rib and not the vertebral centra. Anterior to the pelvis, the intermuscular bones branched off of the neural arches. If these bones are in fact, not intermuscular, then they are skeletal structures that have not previously been identified in fishes.
The vertebral column of Cryptotora shared several features associated with the evolution of terrestriality and the functional need to support the body weight of an organism outside of water: in particular, the wide neural spines, zygapophyses and transverse processes of Cryptotora are features that stabilize the vertebral column in terrestrial vertebrates19,20. Broad neural spines are typically associated with muscle attachment along the vertebral column15. Large transverse processes extended laterally off the vertebral centra and zygapophyses extended in anterior and posterior directions (Fig. 2C). The centra of the sacral vertebrae were tapered ventroposteriorly, resulting in narrowing of the notochord which passed through the center.
While a study of the development of the pelvic girdle in this cavefish has not yet been possible, it appears that structurally, the pelvic girdle of Cryptotora converges on tetrapodal morphology that supports muscular attachment and transfer of forces for terrestrial walking16,21. In other fishes, including those that can walk by crutching or lunging, there is no bony connection between the pelvis and vertebral column and the pelvis is held in place by a muscular sling (Fig. 3)16 or fused anteriorly to the pectoral girdle, as in the mudskipper22. However, in tetrapods, the ilium originates from the pubis and extends dorsally to fuse with a sacral rib16. The incomplete fusion of these two skeletal elements is visible in amphibians (Fig. 4), in which the sacral rib and ilium are joined by cartilage.

Computed microtomography scan (voxel size = 7 μm) of salamander Eurycea longicauda (USNM 543595) pelvis (30.9 mm body length minus tail).
Anterolateral view of pelvic girdle. fe, femur (green); il, iliac bone (pink); is, ischial region of puboischiadic plate (tan); pu, pubic region of puboischiadic plate (tan); sr, sacral rib (dark purple).
Walking kinematics
Cryptotora thamicola walked on rough and smooth wet surfaces while out of water. We observed that Cryptotora could walk towards the direction of flowing water (Supplementary Video 3). Hillstream loach relatives of Cryptotora in the family Homalopteridae are known to have epidermal ridges and spines to facilitate adhesion to surfaces in fast-flowing water1,23, but it was not obvious from the μCT scans that these were present on Cryptotora.
During walking, the body midline of Cryptotora (Fig. 5A) moved in a standing wave and did not follow an undulatory, traveling wave pattern as a swimming fish might use. Swimming fish generally undulate in such a way that little lateral displacement occurs near the trunk and the greatest amplitude of undulation occurs posteriorly, in the tail24. All of the fish species that “walk” on land, including mudskippers9, lungfish12, polypterus8, snakeheads, climbing perch25 and catfishes, do so by using their tail to push their bodies forward, pivoting over the pectoral fins. In Cryptotora, there was no undulation of the tail and body bending occurred primarily between the appendicular girdles, as in salamanders26, as the pectoral and pelvic girdles rotate opposite to each other. Cryptotora used a diagonal-couplets lateral sequence gait that was observed both in video taken in the fish’s natural habitat (in flow speeds of approximately 60 cm/s) and in a glass aquarium tilted to 45° and 90° (Fig. 5B, Supplementary Video 1, 3). This symmetric alternating gait is typically the most stable for tetrapods because the center of mass is supported by the triangular orientation of the limbs 19,26,27 and was characterised by semi-synchronous movement of the right forefin and lefthindfin (RF-LH) followed by semi-synchronous movement of the left forefin and right hindfin (LF-RH). There was no observable difference in kinematics between sequences collected at 45˚ and 90° inclinations. Walking motion appeared to be primarily driven by rotation of the appendicular girdles relative to the long axis of the fish rather than by protraction and retraction of individual fins; however, further analysis of higher resolution video is necessary to test this.

Walking kinematics of Cryptotora thamicola climbing a glass surface at a 45 degree angle, ventral view.
(A) Midline spline curve from five consecutive step cycles, head is to the right and aligned to same position among frames. (B) stance phase averaged by proportion of step cycle for right forefin (RF, blue), left forefin (LF, green), right hindfin (RH, orange) and left hindfin (LH, purple). The ends of the bars are calculated as mean ± SE touchdown/liftoff for n = 10 step cycles. Boxes at the ends of bars represent 75% confidence intervals. Time zero was designated as the frame where the LH could be seen pushing away from the glass, which was chosen because this motion was consistently observed to happen in a single frame.
A step cycle (i.e. stride) was defined as the amount of time between the push off of the left hindfin (LH), which from the ventral view could be seen to rotate the pelvis to the right, until the next LH push off. The gait duty factor was the proportion of the step cycle in which a fin was in contact with the substrate. The duty factor of Cryptotora was 65% for each fin with no significant difference among fins (F(3,36) = 0.069, p = 0.976). This aquatic walking duty cycle falls between the aquatic and terrestrial modes of the salamander Dicamptodon tenebrosus, for which the gait pattern duty cycle is approximately 41% while submerged and 77% while walking on a treadmill26,28.
It has been hypothesized that early tetrapods that could not lift their bodies off the ground would use a crutching29 or walking trot gait, in which the body moved with an undulatory wave and those that could support their weight would perform a lateral sequence walk, resulting in a standing wave of the axial body19. Cryptotora performs a tetrapodal diagonal-couplets lateral sequence walk with a standing wave but it’s body remains close to the ground, which may be hydrodynamically important when walking in high velocity flow conditions. In reality, the behaviour and morphology of Cryptotora are unique as compared to both swimming fishes and walking salamanders and represent a locomotor strategy that may be heavily constrained by their cave environment.
Evolution of walking in fishes
Tetrapod locomotion began in the water, where the first limbs were used as paddles30. A hallmark of the evolution of tetrapods is the regionalisation of vertebral column articulations between neural centra and with the pelvic girdle for support out of the buoyant fluid environment19. Indeed, the evolution of the pelvic girdle structure points to development on land21; where it is absolutely essential to have a robust mechanical connection between the axial skeleton and the ground to generate upright walking forces30. In fishes, both living and extinct, the body is supported by the surrounding fluid; pelvic fins are used primarily for stabilization against roll31.
A number of researchers have shown that it is possible to use the biomechanical knowledge of extant vertebrates to attempt to better understand the functional constraints that shaped the origin of tetrapod limbs32,33,34,35,36. Even though fish bone is anosteocytic, it has been shown to remodel and reinforce skeletal structures in response to mechanical stress37. This process is known to cause the formation of intermuscular bones in fishes15 and to increase pectoral girdle shape and thickness in polypterus, which walks on land using its pectoral fins8. Thus, the enlarged neuropophyses, ribs and pelvic girdle of Cryptotora may have evolved as a result of mechanical stressors from climbing and walking. Developmental plasticity plays an important role in the appearance of complex heritable phenotypic traits38,39 and extreme environments, like those in caves, are catalysts for the evolution of novel traits. Standen et al. hypothesized that environmentally induced phenotypic plasticity may facilitate macroevolutionary change8; however, our current data does not allow us discriminate between selection for a robust pelvic girdle specifically or for the plastic response to extreme environmental conditions.
Interestingly, the fin shape and footfalls of walking Cryptotora (Fig. 6A) is similar to the patterns of footprints seen in the oldest known Devonian subaqueous tetrapod trackway (382–358 mya; Fig. 6B) from the Genoa River Beds in New South Wales, Australia40. In both cases the pes (hindfin) print overlies the manus (forefin) on the same side33,40. Also, the six or more digit prints at the edges of the trackway prints match the orientation and position of the distal end of fin rays in walking Cryptotora; it is not unreasonable that fin rays digging into a substrate, especially when the fin ray tips are free of webbing as they are in Cryptotora, would create a pattern like digits. No known limbed early tetrapods made such tracks33,41 but they could have been made by a fish that walked like Cryptotora. Similar trackways are known from the Valencia Slate Formation in Ireland and these sediments have an earlier date than the presumed origin of tetrapods as indicated by body fossil data19. Notably, many Homalopterid fishes are able to push their bodies off the substrate and stand perched on their fins1; however, we predict that if Cryptotora were to walk on a soft substrate (which is non-native to its cave environment), it would likely leave a body and tail imprint, like that seen with some of the tracks in the Genoa River Bed40.

Finprint comparison.
(A) Tracing of Cryptotora fins from ventral view during walking sequence on glass at 45° angle. Final LF (green) fin tracing is shown with fin ray orientations drawn. (B) footprint outlines of earliest known tetrapod trackway from the Devonian, redrawn from Warren and Wakefield40, rotated to match walking direction of Cryptotora (towards top of page) and scaled to similar size (scale bars are 50 mm, bottom).
It is, however, crucial to note that Cryptotora is not an analogous representative of any early tetrapodamorph described to date. Description of the morphological attributes of early tetrapods that were instrumental in the fin to limb transition included the presence of zygapophyses and large neural spines on vertebrae and a pelvic girdle that was firlmly attached to the axial skeleton; Cryptotora convergently evolved these while no other fish has. However, Cryptotora obviously lacks digited appendages, which evolved before the pelvic girdle in the fin-to-limb transition.
In the Devonian period, vertebrates gained novel morphological and behavioural traits that facilitated life in a terrestrial environment. Among these, solid-substrate based locomotion, alternating gait by pelvic propulsors and digit-bearing limbs are considered to be critical adaptations for the emergence of tetrapods12,33,42. Recent works on terrestrial locomotion in fishes have shown that pelvic appendage driven locomotion on a hard surface was possible before the evolution of digited limbs12. Our work supports those findings and goes on to show that fish are capable of evolving a robust pelvic girdle that is firmly attached to the vertebral column in the absence of digited limbs; this finding is especially worth considering given the fossils trackways showing tetrapodal locomotion that predate the origin of digited limbs. While there have been multiple cases of secondarily aquatic vertebrates, we only have fossil evidence of one period of time in which vertebrates emerged from an aquatic lifestyle and evolved terrestrial walking behaviour19. Future studies of extant fishes that convergently evolved morphological and behavioural features typical of tetrapods will offer a window into understanding the biomechanical constraints that enhanced selection on this complex transformation.
Experimental Procedures
Morphological analysis
A paratype specimen of Cryptotora thamicola (NIFI 03046, Tham Maelana, Ban Maelana, Tumbon Pangmapha, Pangmapha district, Mae Hong Son Province, Thailand; 47.05 mm total length, TL) was scanned in a Skyscan 1173 computed microtomography (μCT) scanner located at the Advanced Dental Technology Center in Thailand Science Park, Patumthanee Province, Thailand. The scan was performed at 8.8 μm voxel (50 kV, 160 uA) resolution43. One common goldfish (Carassius auratus; 38 mm TL) and one salamander (Eurycea longicauda, USNM 543595; 30.9 mm body length excluding tail) were scanned in a Skyscan 1173 μCT scanner at the molecular imaging center at Rutgers University, New Brunswick, NJ at 6.9 μm voxel resolution. Three-dimensional reconstruction of the μCT scans was performed in Mimics 18 (Materialise USA, Plymouth, MI). Because collection of live Cryptotora specimens and dissection of museum specimens is prohibited, all morphological details for this species were extracted through analysis of the high resolution μCT scan. No manipulation of the Cryptotora specimen was permitted by the Department of Fisheries in Thailand, including staining with contrast solutions that would allow visualisation of soft tissues by μCT scanning.
Kinematic analysis
Cryptotora thamicola were observed in situ at Mae Lana cave, Ban Maelana, Tumbon Pangmapha, Pangmapha district, Mae Hong Son Province, Thailand. Animals are restricted from being removed from the cave and all kinematic video data were obtained at the site the fish were found. Two individuals were carefully scooped into a glass tank for kinematic sequence recording; fourteen walking sequences of 4–9 steps in a straight line at 45° (n = 8) and 90˚ angles (n = 6) were recorded using a Sony HDR-PJ340E camera at 25 fps, which captured 8–10 frames per step cycle. The low sample size in this study reflects the limited access to specimens and therefore the descriptive kinematics of the walking behaviour presented here should not be treated as a comprehensive kinematic analysis. Fin kinematics were digitised using the DLTdv5 MATLAB 2014a (Mathworks, USA) program developed by T. Hedrick44. Cavefish selected for study were released unharmed after approximately 15 minutes of filming. All animals were handled ethically according to institutional animal care and use protocols approved by the New Jersey Intitute of Technology (14–036).
Additional Information
How to cite this article: Flammang, B. E. et al. Tetrapod-like pelvic girdle in a walking cavefish. Sci. Rep. 6, 23711; doi: 10.1038/srep23711 (2016).
References
Lindsey, C. Form, Function and Locomotory Habits in Fish. Fish Physiol. 7, 1–100 (1979).
Gibb, A. C., Ashley-Ross, M. A. & Hsieh, S. T. Thrash, flip, or jump: the behavioral and functional continuum of terrestrial locomotion in teleost fishes. Integr. Comp. Biol. 53, 295–306 (2013).
Soares, D. & Bierman, H. S. Aerial jumping in the Trinidadian guppy (Poecilia reticulata). PloS ONE 8, e61617 (2013).
Gillis, G. B. Environmental effects on undulatory locomotion in the American eel Anguilla rostrata: kinematics in water and on land. J. Exp. Biol. 201, 949–961 (1998).
Gibb, A. C., Ashley-Ross, M. A., Pace, C. M. & Long, J. H. Fish out of water: terrestrial jumping by fully aquatic fishes. J. Exper. Zool. Part A 315, 649–653 (2011).
Clardy, T. R. Aquatic and terrestrial locomotion of the rock prickleback, Xiphister mucosus (Cottiformes: Zoarcoidei: Stichaeidae). Northwest. Naturalist 93, 203–210 (2012).
Edwards, J. L. Two perspectives on the evolution of the tetrapod limb. Amer. Zool. 29, 235–254 (1989).
Standen, E. M., Du, T. Y. & Larsson, H. C. Developmental plasticity and the origin of tetrapods. Nature 513, 54–58 (2014).
Kawano, S. M. & Blob, R. W. Propulsive forces of mudskipper fins and salamander limbs during terrestrial locomotion: implications for the invasion of land. Integr. Comp. Biol. ict051 (2013).
Pace, C. & Gibb, A. Mudskipper pectoral fin kinematics in aquatic and terrestrial environments. J. Exp. Biol. 212, 2279–2286 (2009).
Johnels, A. The mode of terrestrial locomotion in Clarias. Oikos 8, 122–129 (1967).
King, H. M., Shubin, N. H., Coates, M. I. & Hale, M. E. Behavioral evidence for the evolution of walking and bounding before terrestriality in sarcopterygian fishes. Proc. Nat. Acad. Sci. USA 108, 21146–21151 (2011).
Blob, R., Rai, R., Julius, M. & Schoenfuss, H. Functional diversity in extreme environments: effects of locomotor style and substrate texture on the waterfall‐climbing performance of Hawaiian gobiid fishes. J. Zool. 268, 315–324 (2006).
Trajano, E. et al. Habitat, distribution, ecology and behavior of cave balitorids from Thailand (Teleostei: Cypriniformes). Ichthyol. Explor. Freshwaters 13, 169–184 (2002).
Hall, B. K. Fins into limbs: evolution, development and transformation. (University of Chicago Press, 2008).
Boisvert, C. A., Joss, J. M. & Ahlberg, P. E. Comparative pelvic development of the axolotl (Ambystoma mexicanum) and the Australian lungfish (Neoceratodus forsteri): conservation and innovation across the fish-tetrapod transition. EvoDevo 4, 3 (2013).
Boisvert, C. A. The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature 438, 1145–1147 (2005).
Danos, N. & Staab, K. Can mechanical forces be responsible for novel bone development and evolution in fishes? J. Applied Ichthyol. 26, 156–161 (2010).
Clack, J. A. Gaining ground: the origin and evolution of tetrapods. (Indiana University Press, 2012).
Pierce, S. E. et al. Vertebral architecture in the earliest stem tetrapods. Nature 494, 226–229 (2013).
Carroll, R. L. & Holmes, R. B. In Fins into Limbs: Evolution, Development and Transformation (ed Brian K. Hall ) (Chicago: Univ. Chicago Press, 2008).
Harris, V. A. On the locomotion of the mud-skipper Periopthalmus koelreuteri (Pallus) (Gobiidae) Proc. Zool. Soc. Lond. 134, 107–135 (1960).
Das, D. & Nag, T. C. Adhesion by paired pectoral and pelvic fins in a mountain-stream catfish, Pseudocheneis sulcatus (Sisoridae). Environ. Biol. Fishes 71, 1–5 (2004).
Lauder, G. V. & Tytell, E. D. In Fish Physiology Vol. Fish Biomechanics: 23 (eds R. E. Shadwick & G. V. Lauder ) 425 (Academic Press, 2006).
Davenport, J. & Matin, A. Terrestrial locomotion in the climbing perch, Anabas testudineus (Bloch)(Anabantidea, Pisces). J. Fish Biol. 37, 175–184 (1990).
Ashley-Ross, M. A., Lundin, R. & Johnson, K. L. Kinematics of level terrestrial and underwater walking in the California newt. Taricha torosa. J. Exper. Zool. A 311, 240–257 (2009).
Hildebrand, M. Analysis ofthe symmetrical gaits of tetrapods. Fol. Biotheor. 6, 9–22 (1966).
Ashley-Ross, M. Hindlimb kinematics during terrestrial locomotion in a salamander (Dicamptodon tenebrosus). J. Exp. Biol. 193, 255–283 (1994).
Pierce, S. E., Hutchinson, J. R. & Clack, J. A. Historical perspectives on the evolution of tetrapodomorph movement. Integr. Comp. Biol. 53, 209–223 (2013).
Lebedev, O. A. Palaeontology: Fins made for walking. Nature 390, 21–22 (1997).
Standen, E. M. Muscle activity and hydrodynamic function of pelvic fins in trout (Oncorhynchus mykiss). J. Exp. Biol. 213, 831–841 (2010).
Pierce, S. E., Clack, J. A. & Hutchinson, J. R. Three-dimensional limb joint mobility in the early tetrapod Ichthyostega. Nature 486, 523–526 (2012).
Clack, J. A. The fin to limb transition: new data, interpretations and hypotheses from paleontology and developmental biology. Ann. Rev. Earth Planet. Sci. 37, 163–179 (2009).
Bishop, P. J. et al. Oldest pathology in a tetrapod bone illuminates the origin of terrestrial vertebrates. PLoS One 10, doi: 10.1371/journal.pone.0125723 (2015).
Hutchinson, J. R. On the inference of function from structure using biomechanical modelling and simulation of extinct organisms. Biol. Letters 8, 115–118 (2011).
Hutchinson, J. R. & Gatesy, S. M. Dinosaur locomotion: beyond the bones. Nature 440, 292–294 (2006).
Atkins, A., Milgram, J., Weiner, S. & Shahar, R. The response of anosteocytic bone to controlled loading. J. Exp. Biol. 218, 3559–3569 (2015).
Wund, M. A., Baker, J. A., Clancy, B., Golub, J. L. & Foster, S. A. A test of the “flexible stem” model of evolution: ancestral plasticity, genetic accommodation and morphological divergence in the threespine stickleback radiation. Amer. Natur. 172, 449–462 (2008).
Badyaev, A. V. Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation. Proc. Roy. Soc. London B 272, 877–886 (2005).
Warren, J. W. & Wakefield, N. A. Trackways of tetrapod vertebrates from the Upper Devonian of Victoria, Australia. Nature 238, 469–470 (1972).
Clack, J. A. Devonian tetrapod trackways and trackmakers; a review of the fossils and footprints. Palaeogeo. Palaeoclim. Palaeoecol. 130, 227–250 (1997).
Shubin, N. H., Daeschler, E. B. & Jenkins, F. A. Pelvic girdle and fin of Tiktaalik roseae. Proc. Nat. Acad. Sci. USA 111, 893–899 (2014).
Flammang, B. E. Pelvic girdle of Cryptotora thamicola. Open Science Framework. Data archive. Available at: http://osf.io/ahswb (2016). Date of access 01/02/2016.
Hedrick, T. L. Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinsp. Biomim. 3, 034001–034006 (2008).
Acknowledgements
J.R. Hutchinson, M. Ashley-Ross, E. Fortune and G. Haspel provided invaluable comments on early drafts of this manuscript. Thanks to P. Polkhan (Fisheries Department of Thailand); R. Wattanasiriserekul, T. Chamnivikaipong and A. Kampanphet (Faculty of Fisheries Technology and Aquatic Resources); and Maelana villagers who helped in the field trips. S. Suksri provided the specimen from the National Inland Fisheries Institute for μCT scanning and the technician from the Advanced Dental Technology Center. Discussion with S.B. Crofts was invaluable for parsing out functional morphology. B.E.F. is indebted to and inspired by F.A. Jenkins and K.F. Liem for their extraordinary mentorship, lasting morphological insights and ironic conversations about fish pelvic girdles. D.S. is indebted to R. Borowski for insights into Cryptotora biology and to Y. Zhao, who introduced her to A.S.
Author information
Authors and Affiliations
Contributions
The project was conceived by B.E.F. and D.S.; data were collected by D.S. and A.S.; B.E.F. and J.M. processed CT scans; B.E.F. analysed and interpreted the data and drafted the initial manuscript; D.S. and A.S. contributed to revision of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Flammang, B., Suvarnaraksha, A., Markiewicz, J. et al. Tetrapod-like pelvic girdle in a walking cavefish. Sci Rep 6, 23711 (2016). https://doi.org/10.1038/srep23711
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep23711
This article is cited by
-
An adhesive locomotion model for the rock-climbing fish, Beaufortia kweichowensis
Scientific Reports (2019)
-
Extra-Visual Systems in the Spatial Reorientation of Cavefish
Scientific Reports (2018)
-
Trackways Produced by Lungfish During Terrestrial Locomotion
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
