
Abstract
Plant beneficial microbes mediate biocontrol of diseases by interfering with pathogens or via strengthening the host. Although phytohormones, including cytokinins, are known to regulate plant development and physiology as well as plant immunity, their production by microorganisms has not been considered as a biocontrol mechanism. Here we identify the ability of Pseudomonas fluorescens G20-18 to efficiently control P. syringae infection in Arabidopsis, allowing maintenance of tissue integrity and ultimately biomass yield. Microbial cytokinin production was identified as a key determinant for this biocontrol effect on the hemibiotrophic bacterial pathogen. While cytokinin-deficient loss-of-function mutants of G20-18 exhibit impaired biocontrol, functional complementation with cytokinin biosynthetic genes restores cytokinin-mediated biocontrol, which is correlated with differential cytokinin levels in planta. Arabidopsis mutant analyses revealed the necessity of functional plant cytokinin perception and salicylic acid-dependent defence signalling for this biocontrol mechanism. These results demonstrate microbial cytokinin production as a novel microbe-based, hormone-mediated concept of biocontrol. This mechanism provides a basis to potentially develop novel, integrated plant protection strategies combining promotion of growth, a favourable physiological status and activation of fine-tuned direct defence and abiotic stress resilience.
Similar content being viewed by others


Introduction
Throughout their life cycle, plants interact with a multitude of environmental factors, including unfavourable abiotic stress conditions and threats from a wide range of insects and pathogenic microbes. Phytohormone signalling plays a crucial role in accurately regulating plant responses. Ethylene (ET), jasmonic (JA) and salicylic acid (SA) are essential phytohormonal regulators of plant immunity that form a central signalling backbone which specifically coordinates defence responses against biotrophic and necrotrophic pathogens1. Detailed analyses of phytohormone function in plant immunity have extended this network to other classic growth-regulating phytohormones such as abscisic acid (ABA), auxins and gibberellins2,3,4. The classic growth-stimulating phytohormone family of cytokinins (CKs) comprises important regulators of many physiological and developmental plant processes such as cell division, leaf senescence, nutrient mobilization, apical dominance and seed germination5,6. In the interaction of plants with insects and microbes, CK alterations have been identified to cause green island formation, galls, growth abnormalities7 and modulation of primary carbon metabolism8. As they induce sink metabolism7,9, CKs have been suggested to alter host physiology to facilitate maximum access of (hemi)biotrophic pathogens to nutrients during early interactions7. However, recently, significant direct functions for CKs in plant immunity have been identified in different plant species such as Arabidopsis thaliana10,11, tobacco12 and rice13 via induction of resistance against primarily (hemi)biotrophic pathogens such as Pseudomonas syringae and Hyaloperonospora arabidopsidis or by activation of defence responses (independent of pathogen infection). The underlying mechanisms mediating CK-dependent resistance against P. syringae include induction of SA in Arabidopsis and tobacco10,12, induction of phytoalexin accumulation12,14 and reduction of ABA levels in tobacco15. Furthermore, CKs were demonstrated to induce defence gene expression synergistically with SA13 and to enhance diterpenoid phytoalexin accumulation16 in rice.
In addition to pathogens, plants interact with a multitude of beneficial microbes, many of which belong to the genera Azospirillum, Bacillus or Pseudomonas and are characterized by their ability to promote plant growth, increase tolerance to environmental stress and/or enhance disease resistance. Agricultural food production faces many challenges due to increasing world population, climate change and restrictions on use of classic pesticides. Consequently, alternative plant protection strategies are urgently required. The biological control of plant diseases by beneficial microbes offers significant potential for integrated plant disease management17. To facilitate the development of microbe-based biocontrol strategies, their underlying mechanisms have to be fully elucidated. Known biocontrol mechanisms include (i) direct interference with the pathogen, such as competition for nutrients and space, secretion of antibiotics or degradation of virulence factors and (ii) the induction of host plant resistance, which is often related to induced systemic resistance (ISR) involving the phytohormones ET and JA18,19,20. Interestingly, beneficial microbes are capable of producing different phytohormones, notably including CKs. Therefore, it is intriguing that CKs exhibit similar biological effects as described for beneficial microbes including the induction of plant growth promotion (PGP), environmental stress tolerance and disease resistance. Despite this correlative evidence, microbial phytohormones - and particularly CKs - have not been considered as a determinant for effective biocontrol of plant diseases. Microbial CK production has so far only been linked to PGP21,22 and suggested as a mechanism for increasing abiotic stress tolerance in plants23. Considering the widespread CK production by beneficial microbes and recent advances in understanding CK function in plant resistance, we analysed the impact of microbial CK production on the microbe’s biocontrol ability. We established a causal relationship between the production of CKs by Pseudomonas fluorescens (Pfl) strain G20-1824,25 and its ability to control the infection of Arabidopsis by P. syringae pv. tomato DC3000 (Pto) through comparisons with G20-18-derived loss-of-function and gain-of-function strains in a leaf infiltration assay. Analyses of Arabidopsis mutant lines impaired in defence or hormone signalling pathways revealed the necessity of functional CK perception in combination with SA defence signalling and a potential minor impact of ET, JA signalling as well as camalexin accumulation to fully establish microbial CK-mediated biocontrol. These data provide the basis for a novel microbe-based concept of biocontrol.
Results
Microbial CKs mediate G20-18 biocontrol
Since the CK-producing PGP Pfl strain G20-18 had not been tested for its biocontrol abilities, we first examined its biocontrol potential in the Arabidopsis–Pto pathosystem26 in comparison to its CK-deficient transposon mutants CNT1 and CNT224,25. As CKs have been demonstrated to induce defence responses or resistance against (hemi)biotrophic foliar pathogens when applied to leaves of Arabidopsis10,11,27, rice13,16 and tobacco12,28, we decided to analyse the biocontrol potential of the Pfl strains when directly applied to Arabidopsis leaves by infiltration of cell suspensions 48 h prior to Pto infection. The leaf infiltration assay widely used in model pathosystems was chosen to allow us to relate the findings to the well-established immunity-relevant CK functions in leaf tissues. Although approaches such as spray inoculation or application to the root system would address more natural scenarios of interaction, they would contribute additional sources of interference with CK-mediated immunity responses and thus, further complicate the analyses of a potential role of CK in biocontrol.
Pre-treatment with Pfl G20-18 heavily suppressed Pto symptom development at 4 days post infection (dpi), resulting in maintenance of tissue integrity, an important beneficial aspect of biocontrol applications in sustaining biomass yield. Mock pre-treatment had no effect on Pto symptoms compared to control infections without pre-treatment (Fig. 1a). Thus, G20-18 is considered an efficient strain for biocontrol of Pto in Arabidopsis in the leaf infiltration assays. In comparison to G20-18, both CNT transposon mutants had only a slight suppressive effect on Pto symptom development (Fig. 1a). The quantification of the average symptom scores over all experiments further demonstrates this biocontrol effect: G20-18 pre-treatment efficiently suppressed Pto symptoms by approximately 75%, CNT pre-treatments suppressed Pto symptoms only by 15 to 20% compared to untreated and mock controls, indicating that the CK-deficient mutants were significantly less effective than G20-18 (Fig. 1b). This highly reduced effect of the CK-deficient CNT transposon mutants on Pto symptom development strongly supports a role for microbial CK production in the biocontrol ability of G20-18.

Pfl G20-18 suppresses Pto symptoms in Arabidopsis.
(a) Pto symptom development in Arabidopsis leaves (right halves) 4 days post infection (dpi) with 106 cfu ml−1 is strongly suppressed by G20-18 compared to controls and CNT pre-treatments. (b) Average Pto symptom score in Arabidopsis 4 dpi with 106 cfu ml−1 is significantly lower after G20-18 pre-treatment compared to controls and CNT pre-treatments. Data are means ± s.e. n ≥ 300, letters indicate different significance groups (P < 0.05).
As the CNT transposon mutants were generated by undirected mutagenesis via the introduction of the TnphoA transposon into G20-18 and were selected based on CK deficiency without detailed genetic characterization24, we analysed the only known CK biosynthetic gene in Pfl strains, tRNA delta(2)-isopentenylpyrophosphate transferase (miaA). Using primers based on known Pfl miaA sequences the gene was amplified from G20-18 and sequenced (Supplementary Fig. 1). Size comparison of full-length miaA amplicons of G20-18 and the CNT transposon mutants as well as sequence analysis ruled out miaA as the direct target of TnphoA. Subsequent semi-quantitative RT-PCR analysis revealed that miaA transcript levels in the CNT transposon mutants were strongly reduced by approximately 50% compared to G20-18 (Supplementary Fig. 2). This suggests that regulatory components in the CNT mutants were affected by the transposon mutagenesis, potentially interfering with miaA transcription or the processing and stability of miaA transcripts. Since the mechanism of transcriptional regulation of miaA is not elucidated, we used directed functional approaches to further substantiate the link between miaA as a determinant of microbial CK production and subsequent biocontrol activity against Pto.
Considering the reduced miaA transcript levels in the CNT transposon mutants, compared to G20-18, as the cause for the difference in biocontrol efficacy, functional complementation for CK production by the CNT transposon mutants (gain-of-function) was performed to assess the possible restoration of their biocontrol ability. Therefore, the CK biosynthetic genes isopentenyltransferase from Agrobacterium tumefaciens (ipt) for heterologous expression and the endogenous Pfl G20-18 miaA for homologous expression were fused to a lac-promoter in the expression vector pBBR1MCS-5. The different Pfl strains were transformed with these gain-of-function constructs or the empty vector (EV) and analysed for their biocontrol activities. The presence of the EV did not affect biocontrol activity of G20-18 as this strain efficiently restricted Pto symptom development (Fig. 2a) comparable to G20-18 wild-type (Fig. 2b). Also in the CNT transposon mutants, the EV did not cause changes (Fig. 2a) as symptoms were still significantly less suppressed compared to G20-18 (Fig. 2b). In contrast, the ipt- or miaA-complemented CNT transposon mutants, exhibited restored biocontrol activities as evidenced by a strong suppression of Pto symptom development (Fig. 2a), comparable to G20-18 biocontrol activity (Fig. 2b). This wild-type-like biocontrol activity in the two CNT transposon mutants functionally complemented via restored CK production by two different CK biosynthetic genes supports the role of microbial CKs as a key determinant for efficient biocontrol of Pto.

Complementation of the CNT transposon mutants with a functional CK biosynthetic gene restores their biocontrol ability.
(a) The biocontrol ability of CNT transposon mutants is restored by complementation with functional Atipt or G20-18miaA evident from strongly reduced Pto symptoms (right leaf halves) 4 days post infection (dpi) with 106 cfu ml−1. Transformation with the empty vector pBBRMCS-5 (EV) has no effect. (b) Average Pto symptom score in Arabidopsis 4 dpi with 106 cfu ml−1 after indicated pre-treatments. Data are means ± s.e. n ≥ 226, letters indicate different significance groups (P < 0.05).
To substantiate the gain-of-function data, a complementary loss-of-function approach was followed, addressing the function of miaA and subsequent CK production in G20-18-mediated biocontrol of Pto. To this end, the impact of directed knockout of the G20-18 miaA gene by insertion of a kanamycin resistance cassette into the miaA coding region on the biocontrol ability was assessed. This resulted in the Pfl knockout mutant ΔmiaA, which tested PCR-positive for the integration of the disrupted miaA gene sequence in its genome. RT-PCR confirmed the lack of miaA transcripts and thus the functional knockout in this strain (Supplementary Fig. 2). Assays with this ΔmiaA knockout mutant revealed a significant reduction in biocontrol compared to G20-18 wild-type as illustrated by stronger Pto symptom development (Fig. 3). Together, the gain-of-function and directed loss-of-function approaches prove the importance of microbial CK production for their biocontrol ability in the leaf infiltration assays. Interestingly, the distinct functional miaA knockout in ΔmiaA (Supplementary Fig. 2) did not further reduce the biocontrol ability compared to the transposon mutants CNT1 and 2 (Fig. 3) in which low levels of miaA transcripts were still detectable (Supplementary Fig. 2). This suggests that the described biocontrol effect depends on minimum threshold levels of miaA transcripts which subsequently determine CK levels that suffice to induce resistance under particular conditions.

Distinct ∆miaA knockout in Pfl G20-18 exhibits a reduced biocontrol activity.
(a) ΔmiaA loss-of-function mutant is impaired in its biocontrol ability indicated by stronger Pto symptom development (right leaf halves) 4 days post infection (dpi) with 106 cfu ml−1 compared to Pfl G20-18 pre-treatment. (b) Average Pto symptom score in Arabidopsis 4 dpi with 106 cfu ml−1 after indicated pre-treatments. Data are means ± s.e. n ≥ 79, letters indicate different significance groups (P < 0.05).
G20-18 biocontrol affects CKs in planta
Based on the established link between Pfl G20-18 CK production and its biocontrol abilities described above, the in planta CK levels were analysed as these should ultimately reflect their contribution to the induction of resistance or defence responses10,11,12,13,14,15,27. Therefore, we analysed the accumulation of 25 individual CK species comprising the free nucleobases as well as conjugates29 in pooled samples of whole Arabidopsis leaves 48 h post infiltration with the different Pfl strains, which corresponds to the time-point of Pto infection. Thus, these samples integrate all processes related to each individual pre-treatment and determine the plant tissue status at the critical time-point of infection that defines the outcome of the plant-pathogen interaction. CKs were analysed in two sample sets, one comparing the pre-treatments with G20-18, the miaA- or ipt-complemented CNT transposon mutants and mock control (Table 1 and Supplementary Table 1) and the second comparing pre-treatments with G20-18, the CNT transposon mutants, the ΔmiaA knockout mutant and mock control (Table 1 and Supplementary Table 2). Eight of ten CK levels that increased after G20-18 treatment in the first set (Supplementary Table 1) also increased in the second set (Supplementary Table 2). A clear trend of lower CK levels in plant tissue pre-treated with loss-of-function CNT transposon or ΔmiaA knockout mutants was observed compared to G20-18 (ratios of 0.82 to 0.89). In contrast, this effect was reversed in tissue treated with the functionally complemented CNT transposon mutants that showed even higher CK levels compared to G20-18 (ratios of 1.08 to 1.11, Table 1). Since CK types differ in their biological activity and signalling function, the individual consideration of specific CK species is important. Total tZ-, cZ-, DHZ- and iP-type CK levels showed similar trends as total CK levels with lower levels after treatments with the CK-deficient mutants (Supplementary Table 2) and reversion in the functionally complemented CNT transposon mutants (Supplementary Table 1), which correlates with their differential effect on Pto symptom development (Fig. 1, 2, 3). Similarly, levels of the free nucleobases as the most active CK species5 were lower in tissue treated with CK-deficient mutants (ratios of 0.63 to 0.97) and higher in tissue treated with the functionally complemented CNT transposon mutants (ratios of 1.34 to 2.38) compared to G20-18 (Table 1). In particular, the individual nucleobases tZ, cZ and iP accumulated to higher levels after treatment with the functionally complemented CNT transposon mutants compared to G20-18 (ratios of 1.30 to 2.50), while these nucleobases in general showed lower accumulation after mutant treatments (Table 1). The most prominent differences were detected in the accumulation of the highly active tZ. Treatment with the three CK-deficient mutants caused significantly lower tZ levels (ratios of 0.47 to 0.60) than G20-18 treatment (Table 1), which can directly be related to the defects in miaA expression as miaA has been identified to be responsible for the specific formation of tZ and derivatives from tRNA in different bacteria30,31,32. Intriguingly, exogenously supplied tZ efficiently restricted infections of Arabidopsis with Pto10 and tobacco with P. syringae pv. tabaci (Pst)12,28, while cZ had a much weaker effect on the resistance against Pst28 and iP treatment did not increase the resistance of rice against Magnaporthe oryzae13. This could explain why increased cZ or iP levels in some samples after treatment with the CK-deficient mutants had no effect on resistance against Pto and emphasizes the role of tZ levels as a key parameter in G20-18-mediated biocontrol.
The analyses of a large set of individual CK levels revealed subtle though distinct changes in the host plant. In both datasets specific differences between G20-18 and its derivatives were successfully monitored and appeared to be robust even against variable background levels indicated by the variable CK levels in the control samples (Supplementary Table 1 and Supplementary Table 2). The complex regulation of CK levels in Arabidopsis depends on nine biosynthesis and seven catabolism genes that are potentially affected by Pfl, similar to other Arabidopsis-microbe interactions (eFP browser33), which could be responsible for the different CK ratios between G20-18-treated and control samples in the two sets in addition to microbial CK production (Supplementary Table 1 and Supplementary Table 2). In addition, the infiltration process, ambient conditions, inter-conversions and transport of CKs contribute to complex spatiotemporal dynamics at the cellular level, which are difficult to resolve by CK determination in plant tissue. Considering the known activity of CKs at low concentrations, the subtle differences caused by bacterial CK production linked to the CK-mediated plant defence ensures minimal interference with general plant CK homeostasis and thus minimal perturbation of other plant processes.
G20-18 biocontrol limits pathogen growth
The differential efficacy in biocontrol by G20-18 and its CK-deficient mutants could result from growth variations of the Pfl strains in planta, since the number of living Pfl cells may determine biocontrol by competition with Pto for nutrients and space34,35. Such growth differences could depend on their capacity to produce CKs, which may interfere with bacterial quorum sensing36 or the communication between microbe and plant for successful niche establishment37. Additionally, growth defects could be caused by pleiotropic effects of the applied mutagenesis unrelated to the CK deficiency, while reduced viability of the Pfl mutants could cause lower CK production. Therefore, the number of viable Pfl cells in planta at the time-point of Pto infection - 48 hours post infiltration (hpi) of Pfl -, was determined for G20-18 and the different mutant strains (Fig. 4a). Similar numbers of viable cells were determined directly after the infiltration (0 hpi) and at 48 hpi for all strains except CNT2 for which viable cells decreased (significantly compared to CNT1). Based on these data, growth differences between G20-18 and the analysed mutant strains can be excluded as the cause of the variations in their biocontrol abilities.

Pfl G20-18 and its CK-deficient mutants do not differ in growth, but differentially affect Pto proliferation in planta.
(a) Number of viable Pfl cells in Arabidopsis leaves 0 hours post infiltration (hpi) with 107 cfu ml−1 and at the time-point of Pto infection (48 hpi). n = 27. (b) Number of viable Pto cells harbouring pMP4662 in Arabidopsis leaves directly after infiltration (0 hpi) with 105 cfu ml−1 and at 72 hpi. n = 18. Data are means ± s.e., letters indicate different significance groups (P < 0.05).
As CKs can directly contribute to a favourable physiological status by modulating primary metabolism8,9 and thus potentially affect tissue integrity, suppression of symptom development during CK-mediated resistance does not necessarily correlate with restriction of pathogen growth28, which is a direct result of increased resistance. To discriminate between increased resistance induced by G20-18-derived CKs and general impact on tissue integrity, we determined Pto growth in planta after pre-treatment with G20-18 and its CK-deficient mutants. Pto proliferation was significantly reduced after G20-18 pre-treatment compared to the mutant and mock pre-treatments at 72 hpi (Fig. 4b) and thus restricted Pto proliferation can be considered as the cause for reduced symptom development in the leaf infiltration assays. Further, Pto proliferation was strongly negatively correlated with the tZ levels determined at the time-point of infection (Table 1) following pre-treatments with the different Pfl strains (ranked data, Spearman’s correlation coefficient of −0.8). G20-18 pre-treatment resulted in the lowest Pto proliferation and the highest tZ levels, followed by pre-treatments with the CNT transposon mutants which similarly caused lower tZ levels and higher Pto proliferation comparable to mock treatment, while ΔmiaA pre-treatment resulted in the lowest tZ levels and the highest Pto proliferation. This correlation supports the role of specific active CKs in determining biocontrol activities by inducing defence responses that act directly on the pathogen in a dose-dependent manner, similar to resistance effects induced by exogenously applied CKs11,12, which in a certain range can act in a dose-dependent manner and require specific threshold levels to be active.
G20-18 biocontrol requires plant pathways
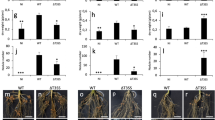
Pfl G20-18 showed suppressive effects on Pto symptom development and multiplication in Arabidopsis indicating direct activation of plant defences, which were lacking after pre-treatment with CK-deficient Pfl mutants. To dissect the underlying plant mechanisms, the efficiency of G20-18-mediated biocontrol was determined in several Arabidopsis lines impaired in phytohormone and/or defence-related mechanisms (Fig. 5a). Since we identified microbial CK production as a determinant of Pfl G20-18-mediated biocontrol against Pto, we assumed functional CK perception as the initial step of CK signalling in the plant to be essential. In Arabidopsis CK perception depends on the three membrane-bound histidine kinases AHK2, AHK3 and AHK4/CRE1/WOL6. The function of these Arabidopsis CK receptors in G20-18 biocontrol was assessed in the double mutant lines ahk2-2/ahk3-3 and cre1-12/ahk3-3 and the triple mutant cre1-12/ahk2-2/ahk3-3(+/−) (homozygous for cre1-12 and ahk2-2, heterozygous for ahk3-3)38. G20-18-mediated biocontrol was reduced in all three mutant lines (Fig. 5a), illustrated by significantly elevated Pto symptom development compared to the wild-type Col-0 (Fig. 5b). This indicates that all three receptors function as signalling components of CK-dependent biocontrol by G20-18, which is supported by the finding that the triple mutant exhibited the strongest defect (Fig. 5b). However, a rudimentary G20-18 biocontrol effect is still observed in these mutant plant lines, which is either due to residual CK perception or is unrelated to G20-18 CK production and/or plant CK signalling.

Pfl G20-18 biocontrol depends on functional hormonal and defence pathways of the host.
(a) Pto symptom development (right leaf halves) 4 days post infection (dpi) with 106 cfu ml−1 in indicated Arabidopsis lines after Pfl G20-18 pre-treatment. (b) Average Pto symptom scores 4 dpi with 106 cfu ml−1 in indicated Arabidopsis mutant or transgenic lines (red bars) compared to Col-0 (Col-gl for myc2) wild-type (blue bars) pre-treated with Pfl G20-18 or the appropriate mock. Data are means ± s. e. n ≥ 28, letters indicate different significance groups (P < 0.05).
SA was demonstrated as a key central defence signalling component of CK-mediated resistance, mainly depending on NPR1 signalling, against Pto in Arabidopsis10, but also as a parameter of CK-induced resistance or defence responses in other plant species12,13,28. The role of SA in G20-18-mediated biocontrol was assessed in Arabidopsis lines either overexpressing nahG (35S::nahG), a SA-degrading enzyme from Pseudomonas putida39, or defective in SA biosynthesis (sid2)40 or SA signalling (npr1)41. In agreement with the known SA-dependent tZ-mediated resistance effect in Arabidopsis10, G20-18 pre-treatment was almost completely ineffective in these lines as Pto symptoms were not suppressed (Fig. 5a). 35S::nahG, sid2 and npr1 (Fig. 5b) showed Pto symptoms after G20-18 treatment comparable to the mock treatment in the plant mutants and Col-0 wild-type, hence SA accumulation as well as functional SA signalling have to be considered as major parameters in CK-mediated biocontrol.
To examine involvement of the defence-related phytohormones JA and ET, which are important for inducing ISR as part of biocontrol and for priming effects mediated by beneficial microbes4,20, G20-18 biocontrol assays were performed in the mutant lines myc2 (jin1)42 which is partially insensitive to JA43 and ein244, which is insensitive to ET. In both Arabidopsis lines, the suppressive effect of G20-18 on Pto symptoms was reduced (Fig. 5a). Although this reduction was significant compared to Arabidopsis wild-type Col-gl (myc2) and Col-0 (ein2; Fig. 5b), it is considerably lower than observed in SA-related plant mutants, indicating a minor role of JA and ET in this biocontrol mechanism in leaf infiltration assays. As phytoalexins also potentially contribute to biocontrol effects as antimicrobial compounds45,46, G20-18 biocontrol effects were analysed in Arabidopsis pad347 and cyp7948 mutants deficient in camalexin, the key phytoalexin in Arabidopsis49. In both lines, the effect of G20-18 on Pto was reduced, as evidenced by stronger symptom development compared to Col-0 (Fig. 5a), which was significantly lower, however, than the mock controls (Fig. 5b). These data suggest a minor role of camalexin in G20-18-mediated biocontrol in the leaf infiltration assays, which possibly depends on microbial CKs similar to CK-induced resistance effects shown in tobacco12,14,15.
Discussion
The biological control of plant diseases by beneficial microbes includes well-known mechanisms such as antibiotic production, competition for nutrients and space, or activation of plant defences34,35. Our results identified Pfl G20-18 as a biocontrol strain that efficiently suppressed Pto infection of Arabidopsis when applied to leaves. By dual functional approaches modulating either CK production in the beneficial microbe or CK perception and defence signalling in the host plant, we, to the best of our knowledge, for the first time, identified CK as a microbe-derived phytohormone that functions as a key determinant of microbial biocontrol activity by directly activating plant resistance. This novel concept seems to be independent of classic biocontrol mechanisms such as competition or antibiotic production and strongly depends on functional CK perception as well as SA accumulation and signalling of the host plant (Fig. 5). This is in agreement with known CK-SA interactions in plant immunity10,12,13, particularly considering the strong dependency on SA of CK-induced resistance against Pto in Arabidopsis leaves by exogenously applied tZ10. In contrast, the CK-mediated biocontrol of G20-18 in Arabidopsis seems to depend not or only to a limited extent on JA, ET and camalexin accumulation. The apparent minor effect of these defence signalling pathways could be due to the specific biocontrol assays performed in Arabidopsis leaves and particularly the contribution of JA and ET is probably more relevant in natural microbe-root interactions. Furthermore, these mechanisms do not necessarily depend on microbial CK production and could therefore be responsible for the rudimentary suppression of Pto symptoms caused by the CK-deficient Pfl mutants. Generally, the contribution of all these underlying, networked mechanisms to CK-mediated biocontrol as well as the relevance of microbial CKs for biocontrol per se, can vary depending on the conditions of the interaction between the beneficial microbe and the plant. Based on the presented results, which strongly support the potential of microbial CKs for biocontrol effects, directed analyses of CK function in more complex biocontrol systems such as the natural interaction between beneficial microbes and plant roots or their relevance in practical applications such as spraying or seed coating can be conducted in the future.
In the model biocontrol assays used here G20-18 seems to cause only subtle and fine-tuned, but highly efficient changes in the host plant CK levels. Induction of the subsequent resistance seems to be specifically regulated, probably in concert with additional mechanisms such as direct modulation of the metabolic plant-pathogen interface50 or interference with the niche establishment of Pto37. Since CKs are critically involved in various aspects of plant growth, development and physiology6, such a highly fine-tuned mechanism ensures minimal perturbation of other central plant processes and thus excludes any kind of dramatic impact apparent in transgenic plants with modulated CK homeostasis51,52,53, or known from plant diseases related to microbial CK production, such as tumours caused by Agrobacterium54,55. This aspect of fine-tuned CK modulation is of particular importance in the context of plant immunity, as exogenously supplied CKs (benzyl adenine or tZ-riboside) below certain threshold concentrations were shown to potentially also increase susceptibility in the interaction of Arabidopsis with H. arabidopsidis11 and Pto56 under the specific experimental conditions used in these studies. In the Arabidopsis-Pto interaction the activity of the Pto effector HopQ1 activated CK signalling, had an impact on some CK levels and interfered with immunity56. Our results together with the increasing experimental evidence within other reports on resistance mediated by increased CK levels emphasize that CK modulation of plant immunity has to be fine-tuned to cause beneficial effects for the plant and that the outcome is influenced by additional parameters.
Considering the assumed widespread ability of microbes to produce CKs based on the presence of CK biosynthetic genes, it may be concluded that this novel CK-dependent defence mechanism also contributes to other yet uncharacterized biocontrol systems, potentially including positive effects on environmental stress tolerance as indicated for CKs57 and CK-producing microbes23. Thus, the possibility to achieve abiotic and biotic co-tolerance via microbial CK production is a potential practical application of the results obtained, provided that they are validated in the biocontrol and application systems of interest. The positive effects mediated by beneficial microbes, such as PGP, increased tolerance to environmental stress and pathogen resistance might be integrated by their capacity to produce CK profiles of specific quantity and composition. Thus, screening for microbial phytohormone, particularly CK production must be considered as an additional parameter when characterizing new beneficial plant-microbe interactions or potential biocontrol strains. This finding that microbial CKs determine biocontrol effects could potentially offer novel options for developing alternative strategies for integrated plant protection combined with PGP. Both effects contribute to improved plant productivity, which is needed to cope with challenges such as increasing world population, climate change and restricted use of classic pesticides. Optimized CK production by beneficial microbes could be an excellent biological alternative to classic pesticides and fertilizers and may work efficiently in different (crop) plant species since CK-dependent resistance mechanisms have been identified in a variety of plants.
Methods
Plant lines and growth conditions
Arabidopsis plants were grown in soil under controlled short day (8/16 h, 22/20 °C day/night) conditions at 60% relative humidity in growth chambers (APT.line™ KBW 720, BINDER GmbH) and were used for experiments approximately 6 weeks after germination as described before58. Genetically modified plant lines were cultivated in proximity to the appropriate wild-type in the same trays (10 pots per tray) to avoid position effects.
Arabidopsis wild-type Col-0 was used for all standard experiments and as comparison for genetically modified lines, except myc2 (Col-gl background). Plant components functionally involved in the described biocontrol mechanism were identified by testing appropriate established genetically modified Arabidopsis lines in biocontrol assays. The relevance of CK perception was analysed in the Arabidopsis CK-receptor mutant lines ahk2-2/ahk3-3, cre1-12/ahk3-3 and cre1-12/ahk2-2/ahk3-3(+/−) (homozygous for cre1-12 and ahk2-2, heterozygous for ahk3-3). The role of SA was analysed in the SA-deficient lines 35S::nahG (overexpressing SA-degrading SA-hydroxylase from P. putida), sid2 and the SA signalling mutant npr1. Involvement of ET and JA was analysed in the ET insensitive line ein2 and the JA insensitive line myc2. The camalexin-deficient lines cyp79 and pad3 were analysed to determine the role of this Arabidopsis key phytoalexin.
Bacterial strains
The virulent hemibiotrophic bacterial pathogen Pseudomonas syringae pv. tomato DC3000 (Pto) was used for all infections as described before58. For determination of Pto proliferation in planta following pre-treatments with Pseudomonas fluorescens (Pfl) strains, Pto was transformed with pMP466259 to facilitate additional selection against tetracycline which is necessary to avoid unspecific background (co-cultivation) by spontaneous rifampicin resistance of Pfl cells24. Freshly grown (28 °C, 200 rpm) Pto cells from liquid cultures in 50 ml LB medium containing 50 mg l−1 rifampicin (and 20 mg l−1 tetracycline for the pMP4662 transformed strain) were pelleted, re-suspended in 10 mM MgCl2 and adjusted to the desired concentration for the experiments using the BioPhotometer plus (Eppendorf AG).
Pfl G20-18 was tested for its biocontrol ability and subsequently used for biocontrol assays. It was tested against its transposon (TnphoA) mediated CK-deficient mutants CNT1 and CNT224, CNT transposon mutants functionally complemented with functional CK biosynthetic genes (homologous expression of G20-18miaA or heterologous expression of Agrobacterium tumefaciens ipt [Atipt]) in pBBRMCS-5 and a loss-of-function mutant of G20-18 with a distinct disruption of its CK biosynthetic gene miaA (∆miaA). A detailed description of the cloning procedure and generation of Pfl derivatives are available as Supplementary Methods. The different Pfl strains were cultivated in 50 ml LB medium (28 °C, 200 rpm) containing 10 μM adenine24 and appropriate antibiotics: 50 mg l−1 ampicillin for G20-18; 50 mg l−1 ampicillin and 20 mg l−1 gentamycin for G20-18 transformed with pBBRMCS-5; 50 mg l−1 ampicillin and kanamycin for CNT1, CNT2 and ∆miaA; 50 mg l−1 ampicillin, kanamycin and 20 mg l−1 gentamycin for CNT mutants transformed with pBBRMCS-5 derivatives. Pfl cells were processed as described for Pto including a washing step in 30 ml 10 mM MgCl2 before the final resuspension.
Biocontrol experiments
For biocontrol assays, whole Arabidopsis leaves were infiltrated with Pfl cell suspensions (OD600 = 0.02) or 10 mM MgCl2 as a mock control using a needleless syringe two days prior to infection with Pto. Pto infection was performed as described before58 by infiltration of Arabidopsis leaf halves with 105 cfu ml−1 for Pto proliferation determination or 106 cfu ml−1 for analysis of symptom development, respectively.
Pto symptom development (infiltrated leaf halves) was evaluated 4 days post infection (dpi) based on an adapted scale12 consisting of 7 categories (Supplementary Fig. 3). Viable Pfl cells and Pto proliferation in planta were determined similar to published procedures60. Discs of infiltrated leaves were excised at indicated hours post infiltration (hpi) using a cork borer of 0.4 cm diameter, ground and re-suspended in 1 ml 10 mM MgCl2. 100 μl of serial 1:10 dilutions were plated in triplicate on LB medium containing appropriate antibiotics for selection and colony formation was determined after 36 h incubation at 28 °C.
Cytokinin determination
For CK determination, a minimum of 10 Arabidopsis leaves per sample were harvested 48 hpi with Pfl strains or the mock control, immediately frozen and ground in liquid nitrogen. CKs were extracted and determined by UHPLC-MS/MS as described before29. The CK-types quantified in this study are cis-zeatin (cZ), cZ-O-glucoside (cZOG), cZ-riboside (cZR), cZR-O-glucoside (cZROG), cZ-9-glucoside (cZ9G), cZR-5′-monophosphate (cZR5′MP), dihydrozeatin (DHZ), DHZ-O-glucoside (DHZOG), DHZ-riboside (DHZR), DHZR-O-glucoside (DHZROG), DHZ-7-glucoside (DHZ7G), DHZ-9-glucoside (DHZ9G), DHZR-5′-monophosphate (DHZR5′MP), isopentenyladenine (iP), iP-riboside (iPR), iP-7-glucoside (iP7G), iP-9-glucoside (iP9G), iPR-5′-monophosphate (iPR5′MP), trans-zeatin (tZ), tZ-O-glucoside (tZOG), tZ-riboside (tZR), tZR-O-glucoside (tZROG), tZ-7-glucoside (tZ7G), tZ-9-glucoside (tZ9G), tZR-5′-monophosphate (tZR5′MP).
Statistical analysis
Statistical analyses were performed for datasets deriving from a minimum of three biological experiments. Unpaired Student’s t-test was used to compare group differences. P values < 0.05 were considered significant and letters in bar graphs indicate different significance groups. *, ** and *** indicate significant differences at the 0.05, 0.01 and 0.001 levels of confidence, respectively.
Additional Information
Accession codes: The obtained sequence of the Pseudomonas fluorescens G20-18 miaA gene has been deposited in the GenBank database under the accession code KM593658.
How to cite this article: Großkinsky, D. K. et al. Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci. Rep. 6, 23310; doi: 10.1038/srep23310 (2016).
References
Bari, R. & Jones, J. D. G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 69, 473–488 (2009).
Grant, M. R. & Jones, J. D. Hormone (dis)harmony moulds plant health and disease. Science 324, 750–752 (2009).
Pieterse, C. M. J., Leon-Reyes, A., van der Ent, S. & van Wees, S. C. M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 5, 308–316 (2009).
Pieterse, C. M. J., van der Does, D., Zamioudis, C., Leon-Reyes, A. & van Wees, S. C. M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 28, 489–521 (2012).
Sakakibara, H. Cytokinins: activity, biosynthesis and translocation. Annu. Rev. Plant Biol. 57, 431–449 (2006).
Hwang, I., Sheen, J. & Müller, B. Cytokinin signaling networks. Annu. Rev. Plant Biol. 63, 353–380 (2012).
Walters, D. R., McRoberts, W. K. & Fitt, B. D. Are green islands red herrings? Significance of green islands in plant interactions with pathogens and pests. Biol. Rev. Camb. Philos. Soc. 83, 79–102 (2008).
Balibrea Lara, M. E. et al. Extracellular invertase is an essential component of cytokinin-mediated delay of senescence. Plant Cell 16, 1276–1287 (2004).
Ehneß, R. & Roitsch, T. Co-ordinated induction of mRNAs for extracellular invertase and a glucose transporter in Chenopodium rubrum by cytokinins. Plant J. 11, 539–548 (1997).
Choi, J. et al. The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 19, 284–295 (2010).
Argueso, C. T. et al. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 8, e1002448 (2012).
Großkinsky, D. K. et al. Cytokinins mediate resistance against Pseudomonas syringae in tobacco through increased antimicrobial phytoalexin synthesis independent of salicylic acid signaling. Plant Physiol. 157, 815–830 (2011).
Jiang, C. J. et al. Cytokinins act synergistically with salicylic acid to activate defense gene expression in rice. Mol. Plant Microbe Interact. 26, 287–296 (2013).
Großkinsky, D. K., van der Graaff, E. & Roitsch, T. Phytoalexin transgenics in crop protection – Fairy tale with a happy end? Plant Sci. 195, 54–70 (2012).
Großkinsky, D. K., van der Graaff, E. & Roitsch, T. Abscisic acid-cytokinin antagonism modulates resistance against Pseudomonas syringae in tobacco. Phytopathology 104, 1283–1288 (2014).
Ko, K.-W. et al. Effects of cytokinin on production of diterpenoid phytoalexins in rice. J. Pestic. Sci. 35, 412–418 (2010).
Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 84, 11–18 (2009).
Whipps, J. M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 52, 487–511 (2001).
Haas, D. & Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3, 307–319 (2005).
van Wees, S. C., van der Ent, S. & Pieterse, C. M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 11, 443–448 (2008).
Arkhipova, T. N., Veselov, S. U., Melentiev, A. I., Martynenko, E. V. & Kudoyarova, G. R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 272, 201–209 (2005).
Ortíz-Castro, R., Valencia-Cantero, E. & López-Bucio, J. Plant growth promotion by Bacillus megaterium involves cytokinin signaling. Plant Signal. Behav. 3, 263–265 (2008).
Liu, F., Xing, S., Ma, H., Du, Z. & Ma, B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 97, 9155–9164 (2013).
García de Salamone, I. E., Hynes, R. K. & Nelson, L. M. Cytokinin production by plant growth promoting rhizobacteria and selected mutants. Can. J. Microbiol. 47, 404–411 (2001).
Pallai, R., Hynes, K. H., Verma, B. & Nelson, L. M. Phytohormone production and colonization of canola (Brassica napus L.) roots by Pseudomonas fluorescens 6–8 under gnotobiotic conditions. Can. J. Microbiol. 58, 170–178 (2012).
Katagiri, F., Thilmony, R. & He, S. Y. The Arabidopsis thaliana-Pseudomonas syringae interaction. The Arabidopsis Book 1, e0039 (2002).
Naseem, M. et al. Integrated systems view on networking by hormones in Arabidopsis immunity reveals multiple crosstalk for cytokinin. Plant Cell 24, 1793–1814 (2012).
Großkinsky, D. K., Edelsbrunner, K., Pfeifhofer, H., van der Graaff, E. & Roitsch, T. Cis- and trans-zeatin differentially modulate plant immunity. Plant Signal. Behav. 8, e24798 (2013).
Novák, O., Hauserová, E., Amakorová, P., Doležal, K. & Strnad, M. Cytokinin profiling in plant tissues using ultra-performance liquid chromatography-electrospray tandem mass spectrometry. Phytochemistry 69, 2214–2224 (2008).
Koenig, R. L., Morris, R. O. & Polacco, J. C. tRNA is the source of low-level trans-zeatin production in Methylobacterium spp. J. Bacteriol. 184, 1832–1842 (2002).
Ryu, J. et al. Plant growth substances produced by Methylobacterium spp. and their effect on tomato (Lycopersicon esculentum L.) and red pepper (Capsicum annuum L.) growth. J. Microbiol. Biotechnol. 16, 1622–1628 (2006).
Podlešáková, K. et al. Rhizobial synthesized cytokinins contribute to but are not essential for the symbiotic interaction between photosynthetic bradyrhizobia and Aeschynomene legumes. Mol. Plant Microbe Interact. 10, 1232–1238 (2013).
Winter, D. et al. An “electronic fluorescent pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2, e718 (2007).
Compant, S., Duffy, B., Nowak, J., Clément, C. & Barka, E. A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action and future prospects. Appl. Environ. Microbiol. 71, 4951–4959 (2005).
Wu, C. H., Bernard, S. M., Andersen, G. L. & Chen, W. Developing microbe-plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2, 428–440 (2009).
Chalupowicz, L., Barash, I., Panijel, M., Sessa, G. & Manulis-Sasson, S. Regulatory interactions between quorum-sensing, Auxin, cytokinin and the Hrp regulon in relation to gall formation and epiphytic fitness of Pantoea agglomerans pv. gypsophilae. Mol. Plant Microbe Interact. 22, 849–856 (2009).
Depuydt, S. et al. An integrated genomics approach to define niche establishment by Rhodococcus fascians. Plant Physiol. 149, 1366–1386 (2009).
Higuchi, M. et al. In planta functions of the Arabidopsis cytokinin receptor family. Proc. Natl. Acad. Sci. USA 101, 8821–8826 (2004).
Delaney, T. P. et al. A central role of salicylic acid in plant disease resistance. Science 266, 1247–1250 (1994).
Nawrath, C. & Métraux, J.-P. Salicylic acid induction-deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 11, 1393–1404 (1999).
Cao, H., Bowling, S. A., Gordon A. S. & Dong, X. Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6, 1583–1592 (1994).
Berger, S., Bell, E. & Mullet, J. E. Two methyl jasmonate-insensitive mutants show altered expression of AtVsp in response to methyl jasmonate and wounding. Plant Physiol. 111, 525–531 (1996).
Fernández-Calvo, P. et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 23, 701–715 (2011).
Guzmán, P. & Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell 2, 513–523 (1990).
van Peer, R., Niemann, G. J. & Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas sp. strain WCS417r. Phytopathology 81, 1508–1512 (1991).
Ongena, M. et al. Systemic induction of phytoalexins in cucumber in response to treatments with fluorescent pseudomonads. Plant Pathol. 49, 523–530 (2000).
Glazebrook, J. & Ausubel, F. M. Isolation of phytoalexin-deficient mutants of Arabidopsis thaliana and characterization of their interaction with bacterial pathogens. Proc. Natl. Acad. Sci. USA 91, 8955–8959 (1994).
Glawischnig, E., Hansen, B. G., Olsen, C. E. & Halkier, B. A. Camalexin is synthesized from indole-3-acetaldoxime, a key branching point between primary and secondary metabolism in Arabidopsis. Proc. Natl. Acad. Sci. USA 101, 8245–8250 (2004).
Glawischnig, E. Camalexin. Phytochemistry 68, 401–406 (2007).
Rico, A., McCraw, S. L. & Preston, G. M. The metabolic interface between Pseudomonas syringae and plant cells. Curr. Opin. Microbiol. 14, 31–38 (2011).
Gan, S. & Amasino, R. M. Inhibition of leaf senescence by autoregulated production of cytokinin. Science 270, 1986–1988 (1995).
Kuderová, A. et al. Effects of conditional IPT-dependent cytokinin overproduction on root architecture of Arabidopsis seedlings. Plant Cell Physiol. 49, 570–582 (2008).
Li, X. G. et al. Cytokinin overproduction-caused alteration of flower development is partially mediated by CUC2 and CUC3 in Arabidopsis. Gene 450, 109–120 (2010).
Akiyoshi, D. E. et al. Cytokinin/auxin balance in crown gall tumors is regulated by specific loci in the T-DNA. Proc. Natl. Acad. Sci. USA 80, 407–411 (1983).
Akiyoshi, D. E., Klee, H., Amasino, R. M., Nester, E. W. & Gordon, M. P. T-DNA of Agrobacterium tumefaciens encodes an enzyme of cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 81, 5994–5998 (1984).
Hann, D. R. et al. The Pseudomonas type III effector HopQ1 activates cytokinin signaling and interferes with plant innate immunity. New Phytol. 201, 585–598 (2014).
Rivero, R. M. et al. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 104, 19631–19636 (2007).
Großkinsky, D. K., Koffler, B. E., Roitsch, T., Maier, R. & Zechmann, B. Compartment specific antioxidative defense in Arabidopsis against virulent and avirulent Pseudomonas syringae. Phytopathology 102, 662–673 (2012).
Bloemberg, G. V., Wijfjes, A. H. M., Lamers, G. E. M., Stuurman, N. & Lugtenberg, B. J. J. Simultaneous imaging of Pseudomonas fluorescens WCS365 populations expressing three different autofluorescent proteins in the rhizosphere: new perspectives for studying microbial communities. Mol. Plant Microbe Interact. 13, 1170–1176 (2000).
Zipfel, C. et al. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 428, 764–767 (2004).
Acknowledgements
We thank Helga Hammer for superb plant care, Peter Krbez for excellent technical assistance, Kerstin Edelsbrunner for supporting initial miaA identification, Thomas Engelke for supplying pMP4662 transformed Pto, Susanne Berger and Tatsuo Kakimoto for supplying genetically modified Arabidopsis lines and Matthias Ullrich for supplying the vectors pBBR1MCS-5, pK18mobGII and pRK2013. We are grateful to Claus Wasternack for critical reading and constructive comments on the manuscript. This work was supported by funding from the Society for the Advancement of Plant Sciences (Vienna, Austria) and by the Individual Postdoctoral Grant No. 4093-00255 of the Danish Council for Independent Research, Danish Ministry of Higher Education and Science to D.K.G., by an ERASMUS-EADIC II fellowship to M.V.M., funding from the University of Graz and UNCPBA to S.A.S., the Czech Ministry of Education grant from the National Program for Sustainability I, grant numbers LO1204 and 15-22322S, to M.S. and the Czech Ministry of Education, Youth and Sports of CR within the National Sustainability Program I (NPU I), grant number LO1415, to T.R.
Author information
Authors and Affiliations
Contributions
D.K.G. designed and performed the experiments, analysed and interpreted the data and wrote the manuscript. R.T. performed the experiments, analysed and interpreted the data. M.V.M. and S.A.S. performed the experiments and analysed the data. I.E.G.d.S. and L.M.N. supplied material and interpreted the data. O.N. and M.S. performed cytokinin determination and analysed the data. E.v.d.G. analysed and interpreted the data and wrote the manuscript. T.R. designed the experiments, interpreted the data and wrote the manuscript. All authors discussed the results and commented on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Großkinsky, D., Tafner, R., Moreno, M. et al. Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci Rep 6, 23310 (2016). https://doi.org/10.1038/srep23310
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep23310
This article is cited by
-
Microbial antagonists: diversity, formulation and applications for management of pest–pathogens
Egyptian Journal of Biological Pest Control (2023)
-
Secondary metabolites from methylotrophic bacteria: their role in improving plant growth under a stressed environment
Environmental Science and Pollution Research (2023)
-
Developmental Phytohormones: Key Players in Host-Microbe Interactions
Journal of Plant Growth Regulation (2023)
-
Phytohormone Profiling of Potato (Solanum tuberosum L.) Exposed to French Marigold (Tagetes patula L.) Essential Oil
Journal of Plant Growth Regulation (2023)
-
PGPR-Mediated Breakthroughs in Plant Stress Tolerance for Sustainable Farming
Journal of Plant Growth Regulation (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
