
Abstract
Effects of fire on biogeochemical cycling in terrestrial ecosystem are widely acknowledged, while few studies have focused on the bacterial community under the disturbance of long-term frequent prescribed fire. In this study, three treatments (burning every two years (B2), burning every four years (B4) and no burning (B0)) were applied for 38 years in an Australian wet sclerophyll forest. Results showed that bacterial alpha diversity (i.e. bacterial OTU) in the top soil (0–10 cm) was significantly higher in the B2 treatment compared with the B0 and B4 treatments. Non-metric multidimensional analysis (NMDS) of bacterial community showed clear separation of the soil bacterial community structure among different fire frequency regimes and between the depths. Different frequency fire did not have a substantial effect on bacterial composition at phylum level or bacterial 16S rRNA gene abundance. Soil pH and C:N ratio were the major drivers for bacterial community structure in the most frequent fire treatment (B2), while other factors (EC, DOC, DON, MBC, NH4+, TC and TN) were significant in the less frequent burning and no burning treatments (B4 and B0). This study suggested that burning had a dramatic impact on bacterial diversity but not abundance with more frequent fire.
Similar content being viewed by others
Introduction
Forest ecosystem is among the most heterogeneous and complex environments on the planet. Soil microbes represent a considerable fraction of forest ecosystem, regulating nutrient transformations and energy flow in the soil. Soil microbial diversity is of critical importance in maintaining the sustainability of ecosystem and has suffered greatly due to anthropogenic disturbances, such as fertilization1,2, fire3 and deforestation4. Such disturbances may cause shifts in microbial composition resulting in substantial impacts on biogeochemical processes and ecosystem function5. Understanding biotic response to these disturbances is thus critical in predicting the consequences of long-term human-induced changes in forest ecosystem6.
Fire is one of the main concerns of global climate change. Many studies have documented the fire effects on forest ecosystem including enhancing plant productivity7, shifting in plant community structure8, accelerating carbon (C) cycling process9 and reduction of fungal and bacterial biomass10. However, the impacts of fire on forest ecosystem varied greatly with the differences in its intensity, frequency, forest type, the slope, fuel load and type and environmental conditions (e.g. moisture and temperature) as well as the characteristics of different microbial groups (e.g. sensitivity to fire)11. For example, a meta-analysis based on 42 wildfire studies demonstrated that wildfires lead to a more significant reduction in microbial biomass compared to managed burning12. Prescribed fire is one of the common practices reducing the risk of wildfire occurrence, which in contrast causes extensive damage to forest vegetation and soils13. Extensive studies have demonstrated the effectiveness of prescribed fire in wildfire hazard abatement as well as the mitigation of CO2 emissions from forest fires, while little information is available about the impacts of long-term different fire frequencies on soil microbial community in terms of bacterial diversity and abundance14,15, especially individual bacterial taxonomic change16.
Anthropogenic disturbance such as land use conversion, climate change and altered fire regimes, could have major consequences for microbial genetic resources, which in turn influences ecosystem properties like resistance and stability17,18,19. However, the degree to which specific microbial groups is influenced varies with the microbial sensitivity or recovery capacity when subjected to perturbations16,20. For example, bacterial community composition recovered rapidly and returned to the similar condition as no fire interference after burning for 11 years3. Furthermore, microbial community assembly mainly are driven by neutral processes while changed to niche process along the time since fire disturbance21. On the other hand, as fire effect is spatially heterogeneous within one fire event or one area of interest, due to its patchy nature (i.e. mosaics), which could largely influence the relative contribution of niche and neutral processes ascribing to the uneven distribution of fuels, soil moisture and micro-geographical condition (e.g. slope)11,15. Additionally, as disturbance (e.g. prescribed fire) becomes frequent and exerts continuous pressure, severe implications may be apparent and thereby could cause long-term cumulated negative impacts on key biogeochemical processes22. Therefore, it is of critical importance to understand the role of recurring fire disturbance for microbial groups and environmental assessment23.
It is generally difficult in determining the disturbance effects, which are normally confounded with other factors. Therefore, it is critical to find out the key factors as well as the ecological process driving microbial community composition. Pyrosequencing techniques has shown to be a powerful tool in discovering microbial diversity and community composition via deciphering the distribution of microbial groups and their interaction with abiotic factors21,24,25. We hypothesized that more frequent fire disturbance would have a greater impact on soil bacterial diversity compared to less frequent one due to differences in soil parameters mediated by fire. The objectives of this study were, therefore, to i) determine the effects of long-term prescribed fire on the diversity of bacteria; and ii) identify the main factors contributing to alterations of the bacterial community composition using pyrosequencing technique targeting 16S rRNA region.
Results
Soil chemical properties
Soil chemical properties were greatly impacted by frequent fire at the Peachester site (Table 1). At the depth of 0–10 cm, soil total C (TC), total N (TN), moisture, dissolved organic C (DOC), dissolved organic N (DON) and microbial biomass C (MBC) were all significantly lower in the more frequent fire treatment (B2) than in the less frequent (B4) and no-burning (B0) treatments. The highest soil pH value (ca. 5.41) was recorded in the B2 treatment, followed by the B4 treatment (5.11) and B0 treatment (4.86), while electrical conductivity (EC) showed the opposite trend (Table 1). No significant difference among the treatments was detected at the 10–20 cm depth for all the measured soil properties, except soil moisture. Some of the resource-related variables (i.e. TC, TN, DOC, DON, NH4+, MBC and microbial biomass N (MBN)) at the 10–20 cm depth were significantly lower compared to the 0–10 cm depth for B4 and B0 treatments, but no differences between depths were observed for the B2 treatment except for DOC (Table 1). There was no significant difference in the inorganic phosphorus (Pi), C:N ratio and nitrate (NO3−) irrespective of depth or treatment though the variance was high for nitrate.
Soil bacterial diversity and the relationship with soil environmental properties
In total, 295,185 quality sequences were obtained after eliminating one sample with 411 sequences, resulting into 2,753 to 18,210 sequences per sample (Table S1). Bacterial community diversity was calculated based on the relative abundance of OTU. In the top soil the most frequent fire treatment (B2) had significantly higher bacterial alpha-diversity relative to the other treatments, regardless of the parameters (i.e. OTU, Chao1) based on OTU matrix (Fig. 1). No significant difference between treatments was detected for the Shannon indice. There were no significant treatment effects in all alpha-diversity indexes measured for the 10–20 cm depth. A similar trend of highest diversity in the B2 treatment was also observed by rarefaction curves as measured by OTU richness with average values for triplicate treatments irrespective of sample size (Fig. S1 A and B).

Boxplot of bacterial alpha-diversity indices for the 16S rRNA gene sequences among different treatments at two depths.
B0, B2 and B4 represent the 0–10 cm samples from the treatments of no burning, burning every two years and burning every four years, respectively, while sB0s, sB2 and sB4 represent the 10–20 cm samples. Top two panels are bacterial diversity indices derived from soil samples at the depth of 0–10 cm and bottom two panels are at the depth of 10–20 cm. Fire effects across treatments indicated by asterisks: *P < 0.5.
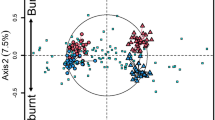
Non-metric multidimensional scaling clustered soil samples by depth and treatment (P = 0.0099, Monte carlo randomization test) (Fig. 2). The most frequent burning treatment (B2) was clearly separated from the other treatments (B4 and B0) within each depth. The separation of community structure was supported by an analysis of similarity of bacterial communities (ANOSIM) for different treatments at each depth (Table 2). ANOSIM of Bray-Curtis and UniFrac distance dissimilarities for depth and frequent fire effects both showed significant differences (P < 0.05), but there was no difference between the burnt (B2 and B4) and unburnt (B0) treatments (P > 0.05) (Table 2).

Non-metric multidimensional scaling plot of soil bacterial community structure in three fire treatments and at two depths using OTU-based Bray-Curtis dissimilarities distance with soil chemical properties.
B0, B2 and B4 represent the 0–10 cm samples from the treatments of no burning, burning every two years and burning every four years, respectively, while sB0s, sB2 and sB4 represent the 10–20 cm samples. Only soil chemical properties with P values less than 0.5 are plotted and the arrow length is proportional to the strength of correlation. Stress value is also indicated in the figure. For abbreviations for soil chemical properties refer to Table 1. The R2 value for stress test is 0.976 (non-metric fit) and 0.875 (linear fit).
A Mantel test further revealed the relationship between soil bacterial community structure and abiotic factors under the different frequent fire treatments. There were significant correlations (P < 0.01) between soil properties (i.e. DOC, DON, pH, EC and NH4+) and bacterial community composition generated by Bray-Curtis (OTU-based) dissimilarities (Table 3). In order to eliminate bias caused by rare OTUs, we also calculated the correlation between soil properties and the dominate OTUs (66 OTUs) and this analysis revealed the similar trends to those calculated using most OTUs (Table 3).
Bacterial 16S rRNA gene composition and abundance
A taxonomic summary of bacterial composition showed that all soils were typically dominated by Acidobacteria, ranging from 36% to 42% of all sequences across the treatments (Fig. 3). More interestingly, phylum Verrucomicrobia appeared to be the second most dominant group (24–39%), which was more abundant than the Proteobacteria group (12–23%) (t23,23 = 4.7, P < 0.001) (Fig. 3). The relative abundance of Verrucomicrobia was much higher at the 10–20 cm depth compared to the top soil (0–10 cm), while Alphaproteobacteria and Bacteroidetes showed an opposite trend along the depth (Fig. 3). There was no significant difference in the relative abundance of dominant phyla among treatments using Chi-Square tests. However, significant responses were detected for the most abundant OTU at lower levels of taxon between burning and non-burning treatments (Fig. 4). For example, the most detected subgroup Gp1 Acidobacteria had a significantly lower abundance except OTU_45 in response to burning (Fig. 4). For Alphaproteobacteria, in the order Rhizobiales, most OTUs showed higher abundance especial to the most frequent burning (Fig. 4b). Similarly, by comparing to no burning treatment, Verrucomicrobia taxa showed a highly variable response in burning every two years (B2) than every four years (B4). Spearman’s correlations were measured between the relative abundance of the main phyla and soil chemical properties to assess the relative contribution of biotic and abiotic factors on the bacterial community composition. Verrucomicroia and Chloroflexi were negatively correlated with some soil properties (i.e. DOC, DON, MBC and MBN), while Gammaproteobacteria and Bacteroidetes were positively correlated with most of the soil properties (Table S2).

Percent community composition of numerically dominant bacterial phyla across three fire treatments at two depths.

Response ratios of (a) burning every 4 years to no burning treatment and (b) burning every 2 years to no burning treatment. Significance was determined at a 95% CI (confidence interval).
As detected by real-time PCR targeting 16S rRNA gene, bacterial abundance were recorded from 4.67 × 109 in the B2 treatment to 2.96 × 109 copy numbers per gram dry soil in the sB4 treatment. There was no significant difference in soil bacterial 16S rRNA gene abundance across the treatments or the depths (Fig. S2). This result is consistent with the bacterial 16S rRNA gene quantitative results of soil samples taken from the same site in 200526 (Fig. S2).
Discussion
Fire has modified majority of earth surface and it has not only affect the aboveground species composition, but also the belowground community5. In this study, NMDS analyses based on pyrosequencing data showed that soil bacterial communities in the most frequently burnt treatment were significantly different from those in the less frequently burnt and long unburnt treatments, especially in the topsoil. Furthermore, a significant increase in bacterial alpha diversity (i.e. OTU and Chao1) was detected in topsoils of the most frequent burning treatment (B2), which is congruent with the observations for fungal communities at the same site27 and with a study of similar fire regimes in the United States28. ANOSIM analysis based on the Bury-curtis and Unweighted UniFrac distances also showed that bacterial community structure was significantly affected by the most frequent fire treatment as well as depth. The increase in bacterial community diversity in response to frequent fire may be partly attributable to the spatial heterogeneity of fire effects caused by the inherent patchiness of low intensity fire due to uneven distribution of fuels, slope position and soil moisture in wet eucalypt forest11,29. Fire disturbance would increase clumping of fire-intolerant species, such as in frequently burned pine savannas of southeastern North America, resulting in higher β-deviations as well as the stronger link with environmental factors in burning treatments comparing to non-burning30. Microbial diversity tended to increase following wildfire disturbance over time21. Fire disturbance creates heterogeneity in forest soil properties that may maintain and even improve the bacterial diversity by affecting the competition ability of bacteria for resources31,32,33. This phenomenon is compatible with Connell’s intermediate disturbance hypothesis proposing that maximum levels of diversity are obtained at intermediate frequencies of disturbance as corroborated by previous studies34. Indeed, more recent research focusing on diversity-disturbance relationships has found positive relationships between diversity and both disturbance intensity and frequency35, which has further corroborated our finding.
The observed shift of bacterial community structure was more directly attributable to the change of soil chemical factors as evidenced by the significant correlation between bacterial community structure and selected soil properties with Mantel test using OTU-based dissimilarity (Table 3). Among the selected variables, soil pH was one of the major factors contributing to the observed bacterial community variations, showing a positive correlation with frequent fire treatments. Fires, whether prescribed or wildfire, can increase soil pH due to the deposition of ash20,36 and therefore drive a shift in the bacterial community structure37. Accumulating literature has already documented this pH-driver phenomena for different microbial groups, including bacteria38, fungi39 and some specific taxa Acidobacteria40. However, it is still unclear whether communities were influenced directly by pH or indirectly through influencing other factors altered by pH37. Other research on fire regimes suggested that microbial structure is largely related to the availability of substrates, but not soil pH20. Furthermore, it has been suggested that the reduction of total C and N stocks contributed to the shifts of the general fungal and ectomycorrhizal communities at the same site27,41. As such, we proposed that soil pH was the main factor for the shift of bacterial community structure under the most frequent fire disturbance (e.g. B2), while for those treatments with little variation in soil pH (i.e. B4 and B0), resource availability mainly attributed to the distribution pattern of bacteria.
In contrast to bacterial community structure, little variation of bacterial abundance was observed across the fire treatments in this study based on quantitative analysis of 16S rRNA gene using real-time PCR technique. This finding is consistent with earlier studies at the same site for the species richness of Basidiomycete fungal communities27 and a seasonal comparison of bacterial 16S rRNA gene copies in 201142. When combined with results from previous measurements for bacterial 16S rRNA gene copies from 2005 and 2011, all showed a consistent trend (i.e. no significant difference among fire treatments) using real-time PCR technique1,42. The real-time PCR technique is a robust method and has been widely used in microbial abundance analysis. However, it cannot be ruled out that variation in the abundance of specific bacterial groups at lower taxonomic levels may occur in response to fire. Furthermore, the variable impacts of frequent fire on bacterial diversity and relative abundance at phylum level were largely driven by the less frequent taxa that responded rapidly to recurring fire disturbance28,43. Indeed, a comparison of unique OTUs and relative abundance of sequences for each dominant bacterial taxon further supported this finding (Fig. S3). The classification of bacterial groups was recently been proposed as copiotrophic and oligotrophic according to nutrition and growth44. This ecological feature of bacterial phyla was largely related to the variation in relative abundance of dominant taxa following exotic disturbance45,46. Bacteroidetes and Proteobacteria, for instance, are typically copiotrophic and respond rapidly to the availability of resources, such as soluble carbon44,47. This is supported by the positive relationship between relative abundance for these two groups and soil resource variables (i.e. DOC, DON, NH4+, MBC and MBN) in our study. In contrast, an unexpected higher abundance of Verrucomibia was observed, with negative correlations with soil variables DOC, DON, MBC and MBN, suggesting their slow-growing life strategy is favoured by low resource habitats48.
This study clearly demonstrated for the first time that significant alteration of bacterial community diversity and structure by the long-term frequent fire disturbance. Biennial fire greatly changed bacterial community structure, while minimal effects on bacterial composition and abundance. Moreover, this study has also shown the dominance of taxa Verrucomicrobia, implying that this group may play an important role in biogeochemical processes, such as C cycling49. Further work on its contribution to ecosystem function is required to better understand the response of microbial communities to frequent and severe disturbances. Moreover, future work also needs to look into the time series of microbial community pattern in order to predict the response of microbial groups as well as assess the recover capacity for recurring disturbances.
Methods
Sampling site
Our experimental site is located in Peachester State Forest, southeast Queensland, Australia (26°50′S, 152° 53′E), which has been described in detail previously42,50. In brief, three prescribed burning treatments, i.e. burning every two years (B2), burning every four years (B4) and no burning (B0), were established in 1972 and low intensity fires (<2,500 kWm−1) were carried out in the winter. The Peachester site has an average annual rainfall of 1,711 mm and average minimum and maximum temperatures of 14 °C to 24 °C, respectively. It is a typical wet sclerophyll forest and the vegetation is dominated by Eucalyptus pilularis Smith with lesser proportions of other canopy species including Corymbia intermedia, Eucalyptus microcorys, Eucalyptus resinifera, Syncarpia glomulifera and Lophostemon confertus. Our previous research in this experiment site has demonstrated that long-term repeated burning had significant impact on vegetation community structure and composition51. Soil samples were taken at depths of 0–10 cm and 10–20 cm in July 2010, before the next scheduled burning (August 2010) for the B2 and B4 treatments. Fifteen soil cores were randomly taken from each plot and bulked as a composite sample and in total 24 composite samples (across two depths) were collected from four plots (replicates) in each treatment. Soil samples were passed through a 2.0- mm sieve after removing root debris and stones and kept at 4 °C for chemical and biological analyses. Subsamples were kept at −80 °C for further analysis.
Soil chemical properties analysis
Soil moisture was determined gravimetrically via drying the soil at 105 °C for 24 h and methods for soil chemical properties have been described in previous research52. Soil pH and electrical conductance (EC) were measured in the ratio of 1:5 (soil-to-water) with a pH/EC meter. Microbial biomass C (MBC) and N (MBN) were measured by the chloroform fumigation extraction method. Soil samples were extracted using 2 M KCl solution for analysis of ammonium (NH4+-N) and nitrate (NO3−-N) with Westco Smart Chem Discrete Wet Chemistry Analyzer (Westco Scientific Instruments, USA). Soil total C (TC) and N (TN) content were measured with a Eurovector Elemental Analyser (Isoprime-EuroEA 3000, Milan, Italy). Dissolved organic carbon (DOC) and total dissolved organic nitrogen (DON) were extracted by deionized water (1:5, soil:water) via incubating at 70 °C for 16 hours, then measured with a TOC-VCPH/CPN analyser fitted with a TN unit (Shimadzu Scientific Instruments, Columbia, USA).
Soil DNA isolation and pyrosequencing
The total genomic DNA was extracted from 0.3 g of frozen soil using MoBio Powersoil DNA isolation kit following the manufacture’s instruction. 10-fold dilution of DNA template was used for downstream PCR analysis to reduce PCR inhibitors such as humic acid. PCR products for pyrosequencing were amplified with the bar-coded primers F515 (5′-GTGCCAGCMGCCGCGGTAA-3′) and R806 (5′-GGACTACVSGGGTATCTAAT-3′), targeting the V4 region of the bacterial and archaeal 16S rRNA genes53,54. The forward primer consisted of the 454 adapter A, a 10-based MID barcodes, 4-bp linker sequence (TCAG), along with the primer F515. The reverse primer consisted of the 454 adapter B, 4-bp linker sequence (TCAG) along with the primer R806. PCR amplification were performed in triplicate in 30 μl reaction containing 1 × PCR buffer, 3.0 mM MgCl2, 400 μM each dNTP, 2.5 U Ex TaqTM DNA polymerase (TaKaRa, Shiga, Japan), 0.5 μM of each bar-coded primer. We used the following PCR thermal profile: 94 °C for 5 min, 34 cycles consisting of 95 °C for 30 s, 53 °C for 45 s, 72 °C for 60 s and final extension at 72 °C for 60 s. Pooled PCR products for each soil sample were purified using a Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI). The purity and concentration were checked using a NanoDrop 1000 spectrophotometer (Thermo Scientific, Wilmington, DE) and Qubit fluorometer with Quant-iT dsDNA HS Assay Kits (Invitrogen, Eugene, OR), respectively. Equimolar amounts of PCR products with different barcodes were combined into one sample and sent to Macrogen (Seoul, Korea) for pyrosequencing using Roche/454 GS FLX Titanium platform.
In total, 43, 8678 raw sequences were obtained from 24 samples. Mothur 1.30.1 software55 was employed to process 454 raw data. Sequences were trimmed and aligned for quality check with Mothur pipeline after deionising with the command of Shhh.flows, following the main steps of previous research56. Sequences with eight or more homopolymers and two or more ambiguous bases were removed. Filtered sequences were aligned against the SILVA reference database57 for chimera check using chimera.uchime function58. There were 7,654 singleton sequences removed before final diversity analysis59. The bacterial sequences were eventually binned into operational taxonomic units (OTU) to generate a rarefaction curve and to calculate species richness estimators at the sequence identity level of 97% using the average neighbor method. OTU as a measure of a single sample alpha diversity indicates the richness that the numbers of OTUs are observed. Rarefaction curve is one of the commonly used species richness estimators to determine whether sampling depth is sufficient to accurately characterize the bacterial community60. The Chao 1 index as another commonly used species richness estimator will also be used in our study based on the number of rare OTUs in a sample61. The Shannon index accounts for both abundance and evenness of the species present, which is not sensitive to sample size. The taxonomic assignment of representative sequences was achieved using the Ribosomal Database Project (RDP) Classifier62 with a minimum confidence of 80%.
Alpha diversity was calculated based on the relative abundance of OTU. In order to minimize the variation caused by the sampling sequences, each sample was rarefied to the minimum sequence number (2,753 sequences) using subsample argument in Mothur.
Quantitative PCR for 16S rRNA gene was carried out using primer pair 338F/518R and PCR thermal profile and standard curve preparation followed the procedure as described in detail by Liu et al.42. Amplification efficiency for 16S rRNA gene was 95–98% with R2 values greater than 0.99 for the standard curve.
Pyrosequencing data analysis
In total, 295,596 quality sequences were obtained from 24 samples with the fragment length ranging from 235 to 313 bp. However, one of the samples from the burning every four years (B4) with the minimum number of sequences (i.e. 411) was eliminated for subsequent analysis of alpha and beta diversity, eventually, resulting into 2,753–18,210 sequences per sample after data trimming and quality control (Table S1). There were 35.2%, 22.5%, 11.20%, 10.49%, 6.20% and 6.19% of OTUs affiliated with Proteobacteria, Acidobacteria, Planctomycetes, Verrucomicrobia, Actinobacteria and Bacteroidetes (Fig. S3 bottom panel), respectively. These six phyla contributed to 91.8% of total OTUs. Approximately 0.7% OTUs could not be assigned to any phylum. Of all the assigned OTUs at 97% similarity, the most abundant OTU belonged to Verrucomicrobia, which accounted for 20.4% of all the sequences.
Statistical analysis
Before performing ordination and correlation analysis based on the OTU matrix, those OTUs with less than five detected sites were eliminated in order to achieve a robust result (eventually 736 unique OTU were selected). Relationships between bacterial community distribution data and soil chemical parameters were evaluated with a Mantel test using OTU-based Bray-Curtis distance matrices with 999 permutations in R software Vegan v2.0–8 package. Euclidean distance was used to generate a soil chemical parameters distance matrix. Ordination and then the relationship of these data with soil chemical parameters was estimated using evfit function of Vegan v2.0–8 library with 999 permutations. The accuracy of the two-dimensional representation is indicated by the “stress” value. Distances between symbols represent relative dissimilarity between communities. We also did a Monte Carlo test to check the statistical significance of the generated two dimensions (k = 2). To determine if any group of samples contained significantly different bacterial communities, the analysis of similarities (ANOSIM) function was used with Bray-Curtis (OTU-based) dissimilarity matrices and UniFrac (Phylogenetically-based) distance using the statistical software R, Vegan package63. All the tests were performed using R v2.15.3 statistical program (http://www.r-project.org).
One-way ANOVA was used for comparison of bacterial diversity indices using SPSS 16.0. All values including soil chemical properties were log or square-root transformed to meet test assumptions. Spearman’s rank correlation was calculated to compare the relationship within soil chemical properties and with the composition of taxa associated with fire treatments using SPSS Bivariate correlations. To test heterogeneity in frequency of taxon among different burning treatments, chi-square tests were performed with the relative abundance of dominant phyla using SPSS 16.0. Response ratio was applied to analyse the effects of different frequent fire on the main dominant OTU (the 50 most dominant OTUs) compared to non-burning site using the calculation methods as suggested by Luo et al.64 and Liang et al.65. Significance was determined at a 95% CI (confidence interval).
Additional Information
Accession codes: The sequence data have been submitted to the GenBank database under SRA accession number SRP032649. Sample site and barcode information are both associated with NCBI SRA submission.
How to cite this article: Shen, J.- et al. Long term repeated fire disturbance alters soil bacterial diversity but not the abundance in an Australian wet sclerophyll forest. Sci. Rep. 6, 19639; doi: 10.1038/srep19639 (2016).
References
Long, X., Chen, C. R., Xu, Z. H., Oren, R. & He, J. Z. Abundance and community structure of ammonia-oxidizing bacteria and archaea in a temperate forest ecosystem under ten-years elevated CO2 . Soil Biol & Biochem 46, 163–171 (2012).
Turlapati, S. A. et al. Chronic N-amended soils exhibit an altered bacterial community structure in Harvard Forest, MA, USA. FEMS Microbiol Ecol 83, 478–493 (2013).
Xiang, X. J. et al. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci Rep 4 (2014).
Muller, D. et al. Regime shifts limit the predictability of land-system change. Global Environ Chang 28, 75–83 (2014).
Ward, S. E. et al. Fire accelerates assimilation and transfer of photosynthetic carbon from plants to soil microbes in a northern peatland. Ecosystems 15, 1245–1257 (2012).
Falkowski, P. G., Fenchel, T. & Delong, E. F. The microbial engines that drive Earth’s biogeochemical cycles. Science 320, 1034–1039 (2008).
Blair, J. M. Fire, N availability and plant response in grasslands: A test of the transient maxima hypothesis. Ecology 78, 2359–2368 (1997).
Heisler, J. L., Briggs, J. M., Knapp, A. K., Blair, J. M. & Seery, A. Direct and indirect effects of fire on shrub density and aboveground productivity in a mesic grassland. Ecology 85, 2245–2257 (2004).
Niboyet, A. et al. Global change could amplify fire effects on soil greenhouse gas emissions. Plos One 6 (2011).
Campbell, C. D., Cameron, C. M., Bastias, B. A., Chen, C. R. & Cairney, J. W. G. Long term repeated burning in a wet sclerophyll forest reduces fungal and bacterial biomass and responses to carbon substrates. Soil Biol Biochem 40, 2246–2252 (2008).
McMullan-Fisher, S. J. M. et al. Fungi and fire in Australian ecosystems: a review of current knowledge, management implications and future directions. Aust J Bot 59, 70–90 (2011).
Dooley, S. R. & Treseder, K. K. The effect of fire on microbial biomass: a meta-analysis of field studies. Biogeochemistry 109, 49–61 (2012).
Turetsky, M., Wieder, K., Halsey, L. & Vitt, D. Current disturbance and the diminishing peatland carbon sink. Geophys Res Lett 29 (2002).
Wardle, D. A. et al. Ecological linkages between aboveground and belowground biota. Science 304, 1629–1633 (2004).
Parr, C. L. & Andersen, A. N. Patch mosaic burning for biodiversity conservation: a critique of the pyrodiversity paradigm. Conserv Biol 20, 1610–1619 (2006).
Coolon, J. D., Jones, K. L., Todd, T. C., Blair, J. M. & Herman, M. A. Long-term nitrogen amendment alters the diversity and assemblage of soil bacterial communities in tallgrass prairie. Plos One 8 (2013).
Wolters, V. et al. Effects of global changes on above- and belowground biodiversity in terrestrial ecosystems: Implications for ecosystem functioning. Bioscience 50, 1089–1098 (2000).
Girvan, M. S., Campbell, C. D., Killham, K., Prosser, J. I. & Glover, L. A. Bacterial diversity promotes community stability and functional resilience after perturbation. Environ Microbiol 7, 301–313 (2005).
De Vries, F. T. & Shade, A. Controls on soil microbial community stability under climate change. Front Microbiol 4 (2013).
Baath, E., Frostegard, A., Pennanen, T. & Fritze, H. Microbial community structure and pH response in relation to soil organic-matter quality in wood-ash fertilized, clear-cut or burned coniferous forest soils. Soil Biol Biochem 27, 229–240 (1995).
Ferrenberg, S. et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. ISME J 7, 1102–1111 (2013).
Ives, A. R. & Carpenter, S. R. Stability and diversity of ecosystems. Science 317, 58–62 (2007).
Villnas, A. et al. The role of recurrent disturbances for ecosystem multifunctionality. Ecology 94, 2275–2287 (2013).
Buee, M. et al. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol 184, 449–456 (2009).
Hu, H. W., Zhang, L. M., Yuan, C. L. & He, J. Z. Contrasting Euryarchaeota communities between upland and paddy soils exhibited similar pH-impacted biogeographic patterns. Soil Biol Biochem 64, 18–27 (2013).
Long, X., Chen, C., Xu, Z. & He, J. Shifts in the abundance and community structure of soil ammonia oxidizers in a wet sclerophyll forest under long-term prescribed burning. Sci Total Environ 470–471, 578–586 (2014).
Anderson, I. C., Bastias, B. A., Genney, D. R., Parkin, P. I. & Cairney, J. W. G. Basidiomycete fungal communities in Australian sclerophyll forest soil are altered by repeated prescribed burning. Mycol Res 111, 482–486 (2007).
Brown, S. P., Callaham, M. A., Oliver, A. K. & Jumpponen, A. Deep Ion Torrent sequencing identifies soil fungal community shifts after frequent prescribed fires in a southeastern US forest ecosystem. Fems Microbiol Ecol, 86(3), 557–66 (2013).
Burrows, N. D. Linking fire ecology and fire management in south-west Australian forest landscapes. For Ecol Manage 255, 2394–2406 (2008).
Crandall, R. M. & Platt, W. J. Habitat and fire heterogeneity explain the co-occurrence of congeneric resprouter and reseeder Hypericum spp. along a Florida pine savanna ecocline. Plant Ecol 213, 1643–1654 (2012).
Chesson, P. Mechanisms of maintenance of species diversity. Annu Rev Ecol Syst 31, 343–366 (2000).
Artz, R. R. E., Reid, E., Anderson, I. C., Campbell, C. D. & Cairney, J. W. G. Long term repeated prescribed burning increases evenness in the basidiomycete laccase gene pool in forest soils. Fems Microbiol Ecol 67, 397–410 (2009).
Otsuka, S. et al. Community Structure of Soil Bacteria in a Tropical Rainforest Several Years After Fire. Microbes and Environ 23, 49–56 (2008).
Kassen, R., Buckling, A., Bell, G. & Rainey, P. B. Diversity peaks at intermediate productivity in a laboratory microcosm. Nature 406, 508–512 (2000).
Tanentzap, A. J., Lee, W. G. & Schulz, K. A. C. Niches drive peaked and positive relationships between diversity and disturbance in natural ecosystems. Ecosphere 4, art133 (2013).
Certini, G. Effects of fire on properties of forest soils: a review. Oecologia 143, 1–10 (2005).
Rousk, J. et al. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J 4, 1340–1351 (2010).
Fierer, N. & Jackson, R. B. The diversity and biogeography of soil bacterial communities. P Natl Acad Sci USA 103, 626–631 (2006).
Rousk, J., Brookes, P. C. & Baath, E. Investigating the mechanisms for the opposing pH relationships of fungal and bacterial growth in soil. Soil Biol Biochem 42, 926–934 (2010).
Jones, R. T. et al. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J 3, 442–453 (2009).
Bastias, B. A., Huang, Z. Q., Blumfield, T., Xu, Z. H. & Cairney, J. W. G. Influence of repeated prescribed burning on the soil fungal community in an eastern Australian wet sclerophyll forest. Soil Biol Biochem 38, 3492–3501 (2006).
Liu, X. et al. Soil environmental factors rather than denitrification gene abundance control N2O fluxes in a wet sclerophyll forest with different burning frequency. Soil Biol Biochem 57, 292–300 (2013).
Jumpponen, A. & Jones, K. L. Massively parallel 454 sequencing indicates hyperdiverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol 184, 438–448 (2009).
Fierer, N., Bradford, M. A. & Jackson, R. B. Toward an ecological classification of soil bacteria. Ecology 88, 1354–1364 (2007).
Fierer, N. et al. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J 6, 1007–1017 (2012).
Nemergut, D. R., Cleveland, C. C., Wieder, W. R., Washenberger, C. L. & Townsend, A. R. Plot-scale manipulations of organic matter inputs to soils correlate with shifts in microbial community composition in a lowland tropical rain forest. Soil Biol Biochem 42, 2153–2160 (2010).
Smit, E. et al. Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. App Environ Microbiol 67, 2284–2291 (2001).
Cruz-Martinez, K. et al. Despite strong seasonal responses, soil microbial consortia are more resilient to long-term changes in rainfall than overlying grassland. ISME J 3, 738–744 (2009).
Fierer, N. et al. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the United States. Science 342, 621–624 (2013).
Toberman, H., Chen, C., Lewis, T. & Elser, J. J. High frequency fire alters C:N:P stoichiometry in forest litter. Global Change Biol 20, 2321–2331 (2013).
Lewis, T., Reif, M., Prendergast, E. & Tran, C. The effect of long-term repeated burning and fire exclusion on above- and below-ground Blackbutt (Eucalyptus pilularis) forest vegetation assemblages. Aust Ecol 37, 767–778 (2012).
Chen, C. R., Xu, Z. H., Zhang, S. L. & Keay, P. Soluble organic nitrogen pools in forest soils of subtropical Australia. Plant Soil 277, 285–297 (2005).
Lauber, C. L., Hamady, M., Knight, R. & Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. App Environ Microbiol 75, 5111–5120 (2009).
Bates, S. T. et al. Examining the global distribution of dominant archaeal populations in soil. ISME J 5, 908–917 (2011).
Schloss, P. D. et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. App Environm Microbiol 75, 7537–7541 (2009).
Hodkinson, B. P., Gottel, N. R., Schadt, C. W. & Lutzoni, F. Photoautotrophic symbiont and geography are major factors affecting highly structured and diverse bacterial communities in the lichen microbiome. Environ Microbiol 14, 147–161 (2012).
Quast, C. et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41, D590–D596 (2013).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200 (2011).
Kunin, V., Engelbrektson, A., Ochman, H. & Hugenholtz, P. Wrinkles in the rare biosphere: pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ Microbiol 12, 118–123 (2010).
Gotelli, N. J. & Colwell, R. K. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4, 379–391 (2001).
Chao, A. Nonparametric estimation of the number of classes in a population. Scand J Stat 11, 265–270 (1984).
Wang, Q., Garrity, G. M., Tiedje, J. M. & Cole, J. R. Naïve bayesian classifier for rapid sssignment of rRNA sequences into the new bacterial taxonomy. App Environ Microbiol 73, 5261–5267 (2007).
Oksanen, J. K. R., Legendre, P., O’Hara, B., Simpson, G. L., Solymos, P., Stevens, M. H. & Wagner, H. vegan: Community Ecol Pack (2009).
Luo, Y. Q., Hui, D. F. & Zhang, D. Q. Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A meta-analysis. Ecology 87, 53–63 (2006).
Liang, Y. T. et al. Microarray-based analysis of microbial functional diversity along an oil contamination gradient in oil field. FEMS Microbiol Ecol 70, 324–333 (2009).
Acknowledgements
We would like to thank Marijke Heenan, Xian Liu and Xien Long for their assistance in sampling and lab analyses, also thanks to the Department of Agriculture, Fisheries and Forestry, Queensland, Australia for the access to the Peachester State Forest experimental site. This study was supported by ARC Future Fellowship (FT0990547) and Griffith University.
Author information
Authors and Affiliations
Contributions
C.C. and T.L. conceived the experiments, J.S. and C.C conducted the experiments, analysed the results and wrote the paper. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Shen, Jp., Chen, C. & Lewis, T. Long term repeated fire disturbance alters soil bacterial diversity but not the abundance in an Australian wet sclerophyll forest. Sci Rep 6, 19639 (2016). https://doi.org/10.1038/srep19639
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep19639
This article is cited by
-
Anthropogenic evidences as precursors to forest fire trigger in Western Himalayan Region
Environment, Development and Sustainability (2023)
-
Substrate and low intensity fires influence bacterial communities in longleaf pine savanna
Scientific Reports (2022)
-
The soil microbiomes of the Brazilian Cerrado
Journal of Soils and Sediments (2021)
-
The multi-element stoichiometry of wet eucalypt forest is transformed by recent, frequent fire
Plant and Soil (2020)
-
Assessment of yeasts in tropical peat swamp forests in Thailand
Mycological Progress (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
