
Abstract
Salt stress inhibits soybean growth and reduces gain yield. Genetic improvement of salt tolerance is essential for sustainable soybean production in saline areas. In this study, we isolated a gene (Ncl) that could synchronously regulate the transport and accumulation of Na+, K+ and Cl− from a Brazilian soybean cultivar FT-Abyara using map-based cloning strategy. Higher expression of the salt tolerance gene Ncl in the root resulted in lower accumulations of Na+, K+ and Cl− in the shoot under salt stress. Transfer of Ncl with the Agrobacterium-mediated transformation method into a soybean cultivar Kariyutaka significantly enhanced its salt tolerance. Introgression of the tolerance allele into soybean cultivar Jackson, using DNA marker-assisted selection (MAS), produced an improved salt tolerance line. Ncl could increase soybean grain yield by 3.6–5.5 times in saline field conditions. Using Ncl in soybean breeding through gene transfer or MAS would contribute to sustainable soybean production in saline-prone areas.
Similar content being viewed by others

Introduction
Soybean [Glycine max (L.) Merr.] is the world’s primary crop source for protein and oil. The total world soybean production is 268.0 million metric tons in 2012, providing 68% of world protein meal consumption and 28% world vegetable oil consumption (www.soystats.com). Soybean cultivation is conducted in a wide range of environments and is exposed to many biotic and abiotic stresses that influence the sustainability of soybean production. Soybean is generally regarded as a salt-sensitive crop compared with other major crops such as wheat, rice and cotton. Salt stress inhibits soybean germination and plant growth1,2, nodule formation3 and seed yield4.
Genetic variation for salt tolerance has been described in soybean1,4,5,6, The salt tolerance of partial soybean germplasm conserved in the GRIN, USDA has been evaluated. One hundred and fifty one germplasms, such as Lee, Lee 68 and S-100, were identified to be salt-tolerant, whereas 413 germplasms were designated as susceptible to salt reaction (www.ars-grin.gov/npgs/searchgrin.html). Salt tolerance variation was also observed in wild soybean [Glycine soja Sieb. & Zucc.] and some wild soybean accessions were identified to be salt-tolerant7,8,9,10,11,12. The high level of variation in soybean germplasm, including wild and cultivated soybean species, suggest that genetic improvement of salt tolerance is feasible.
The heredity of salt tolerance in soybean was previously analyzed as a quality trait using the classical genetics approach13,14 and the gene symbols Ncl and ncl were proposed as the dominant for tolerance and the recessive for sensitive, respectively13. The recent development of soybean molecular maps provides a promising approach in identifying genes associated with traits of interest in soybean15,16. Several quantitative trait loci (QTLs) for salt tolerance were reported in different germplasms8,10,17,18,19. Of these, a major QTL for salt tolerance was constantly detected on soybean chromosome 3 (Linkage group N) in different populations8,10,17,19. The QTL on chromosome 3 is likely to be the Ncl locus based on pedigree tracing17. The salt tolerance QTL, in which the tolerant allele was from a wild soybean accession PI 483463, was recently mapped within a 658-kb region between SSR03_1335 and SSR_1359 on chromosome 310. The 658-kb region contained 80 annotated genes, including two genes (Glyma03g32890 and Glyma03g32900) belonging to the sodium/hydrogen exchanger family. Using the whole-genome sequencing approach, a candidate causal gene, GmCHX1, a counterpart of Glyma03g32900 in Williams 82, was identified in a wild soybean accession W0512. Glyma03g32900 (GmSALT3), which associated with limiting the accumulation of sodium ion, was also identified as a candidate causal gene underlying the QTL on chromosome 3 from a Chinese commercial soybean cultivar, Tiefeng 8, by using fine mapping method20.
Functional analyses of sodium (Na+) and potassium (K+) transporters have been extensively conducted because of their roles in both nutrition and salt tolerance21,22,23,24,25,26,27. Several candidates for the cation antiporter associated with salt tolerance such as GmCHX112, GmSALT320, GmCAX128, GmNHX129, GmPIP30 and GmHKT131 have been reported in soybean. In contrast, mechanisms of chloride (Cl−) transport in plants are poorly understood, despite the importance of minimizing Cl− toxicity for salt tolerance. Control of Cl− transport and Cl− “exclusion” from shoots is correlated with salt tolerance in soybean1,7,11,13,32 and other legumes Trifolium33, Medicago34 and Lotus35 as well as in other crops, such as wheat36; however, no gene that regulates the transport and accumulation of Cl− in soybean has been reported.
Map-based cloning or positional cloning strategies have been successfully used in the isolation of genes underlying QTLs of environmental stress tolerant traits such as salt25, submergence37 and phosphorus deficiency38 tolerances in crops. In this study, we isolated a QTL for salt tolerance using the map-based cloning strategy from a from a Brazilian soybean cultivar FT-Abyara to facilitate its use in breeding. Our results showed that the salt tolerance gene could synchronously regulate the transport and accumulation of Na+, K+ and Cl− in soybean. The near isogenic lines (NILs) carrying the tolerant allele of the QTL could increase soybean grain yield by 3.6–5.5 times in saline field conditions.
Results
Map-based cloning of the salt tolerance QTL
To identify the gene that conditioned the salt tolerance QTL, we initially conducted fine mapping using 1,053 plants derived from 6 F8 residual heterozygous plants and narrowed down the QTL to a 58.8-kb region between simple sequence repeat (SSR) markers BARCSOYSSR_03_1342 and BARCSOYSSR_03_1338 (Fig. 1a). However, this genomic region was not enough for identification of the causal gene underlying the QTL because there were seven predicted genes within this region based on the Glyma 1.0 soybean gene annotation database of the Williams 82 genome sequence39, including two genes (Glyma03g32890 and Glyma03g32900) belonging to the sodium/hydrogen exchanger family. Next, a segregation population consisting of 5,828 plants derived from F9 residual heterozygous plants was used for screening recombination between BARCSOYSSR_03_1342 and BARCSOYSSR_03_1338. The recombination screen facilitated in the selection of 29 plants with homozygous genotype at one marker and heterozygous genotype at the other marker. The selected plants were self-pollinated for one more generation to generate fixed recombination lines. Salt tolerance evaluation and further genotyping of the recombination lines using newly developed DNA markers delimited the QTL to a 16.6-kb interval between SSR markers SSR25.8 and CAPS42.4. Only one predicted gene, Glyma03g32900, existed within the 16.6-kb region (Fig. 1a). RT-PCR analysis showed that the level of transcription of Glyma03g32900 was higher in the salt-tolerant line NILs18-T than in the salt-sensitive line NILs18-S (Fig. 1b). Glyma03g32900 was thus determined as the causal gene underlying the salt tolerance QTL and was named as Ncl followed Abel (1969)13 (previously designated as qNaCl3).

Map based cloning and expression of the salt tolerance gene Ncl.
(a) Fine mapping delimits Ncl to a 16.6-kp region between SSR25.8 and CAPS42.4 on chromosome 3. amarker position (kb), bmarker position (bp). Red and blue bars represent homozygous chromosome segments for tolerance and sensitivity, respectively. Na+, K+ and Cl− leaf contents (n = 3) and shoot dry weight (n = 5–8) for each recombinant line after a treatment with 100 mM NaCl for approximately three weeks in a hydroponic condition. Data are presented as means ± s.d. **significant difference at P < 0.01 level versus NILs18-28-14-S (Dunnett’s multiple comparison test). (b) Expression of Ncl analyzed by semi-quantitative RT-PCR in the roots at one and three days after treatment with 100 mM and 0 mM (Control) NaCl in a hydroponic condition. The actin gene was used as a control.
A Ty1/copia type retrotransposon might cause loss of function of Ncl in soybean
Using the 3′-Full rapid amplification of cDNA ends (RACE) and the 5′-Full RACE methods, full-length cDNAs of the tolerant and the sensitive alleles were determined from the salt-tolerant line NILs18-T and the sensitive line NILs18-S, respectively. On the basis of the positions of start and stop codons, the coding sequence (CDS) of NILs18-T was determined to be 2,436 bp in length and encodes 811 amino acid residues; the CDS of NILs18-S was 1,131 bp in length and encodes 376 amino acid residues. Comparison of genomic and cDNA sequences showed that Ncl harbored five CDSs in the salt-tolerant line NILs18-T, whereas three CDSs were detected in the sensitive line NILs18-S (Supplementary Fig. S1a online). The NILs18-S possessed a 3.8-kb insertion compared with the tolerant line NILs18-T. Because there were long repeated sequences within the 3.8-kb fragment, which suggested that the element belongs to a retrotransposon family, we conducted a BLAST analysis using a soybean transposable element database. We found that the 3.8-kb fragment is probably a Ty1/copia type retrotransposon. Alignment of the full-length cDNA sequences for NILs18-T and NILs18-S showed a point mutant at the 5′-untranslated region (UTR). In the 3′-end of the sensitive line (NILs18-S) cDNA, two polyadenylation signals of AATAAA were detected at 38 bp and 60 bp upstream of the poly (A) tail (Supplementary Fig. S2 online). The polyadenylation signal, which was introduced by the Ty1/copia type retrotransposon, may be responsible for the transcription termination of Ncl in the sensitive line.
The genotype of the soybean root, but not shoot, determines salt tolerance
In this study, Ncl expression was significantly higher in the root tissues compared with the other plant parts (data not shown). To understand the role of the root system in salt tolerance, we grafted the arable part of the salt-tolerant line NILs25-T to the root of the sensitive line NILs25-S, or in reverse, to produce two kinds of graft hybrids: NILs25-T (shoot)/NILs25-S (root) and NILs25-S (shoot)/NILs25-T (root). Salt tolerance evaluation for the two kinds graft hybrids as well as the NILs pair NILs25-T and NILs25-S with 100 mM NaCl in a hydroponic condition revealed that NILs25-T and the graft hybrid NILs25-S/NILs25-T showed higher salt tolerance than NILs25-S and the graft hybrid NILs25-T/NILs25-S in terms of leaf SPAD value, shoot dry weight and Na+, K+ and Cl− concentration in the shoots, indicating that the root genotype, but not the shoot genotype, determines salt tolerance in soybean (Supplementary Fig. S3 and S4 online).
Ncl synchronously regulates the accumulation of Na+, K+ and Cl− in soybean leaves
BLAST analysis showed that the Ncl was homologous to the Na+/H+ antiporter gene family (Supplementary Fig. S1b online). It had regulatory functions for Na+ and K+ transport in soybean plant. However, our analysis observed that Ncl also regulated Cl− accumulation. The salt-tolerant lines showed lower Na+, K+ and Cl− contents in the leaves than the sensitive lines under salt stress conditions (Fig. 1a; Supplementary Fig. S4 online). Another interesting result was that the Na+ and K+ distributions in soybean plants (roots and shoots) were different from that of Cl− under salt stress conditions. In comparison with the salt-sensitive lines, the salt-tolerant line accumulated lower Na+ and K+ concentrations in both shoots (leaves and stems) and roots. In contrast, the salt-tolerant line had lower Cl− concentrations in shoots (leaves and stems) and higher Cl− concentration in root (Supplementary Fig. S4 online). We also tested the three sets of NILs under the stress condition of 100 mM KCl. No significant differences in leaf Cl− content were observed between the tolerant and sensitive lines (Supplementary Fig. S5a online). Both tolerant and sensitive lines showed chlorosis symptom under 100 mM KCl stress condition (Supplementary Fig. S5b online). Ncl functioned like the cation-chloride cotransporter (CCC) genes that were identified in other plants. However, Ncl showed a very low similarity to these putative CCC genes identified in other plant species (Supplementary Fig. S1b online).
Overexpression of Ncl resulted in improved salt tolerance in transgenic soybean
We transformed Ncl full-length cDNA driven with 35S promoter (35S:Ncl) into a Japanese soybean cultivar Kariyutaka using the Agrobacterium-mediated transformation method. We used Kariyutaka for gene transformation because it is highly amenable to Agrobacterium-mediated transformation40 and is a salt-sensitive cultivar with a relatively low level of expression of Ncl and the same gene structure as the sensitive line NILs-18S (this study). We investigated four T2 transgenic lines (54-1-1, 34-2-7, 20-1-4 and 16-1-8), which were derived from independent explants, for their expression of Ncl and salt tolerance performances. All the four T2 lines showed significantly higher levels of expression of Ncl compared with that of the wild-type Kariyutaka and a 35S:GFP transgenic line that was used as control, either in the salt stress of 100 mM NaCl or in the control condition (Fig. 2a; Supplementary Fig. S6a online). A close association between expression of Ncl and salt tolerance was observed (Fig. 2b–e). The 20-1-4 line, which was single copy number for Ncl (Supplementary Fig. S6b online), showed the highest expression level in the four transgenic lines. As expected, the 20-1-4 line showed the highest salt tolerance in terms of leaf SPAD value, shoot dry weight and ions (Na+, K+ and Cl−) concentration in leaves. The results generated by the soybean transgenic line confirmed the function of the Ncl gene in relation to soybean salt tolerance.

Overexpression of Ncl in transgenic lines enhanced salt tolerance.
(a) Real-time quantitative RT-PCR analysis of Ncl expression levels in the transgenic soybean lines and control materials after treatments with 100 mM NaCl for 24 hours in a hydroponic condition. 54-1-1, 34-2-7, 20-1-4 and 16-1-8 are T2 35S:Ncl transgenic lines. Kariyutaka: wild-type soybean cultivar. GFP is 35S:GFP transgenic line (T6). NILs18-T and NILs18-S are tolerant and sensitive near isogenic lines. Values represent means from three biological replicates. (b,c) Leaf SPAD values and shoot dry weight for the transgenic lines after treatments with 100 mM NaCl for approximately three weeks in a hydroponic condition. (d) Na+, K+ and Cl− leaf contents for the transgenic lines after treatments with 100 mM NaCl and 0 mM NaCl (Control) for approximately three weeks in a hydroponic condition. (e) Comparison of salt tolerance between 35S:Ncl plants and null plants of the T2 transgenic line 20-1-4. Data are expressed as means ± s.d. **indicate significant difference at P < 0.01 levels versus from Kariyutaka (wild-type) (Dunnett’s multiple comparison test).
Expression of Ncl was closely associated with salt tolerance in 123 soybean germplasms from different countries
To understand the function of Ncl on salt tolerance in different genetic backgrounds, we analyzed the relationship between Ncl expression level and salt tolerance in 123 soybean germplasms, including the three sets of NILs for salt tolerance and seven wild soybean accessions. The cultivated soybean germplasms were collected from nine countries (Supplementary Table S2 online). The expression levels of Ncl showed a close correlation with salt tolerance in terms of leaf SPAD values (r = 0.7214, P < 0.01) and STR (r = 0.7476, P < 0.01) (Supplementary Fig. S7 online), despites the fact that the mRNA level and protein level were not always associated. This result indicated that Ncl was effective in different genetic backgrounds. Other genes need to be considered to explain the difference of expression level of Ncl among the accession that harbored Ncl.
Introgression of Ncl into the salt-sensitive cultivar Jackson by MAS
To determine the usefulness of Ncl in soybean breeding, we conducted introgression of the salt tolerance allele identified in a wild soybean8 into a salt-sensitive variety, Jackson (PI 548657), through continuous backcross, followed by MAS using primers Satt339, SSR222042 and SSR112166 in each generation. A BC4F2 plant, which was heterozygous for the three SSR markers, was self-pollinated to generate a salt-tolerant homozygous line BC4F3-J1T and a sensitive homozygous line BC4F3-J1S. Salt tolerance evaluation with 100 mM NaCl in a hydroponic condition showed that BC4F3-J1T had significant higher leaf SPAD values and shoot dry weight and lower leaf ion (Na+, K+ and Cl−) contents than BC4F3-J1S as well as Jackson. In contrast, no significant differences in leaf SPAD value, shoot dry weight and leaf ion (Na+, K+ and Cl−) content in the control condition were observed between BC4F3-J1T and BC4F3-J1S (Fig. 3a–d). This result demonstrated that the DNA markers around Ncl could be used for introgression of Ncl into a salt-sensitive cultivar for developing of a soybean variety for high salt tolerance.

Effect of Ncl on salt tolerance in terms of leave SPAD value (a), shoot dry weight (b) and ion (Na+, K+ and Cl− leaf contents (c) in BC4F3 lines developed by MAS by introducing the tolerance allele from a wild soybean accession JWS156-1 into Jackson. BC4F3-J1T and BC4F3-J1S was derived from progeny of self-pollination of a BC4F2 plant, which was heterozygous in the Ncl region. (d) Performance of BC4F3-J1T and BC4F3-J1S after treatment with 100 mM NaCl for approximately three weeks in a hydroponic condition. Data are shown as mean ± s.d. (n = 11). **indicate significant difference at P < 0.01 levels versus Jackson (Dunnett’s multiple comparison test).
The Ncl gene could increase soybean grain yield 3.6–5.5 times in saline field conditions
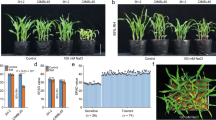
To determine the effect of Ncl in a salt stress field condition and its potential in soybean breeding for salt tolerance, we evaluated the NILs for salt tolerance in a salt stress field condition. The field experiments were conducted for three years. In 2009 and 2011, the three sets of NILs, including a salt-sensitive cultivar, Tachiyutaka, were tested. As a result, all the lines with the Ncl allele showed a significantly higher yield than the lines with the sensitive allele (Fig. 4a–e; Supplementary Fig. S8 online). In the 2011 field experiment, the average yield of the three tolerant lines (NILs18-T, NILs25-T and NILs72-T) was 2.383 t/h, whereas the average yield of the three sensitive lines (NILs18-S, NILs25-S and NILs72-S) was 0.437 t/h, increasing the grain yield by 5.5 times in a saline field condition (Fig. 4c). In 2012, we tested another six fixed lines derived from an F8 residual heterozygous line for the QTL region. These six lines have the same genetic background but different chromosome length for the salt tolerance QTL region. Three (N18-39, N18-99 and N18-122) of these lines were salt-tolerant, whereas three lines (N18-9, N18-61, N18-180) were salt-sensitive. The results of field experiment showed that the average yield for the three tolerant lines was 2.973 t/h, whereas that for the three sensitive lines was 0.828 t/h, indicating an increase in the grain yield by 3.6-fold in a saline field condition (Fig. 4d). In contrast, no significant differences in grain yield were observed between the tolerant and sensitive NILs in a control condition (Fig. 4c,d). Salt stress reduced the grain yield for all the lines by 55.8% in 2011 and 44.0% in 2012. For the lines with the Ncl allele, the yield decreased on average by 26.7% in 2011 and 29.5% in 2012, whereas those without the Ncl allele were 86.0% in 2011 and 76.5% in 2012. These results clearly indicated that Ncl contributed to achieving sustainable soybean production in a salt stress condition.

Yield performances of lines carrying Ncl in a saline field condition in Miyagi Prefecture, Japan.
(a) Top view of NILs25-S and NILs25 T grown in a salt stress field in 2009. (b) Top view of N18-61 and N18-99 grown in a salt stress field in 2012. (c) Grain yield result of the three sets of NILs in 2011 field test. (d) Grain yield result of six recombinant lines derived from an F8 residual heterozygous line (RHL18-28) in 2012 field test. N18-39, N18-99 and N18-122 are lines carrying tolerant allele of Ncl, whereas N18-9, N18-61, N18-180 are lines that had the sensitive allele. (e) Na+, K+ and Cl− leaf contents for the three sets of NILs in 2011 field test. Data are shown as mean ± s.d. from three replicates. **Significant difference (P < 0.01) based on ANOVA (Tukey’s multiple comparison test).
Discussion
Previous studies on plant salt tolerance mainly focused on the toxic effects of Na+ or the balance of Na+ and K+ 21,22,23,24,25,26,27. Because BLAST homology search showed that Ncl belonged to the Na+/H+ antiporter gene family, we expected to observe Na+ and K+ regulation by Ncl as observed in the studies of Qi et al.12 and Guan et al.20. However, the present study not only observed this regulatory event in soybean, but also the controlled accumulation of Cl−. The similar results for regulation of Cl− accumulation were observed in the fine-mapping, grafting, transgenesis, gene introgression by marker-assistant selection (MAS) and field testing experiments. Previous reports have described the high accumulation of Cl− in soybean when subjected to salt stress1,7,11,13,31; however, no gene that regulates the uptake, transport and accumulation of Cl− in soybean has been identified. The CCC gene has been well studied in animals41,42; however, information on this gene in various plant species is limited. An OsCCC1 gene that regulates K+ and Cl− but not Na+ was reported in rice43. An AtCCC gene was cloned in Arabidopsis thaliana and its protein operated as a Na+K+Cl− co-transporter44, providing evidence for the occurrence of cation-Cl− cotransport in the plant kingdom. However, Ncl showed very low level of similarity to these putative CCC genes. Although BLAST homology search showed that Ncl belonged to the Na+/H+ antiporter gene family, only the first half part of Ncl (around 1–400 aa) showed homology with Na+/H+ antiporter gene. As for the remaining half part, no significant homologous hit was detected. The function of this region remains unclear. The fact that Ncl synchronously regulates the transport and accumulation of Na+, K+ and Cl− suggests that it may has salt tolerance function differing from the previous salt tolerance genes identified in plant.
Transgenic line 20-4-1 showed a significantly higher Ncl expression compared with that of the wild type Kariyutaka as well as that of the salt-tolerant line NILs18-T. This feature resulted in the transgenic line 20-4-1 having significantly higher salt tolerance compared with those not only in the wild-type cultivar Kariyutaka but also in the tolerant line NILs18-T. This result implied that overexpression of Ncl may generate a higher salt tolerance line than the current available salt tolerant soybean germplasm. To determine the potential of the Ncl transgenic lines in practical breeding, further studies on possible negative effects in the Ncl-overexpressing transgenic line and its salt tolerance performance in more severe salt stress conditions are needed.
A wild soybean accession W05, from which a salt-tolerant gene (GmCHX1) was identified, was proposed to be used be used for soybean breeding for salt tolerance12. Guan et al. also identified the same salt-tolerant gene (GmSALT3) from a Chinese commercial cultivar Tiefeng 820. The present study detected many wild and cultivated soybean accessions that harbored the tolerant allele of Ncl. Some cultivated soybean cultivars that highly express Ncl and show a greater degree of salt tolerance, such as S-100 and FT-Abyrara, were identified. The fact that both wild and cultivated soybeans showed variations in salt tolerance suggested that the loss-of-function mutation in Ncl occurred before the domestication of soybean from its ancestral wild species. The close association between Ncl expression and salt tolerance also demonstrated that Ncl is effective in different genetic backgrounds. In the present study, we transgressed the salt tolerance allele from the wild soybean accession, JWS156-1, to a cultivated soybean cultivar, Jackson. After four cycles of backcrossing, the effect of the salt tolerance gene was confirmed. However, the BC4F3 introgression lines still generated a lower biomass yield compared with its recurrent parent, Jackson, in a control condition. Therefore, the cultivated soybean germplasm may be more efficiently used in breeding cultivars with improved salt tolerance compared with that of wild soybean, which generally possesses several undesirable agronomic traits.
Evaluation of the effect of a salt tolerance gene on grain yield is extremely difficult when conducted in a nature field condition. One reason is that the concentration of salt is generally uneven both horizontally and vertically in the field. Moreover, it is influenced by several environmental factors such as rainfall. In the present study, field experiments were conducted in a paddy field, which was irrigated with diluted seawater. This method provided a uniform salt stress for each soybean line. Moreover, salt tolerance NILs were used to study the effect of the salt tolerance gene. NILs have almost the same genetic background, which in turn can rule out the influences of other traits such as flowering time, maturity and plant size on salt evaluation, thus facilitating in determining the actual effect of the particular gene on yield. The results of the field experiments showed that the Ncl gene could increase soybean grain yield by 3.6–5.5-fold in a saline field condition. To confirm the general effect of Ncl, field tests for salt tolerance of the NILs are currently in progress in China, India and Indonesia. The multi-environmental tests for these materials will enable us to understand the effect of Ncl in these regions using different concentration or kinds of salt soils such as the sodic soil. In addition, introgression of the Ncl into local commercial soybean varieties is presently being conducted in these countries. Isolation and characterization of Ncl may contribute to the sustainability of soybean production in saline areas by introducing Ncl into the local soybean varieties either by MAS or gene transformation methods.
Methods
Map-based cloning of the salt tolerance QTL
A population comprising of 1,053 plants derived from 6 F8 residual heterozygous plants was initially used for fine mapping of the salt tolerance QTL. These residual heterozygous plants were selected from a RIL population derived from a cross between FT-Abyara and C011. They were heterozygous at the QTL region between SSR markers Satt255 and Sat_091 on chromosome 3. Sixteen plants that were homozygous at one marker and heterozygous at another marker were selected. The selected plants were self-pollinated for one more generation to produce 16 fixed recombination lines. Salt tolerance evaluation for the 16 recombination lines using 100 mM of NaCl in a hydroponic condition and DNA marker analysis determined that the QTL was located within a 58.8-kb region between SSR markers BARCSOYSSR_03_1342 and BARCSOYSSR_03_1338. To determine the causal gene conferring salt tolerance at this major QTL, a segregation population consisting of 5,828 plants derived from 32 F9 residual heterozygous plants was used for screening recombination between BARCSOYSSR_03_1342 and BARCSOYSSR_03_1338. From this population, 29 plants with the homozygous genotype for one marker and heterozygous for the other marker were selected. The selected plants were self-pollinated for one more generation to produce fixed recombination lines. On the basis of the soybean genome sequence information, six DNA markers (Supplementary Table S1 online) were developed between BARCSOYSSR_03_1342 and BARCSOYSSR_03_1338. Salt tolerance evaluation and further genotyping of the recombination lines with the newly developed DNA markers were used for high-resolution mapping.
Salt tolerance evaluation using hydroponics in the greenhouse
Salt tolerance evaluation of soybean genotypes was performed using hydroponics under greenhouse conditions at the Japan International Research Center for Agricultural Sciences, Japan, according the method described previously19. After around three weeks of salt treatment with 100 mM of NaCl or KCl, salt tolerance rating (STR), leaf SPAD, dry weight and ion (Na+, K+ and Cl−) contents were measured for each genotype. The STR scale was classified into five grades, ranging from 1 to 5 (1, plants completely dead; 2, two-third or more leaves showed chlorosis symptoms or only upside leafs survive; 3, half or less leaves showed chlorosis symptoms; 4, one-third or less leaves showed chlorosis symptoms; 5, plants bearing normal, healthy leaves). The SPAD value for each genotype was measured using a chlorophyll meter (Konica Minolta SPAD-502, Tokyo, Japan). This SPAD value is proportional to the chlorophyll content in the leaves.
RNA isolation and expression analysis
Soybean plant tissues were collected from different soybean genotypes, frozen in liquid nitrogen and stored at −80 °C. Total RNA was extracted using Plant RNA extraction Kit (QIAGEN, Japan), according to the manufacturer’s protocol. The first strand cDNA was synthesized using the First Strand cDNA Synthesis Kit (Toyobo, Osaka, Japan). RT-PCR was performed using QuantiTect Rev. Transcription Kit (QIAGEN, Tokyo, Japan) with primers of Glyma03g32900_CDS6-F and Glyma03g32900_CDS6-R (Table S1 online). The reaction program was 30 s at 94 °C, 30 s at 56 °C and 1 min at 72 °C for 32 cycles and a final extension of 7 min at 72 °C. The soybean actin gene was used as reference gene in the RT-PCR analysis (GenBank Acc. No. V00450). All PCR products were separated on 1.0% agarose gels and visualized with Pharos Molecular Imager (BIO-RAD, Hercules, CA, USA). Real-time PCR was performed using SsoFastTM EvaGreen® Supermix (BIO-RAD, Hercules, CA, USA) with the same primers used for RT-PCR. All the reactions were performed in a Bio-Rad CFX96TM machine (BIO-RAD, Hercules, CA, USA). The soybean actin was used as reference gene. The reaction included an initial 30 s denaturation step at 95 °C, followed by 5 s at 95 °C and 5 s at 55 °C for 40 cycles. The PCR products were identified by melting curve analysis conducted over the range of 65–95 °C at the end of each PCR run. The 2−ΔΔCt method was used to normalize the relative gene expression data in the qPCR assay45.
Obtaining of full-length cDNA sequences in salt-tolerant and -sensitive lines using Rapid amplification of cDNA ends
Salt tolerance near isogenic lines NILs18-T (tolerant) and NILs18-S (sensitive)19 were used to generate full-length cDNA sequences of the salt tolerance gene Ncl. 3′-Full Rapid amplification of cDNA ends (RACE) and 5′-Full RACE were performed to obtain 3′- and 5′-UTR using the total RNAs extracted from NILs18-T and NILs18-S, respectively. The 3′-Full RACE was conducted using a 3′-Full RACE Core Set (TaKaRa, Otsu, Japan), following the manufacturer’s protocol. Instead of using oligo (dT) primers, an adaptor primer was used to synthesize cDNA. An upstream primer was designed using the Glyma03g32900 gene as reference to amplify 3′ RACE cDNA (Table 1). Five μl of 3′ RACE-ready cDNA was used as template in a 20-μl PCR reaction. The RT-PCR conditions included 32 cycles for 30 s at 94 °C, 30 s at 56 °C and 2.5 min at 72 °C, followed by an extension of 7 min at 72 °C. The 5′-Full RACE was performed using 5′-Full RACE Core Set (TaKaRa, Otsu, Japan), following the manufacture’s protocol except for the 5′ End-phosphorylated reversed transcription primer, 1st primer pair and 2nd primer pair (Supplementary Table S1 online). 5′ RACE cDNA synthesis was performed in 15-μl cDNA synthesis reactions containing 2.5 μg of total RNA. The 1st PCR reactant was diluted 10-fold and used as template in the 2nd PCR reaction. The PCR conditions included 30 cycles for 30 s at 94 °C, 30 s at 56 °C and 30 s at 72 °C, followed by an extension of 7 min at 72 °C. The PCR products from the 3′-Full RACE-PCR and 5′-Full RACE-PCR were purified from the agarose gel using Winzard SV Gel and PCR Clean-Up System (Promega Corporation, Madison, WI, USA) according to the manufacturer’s protocol. The purified DNAs were subjected to sequence analysis.
Soybean grafting experiment
For the soybean grafting experiment, salt tolerance near isogenic lines NILs25-T (tolerant) and NILs25-S (sensitive) were used19. Soybean seeds were sown in vermiculite and germinated in the greenhouse with day and night temperatures at 25 °C/20 °C. One week after emergence, the plants were transferred to a hydroponic culture (half-strength Hoagland’s and Arnon solutions). When the seedlings had developed unifoliate leaves, NILs25-T and NILs25-S were grafted onto each other as scion and rootstock. This procedure resulted in two scion/rootstock combinations: NILs25-T (shoot)/NILs25-S (root) and NILs25-S (shoot)/NILs25-T (root). Ten days after grafting, 12 plants from each grafted hybrid and the two near isogenic lines NILs25-T and NILs25-S were evaluated in hydroponic culture with the half-strength Hoagland’s and Arnon solutions containing 100 mM of NaCl.
Measurements of ion (Na+, K+ and Cl−) contents in soybean plants
Harvested plants were washed with deionized water. Samples were oven-dried for 3 days at 60–70 °C and dry weight were measured. For the analysis of Na+, K+ and Cl− ion contents, dried plant samples were ground using mixer mill (Millser IFM-700G, Iwatani, Japan). Na+and K+ ion contents were determined using atomic absorption spectrometry, (Z-5010, Hitachi, Japan) after dry ashing procedure and digestion with 2 N hydrochloric acid. Cl− ion content was determined using the Chloride Assay Kit (BioChain, Newark, CA, USA) after 2 days of extraction with Milli-Q water at 4 °C.
Analysis of the association between expression level of Ncl and salt tolerance in 123 soybean germplasms from different countries
A total of 123 soybean accessions (117 cultivated soybean accessions and six wild soybean accessions) were used to evaluate the salt tolerance and to investigate the expression level of the Ncl gene (Supplementary Table S2 online). Evaluation of the salt tolerance was performed using hydroponic culture in the greenhouse. RNA samples were collected from three plants from each line after 1 day of treatment with 100 mM NaCl in a hydroponic condition for real-time quantitative PCR analysis of Ncl.
Generation of Ncl transgenic soybean
To generate transgenic soybean overexpressing Ncl, we transformed Ncl using the Agrobacterium-mediated transformation method into a Japanese soybean cultivar Kariyutaka. Kariyutaka was employed for gene transformation because it is highly amenable to Agrobacterium-mediated transformation40; this cultivar is also salt-sensitive, with a relatively low level of expression of Ncl (this study). The full-length cDNA of Ncl was cloned into the pMDC123 vector between the CaMV 35S promoter and the AtHSP terminator, thus obtaining the pMDC123–35S–Ncl–AtHSP terminator vector with the Bar gene. The transformation followed a previously described procedure40. Four T2 transgenic soybean lines (20-1-4, 54-1-3, 34-2-7 and 16-1-8) were subjected for salt tolerance evaluation with hydroponic in the green house.
Introgression of Ncl into salt-sensitive cultivar by DNA MAS
A salt sensitive soybean cultivar Jackson (PI 548657) was crossed with a salt-tolerant wild soybean accession, JWS156-1, to produce F2 plants and subsequent backcrosses with Jackson. DNA MAS was performed using SSR markers Satt339, SSR222042 and SSR112166 in each generation (Supplementary Table S1 online). A BC4F2 plant, which was heterozygous for the SSR markers, was self-pollinated to generate a salt tolerance homozygous line BC4F3-–J1T and a sensitive homozygous line BC4F3-J1S. These two lines, which had similar genetic backgrounds but were different for the salt tolerance QTL allele, as well as the recurrent parent cultivar Jackson, were subjected to salt tolerance evaluation using hydroponic.
Testing the effect of Ncl on soybean grain yield in saline field conditions
To investigate the effect of Ncl on soybean grain yield in response to salt stress, we grew the Ncl near isogenic lines in a saline field condition at the farm of Graduate School of Life Sciences, Tohoku University (38°27′ N, 141°05′ E) located in Miyagi Prefecture, Japan, for three years, namely, 2009, 2011, 2012. In 2009 and 2011, the three pairs of near isogenic lines (NILs18-T, NILs18-S, NILs25-T, NILs25-S, NILs72-T, NILs72-S), as well as a local soybean cultivar, Tachiyutaka, were tested. In 2012, we tested another six fixed-recombination lines derived from an F8 residual heterozygous line RHL18 for the QTL region. These six lines have the same genetic background, but had different lengths for the chromosome segment of the salt tolerance QTL region. Three (N18-39, N18-99 and N18-122) of these lines were tolerant and three lines (N18-9, N18-61, N18-180) were sensitive. The experiment was laid out in a randomized block design with three replicates. The areas of each plot were 6.7, 13.76 and 8.0 m2 in 2009, 2011 and 2012, respectively, with two rows in which soybean plants were plated at 20-cm intervals between hills and 80 cm between rows. Salt stress treatments were conducted at around 5 weeks after sowing by irrigated with around 1/4 concentration of seawater and normal water (Control). Leaf samples from five plants of each line were taken after four weeks of treatment for ion (Na+, K+ and Cl−) in 2011. After maturity, we randomly sampled 10 plants from each plots and harvested the remains to measure grain yield.
Additional Information
How to cite this article: Do, T. D. et al. Ncl Synchronously Regulates Na+, K+ and Cl– in Soybean and Greatly Increases the Grain Yield in Saline Field Conditions. Sci. Rep. 6, 19147; doi: 10.1038/srep19147 (2016).
References
Abel, G. H. & MacKenzie, A. J. Salt tolerance of soybean varieties (Glycine max L. Merill) during germination and later growth. Crop Sci. 4, 157–161 (1964).
Wang, D. & Shannon, M. C. Emergence and seedling growth of soybean cultivars and maturity groups under salinity. Plant Soil 214, 117–124 (1999).
Singleton, P. W. & Bohlool, B. B. Effect of salinity on nodule formation by soybean. Plant Physiol. 74, 72–76 (1984).
Parker, M. B., Gascho, G. J. & Gains, T. P. Chloride toxicity of soybeans grown on Atlantic Coast flatwoods soils. Agron. J. 75, 439–443 (1983).
Yang, J. & Blanchar, R. W. Differentiating chloride susceptibility in soybean cultivars. Agron. J. 85, 880–885 (1993).
Lee, J. D. et al. Evaluation of a simple method to screen soybean genotypes for salt tolerance. Crop Sci. 48, 2194–2200 (2008).
Luo, Q., Yu, B. & Liu, Y. Differential sensitivity to chloride and sodium ions in seedlings of Glycine max and G. soja under NaCl stress. J. Plant Physiol. 162, 1003–1012 (2005).
Hamwieh, A. & Xu, D. H. Conserved salt tolerance quantitative trait locus (QTL) in wild and cultivated soybeans. Breeding Sci. 58, 355–359 (2008).
Lee, J. D., Shannon, J. G., Vuong, T. D. & Nguyen, H. T. Inheritance of salt tolerance in wild soybean (Glycine soja Sieb. and Zucc.) accession PI483463. J. Hered. 100, 798–801 (2009).
Ha, B. K. et al. Genetic mapping of quantitative trait loci conditioning salt tolerance in wild soybean (Glycine soja) PI 483463. Euphytica 193, 79–88 (2013).
Chen, P., Yan, K., Shao, H. & Zhao, S. Physiological mechanisms for high salt tolerance in wild Soybean (Glycine soja) from Yellow River Delta, China: photosynthesis, osmotic Regulation, ion flux and antioxidant capacity. PLoS ONE 8, e83227 (2013).
Qi, X. et al. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 5, 4340 (2014).
Abel, G. H. Inheritance of the capacity for chloride inclusion and chloride exclusion by soybeans. Crop Sci. 9, 697–698 (1969).
Shao, G. H., Chang, R. Z., Chen, Y. W. & Yan, S. R. Study on inheritance of salt tolerance in soybean. Acta Agronomica Sinica 20, 721–726 (1994).
Song, Q. J. et al. A new integrated genetic linkage map of the soybean. Theor. Appl. Genet. 109, 122–128 (2004).
Song, Q. J. et al. Abundance of SSR motifs and development of candidate polymorphic SSR markers (BARCSOYSSR_1.0) in soybean. Crop Sci. 50, 1950–1960 (2010).
Lee, G. J. et al. A major QTL conditioning salt tolerance in S-100 soybean and descendent cultivars. Theor. Appl. Genet. 109, 1610–1619 (2004).
Chen, H., Cui, S., Fu, S., Gai, J. & Yu, D. Identification of quantitative trait loci associated with salt tolerance during seedling growth in soybean (Glycine max L.). Aust. J. Agr. Res. 59, 1086–1091 (2008).
Hamwieh, A. et al. identification and validation of a major QTL for salt tolerance in soybean. Euphytica 79, 451–459 (2011).
Guan, R. et al. Salinity tolerance in soybean is modulated by natural variation in GmSALT3. Plant J. 80, 937–950 (2014).
Apse, M. P., Aharon, G. S., Snedden, W. A. & Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285, 1256–1258 (1999).
Shi, H., Ishitani, M., Kim, C. & Zhu, J. K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. 97, 896–6901 (2000).
Apse, M. P., Sottosanto, J. B. & Blumwald, E. Vacuolar cation/H+ exchange, ion homeostasis and leaf development are altered in a T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter. Plant J. 36, 229–239 (2003).
Shi, H., Lee, B. H., Wu, S. J. & Zhu, J. K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 21, 81–85 (2003).
Ren, Z. H. et al. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 37, 1141–1146 (2005).
Munns, R. et al. Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat. Biotechnol. 30, 360–364 (2012).
Byrt, C. et al. The Na+ transporter, TaHKT1; 5-D, limits shoot Na+ accumulation in bread wheat. Plant J. 80, 516–526 (2014).
Luo, G. Z. et al. A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Mol. Biol. 59, 809–820 (2005)
Sun, Y. X., Wang, D., Bai, Y. L., Wang, N. N. & Wang, Y. Studies on the overexpression of the soybean GmNHX1 in Lotus corniculatus: the reduced Na+ level is the basis of the increased salt tolerance. Chin. Sci. Bull. 51, 1306–1315 (2006).
Li, L. et al. Cloning and function analysis of a plasma membrane intrinsic protein gene, GmPIP in soybean (Glycine max L. Merr). Aust. J. Agr. Res. 8, 738–746 (2014)
Chen, H., He, H. & Yu, D. Overexpression of a novel soybean gene modulating Na+ and K+ transport enhances salt tolerance in transgenic tobacco plants. Physiol. Plant 141, 11–18 (2011)
Zhang, X. K., Zhou, Q. H., Cao, J. H. & Yu, B. J. Differential Cl−/salt tolerance and NaCl-induced alternations of tissue and cellular ion fluxes in Glycine max, Glycine soja and their hybrid seedlings. J. Agron. Crop Sci. 197, 329–339 (2011).
Rogers, M. E., Noble, C. L., Halloran, G. M. & Nicolas, M. E. Selecting for salt tolerance in white clover (Trifolium repens): chloride ion exclusion and its heritability. New Phytol. 135, 645–654 (1997)
Sibole, J. V., Cabot, C., Poschenrieder, C. & Barcelo, J. Efficient leaf ion partitioning, an overriding condition for abscisic acid–controlled stomatal and leaf growth responses to NaCl salinization in two legumes. J. Exp. Bot. 54, 2111–2119 (2003).
Teakle, N., Flowers, T., Real, D. & Colmer, T. Lotus tenuis tolerates the interactive effects of salinity and waterlogging by ‘excluding’ Na+ and Cl− from the xylem. J. Exp. Bot. 58, 2169–2180 (2007).
Genc, Y., Taylor, J., Rongala, J. & Oldach, K. A major locus for chloride accumulation on chromosome 5A in bread wheat. PLoS ONE 9, e98845 (2014).
Xu, K. et al. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 442, 705–708 (2006).
Gamuyao, R. et al. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 488, 535–539 (2012).
Schmutz, J. et al. Genome sequence of the palaeopolyploid soybean. Nature 463, 178–183 (2010).
Yamada, T., Watanabe, S., Arai, M., Harada, K. & Kitamura, K. Cotyledonary node pre-wounding with a micro-brush increased frequency of Agrobacterium-mediated transformation in soybean. Plant Biotechnology 27, 217–220 (2010).
Delpire, E. & Mount, D. B. Human and murine phenotypes associated with defects in cation–chloride cotransport. Ann. Rev. Physiol. 64, 803–843 (2002).
Gamba, G. Molecular physiology and pathophysiology of electroneutral cation–chloride cotransporters. Physiol. Rev. 85, 423–493 (2005).
Kong, X. Q. et al. Cloning and functional characterization of a cation–chloride cotransporter gene OsCCC1. Plant Mol. Biol. 75, 567–578 (2011).
Colmenero-Flores, J. M. et al. Identification and functional characterization of cation–chloride cotransporters in plants. Plant J. 50, 278–292 (2007).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 25, 402–408 (2001).
Acknowledgements
We thank Drs. A. Kanazawa, Z. Xia, R. Takahashi and Ms. E. Hanzawa for support and suggestions. The work was supported in part by the Grant-in-Aid from the Japan Society for the Promotion of Science (No. 26450013).
Author information
Authors and Affiliations
Contributions
T.D.D., H.C. and H.T.T.V. cloned the Ncl gene and performed the experiments for introgression of Ncl into salt-sensitive cultivar by DNA MAS. A.H. and Y.Y. performed fine-mapping of the salt tolerance QTL. T.Y. transformed the Ncl gene into Kariyutaka. T.S. performed the field experiments. M.S. analyzed ions contents and the performed soybean grafting experiments. H.C., K.S. and D.X. designed the experiments. D.X. wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Do, T., Chen, H., Hien, V. et al. Ncl Synchronously Regulates Na+, K+ and Cl− in Soybean and Greatly Increases the Grain Yield in Saline Field Conditions. Sci Rep 6, 19147 (2016). https://doi.org/10.1038/srep19147
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep19147
This article is cited by
-
Identification of the domestication gene GmCYP82C4 underlying the major quantitative trait locus for the seed weight in soybean
Theoretical and Applied Genetics (2024)
-
Twenty years of mining salt tolerance genes in soybean
Molecular Breeding (2023)
-
Morpho-physiological and genetic characteristics of a salt-tolerant mutant line in soybean (Glycine max L.)
Theoretical and Applied Genetics (2023)
-
Time-series transcriptome comparison reveals the gene regulation network under salt stress in soybean (Glycine max) roots
BMC Plant Biology (2022)
-
Identification of novel genetic sources for agronomic and quality traits in soybean using multi-trait allele specific genic marker assays
Journal of Plant Biochemistry and Biotechnology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
