
Abstract
Decomposition of soil organic matter (SOM) plays an important role in the global carbon cycle because the CO2 emitted from soil respiration is an important source of atmospheric CO2. Carbon isotopic fractionation occurs during SOM decomposition, which leads to 12C to enrich in the released CO2 while 13C to enrich in the residual SOM. Understanding the isotope fractionation has been demonstrated to be helpful for studying the global carbon cycle. Soil and litter samples were collected from soil profiles at 27 different sites located along a vertical transect from 1200 to 4500 m above sea level (a.s.l.) in the south-eastern side of the Tibetan Plateau. Their carbon isotope ratios, C and N concentrations were measured. In addition, fiber and lignin in litter samples were also analyzed. Carbon isotope fractionation factor (α) during SOM decomposition was estimated indirectly as the slope of the relationship between carbon isotope ratios of SOM and soil C concentrations. This study shows that litter quality and soil water play a significant role in isotope fractionation during SOM decomposition and the carbon isotope fractionation factor, α, increases with litter quality and soil water content. However, we found that temperature had no significant impact on the α variance.
Similar content being viewed by others

Introduction
Soil organic carbon is the largest pool of terrestrial ecosystem and greatly affects global carbon cycling. The CO2 derived from soil organic matter (SOM) decomposition is an important source of atmospheric CO2. The CO2 released from soil respiration enriches 12C while the residual SOM enriches 13C, relative to the substrate, because of carbon isotopic fractionation during SOM decomposition1,2,3,4,5,6,7. Consequently, the isotopic fractionation affects the carbon isotope composition of atmospheric CO2 because the released CO2 finally goes into the atmosphere. Scientists who study global change incorporate carbon isotope data for tropospheric CO2, derived from an international network of stations, into atmospheric circulation models. This step is used to calculate global carbon balance and to analyze atmospheric carbon source/sink positions and quantities8,9,10,11. Thus, understanding carbon isotope fractionation during SOM decomposition can help scientists use carbon isotope data for atmospheric CO2 in their studies of global carbon cycling. In addition, it has been demonstrated that adding soil carbon isotope variations to carbon-dynamic models provides tighter constraints on certain model parameters having biological and environmental significance12,13. An understanding of carbon isotope fractionation during SOM decomposition can also enhance reconstructions of past environments. This can benefit studies of C4-plant origins and expansions in geological time that use carbon isotope records of ancient terrestrial sediments6,14,15,16,17,18.
It is well known that environmental and biotic factors affect the decomposition rate of organic matter13,19,20,21,22. Substrate quality q is the dominant biotic influential factor12,13,23,24. Substrate quality quantifies how easily organic carbon is used by soil microbes12,13. It can be related to plant type and is often defined using a C/N ratio, lignin content, cellulose content, and/or lignin content/N ratio25,26. The influences of environmental factors and substrate quality on decomposition have been assessed intensively27,28,29,30,31,32,33,34. However, few studies have focused on whether these factors influence carbon isotope fractionation during SOM decomposition3,12,13,25,35. Therefore, current study was undertaken to evaluate the effects of environmental and biotic factors on carbon isotope fractionation during SOM decomposition. In this regard we measured the carbon isotope ratios of soil and litter samples collected from soil profiles along an altitudinal gradient in the south-eastern side of the Tibetan Plateau. Furthermore, their carbon isotope fractionation (α) during SOM decomposition was estimated using an indirect method established by Poage and Feng (2004)13.
Results
Variations in C/N, lignin and cellulose of litters with altitude
The C/N ratio of litters varied greatly and ranged from 8 to 109 (Fig. 1a). Since litters collected in this study were mainly composed of leaves in situ, the C/N ratio of litter depended on the vegetation types and plant species. The C/N ratio increased up to an elevation of 3600 m and then decreased at higher elevations (Fig. 1a). This observation indicates that the coniferous trees (mainly located at 2800–3600 m) had the highest C/N ratios and the broad-leaved trees (mainly located at 1600–2800 m) and shrubs (mainly located at 1600–2800 m and 3600–4200 m) had the second highest C/N ratios. In addition, the herbaceous plants, including grasses, which were mainly composed of C4 species (located below 1600 m) and alpine frigid meadow vegetation (located at 4200–4600 m) had the lowest C/N ratios.

shows the variations in C/N ratios (a), lignin contents (b) and cellulose contents (c) of litter with altitude.
The woody plants including trees and the shrubs had much higher lignin contents than the herbaceous plants (Fig. 1b). This altitudinal pattern of lignin arose because herbaceous plants are mainly distributed at elevations less than 1600 m and in the range of 4200–4600 m. However, the latter had higher cellulose contents than the former (Fig. 1c). Correlation analysis shows that the C/N ratio of litter is significantly and positively related to the lignin content (p = 0.000) and negatively related to cellulose content of litter (p = 0.008). In addition, a significant negative correlation exists between lignin and cellulose (p = 0.000).
Influences of environmental and biotic factors on fractionation factor α
In this study, only the 60 soil profiles above 1800 m show trends of decreasing C concentration and increasing δ13CSOM with soil depth. Figure 2a shows an example. These profiles were selected for calculation of fractionation factor α. The 15 profiles below 1800 m were omitted because the typical pattern of δ13CSOM increase with depth was not regularly observed in these soil profiles (Fig. 2b). This indicated that the ecosystems below 1800 m experienced C3 to C4 succession, or vice versa36,37. The fractionation factor α of studied samples ranged from 1.00051 to 1.00183 with an average of 1.00118. About 80% of these samples fell between 1.0007 and 1.0017. The ε value was between −0.51 and −1.83 with an average of −1.185. Approximately 80% of sample values for ε fell between −0.75 and −1.7.

shows the vertical changes of δ13CSOM (closed cycles) and organic carbon concentration (open cycles).
The soil profiles in Fig. 2(a) and (b) locate at 2700 m a.s.l. and 1700 m a.s.l. on the eastern slope, Mount Gongga. Note that the organic carbon concentration and δ13C values at depth zero in the figures are from L1 litters.
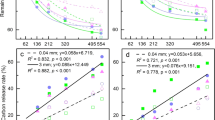
Initially, a series of bivariate correlation analyses were conducted to preliminarily discover the effects of different environmental and biotic factors on fractionation factor α. Figure 3 showed that water contents of 0–5 cm soil layer (W1) exerted positive impact on α (p = 0.006), while C/N and lignin content showed negative influences (p = 0.001 and = 0.004, respectively). The effects of cellulose and water contents of 5–10 cm soil layer (W2) were marginally significant (p = 0.063 and = 0.059, respectively) with positive coefficients. No significant relationship existed between α and mean annual temperature (MAT) and mean summer temperature (MST) (both p = 0.3). Afterwards, a multiple regression of α against the seven factors including C/N ratio, lignin, cellulose, W1, W2, MAT and MST was conducted using ordinary least square (OSL) estimation. This was done to find out how much variances in fractionation factor α were explained by these variables. Although the overall regression is highly significant (p = 0.000, n = 60), the 7 variables just account for 36% (R2) of the total variance. Considering that there are close relationships (collinearity) among variables, for examples, C/N ratio vs. lignin content (p < 0.001), W1 vs. W2 (p < 0.001) and MAT vs. MST (p < 0.001), a stepwise regression (α vs. C/N ratio, lignin, cellulose, W1, W2, MAT and MST) was run with forward direction (criteria: probability of F to enter r ≤ 0.05, probability of F to remove r ≥ 0.100), The result showed that only two variables, C/N and W1 entered the models, suggesting that they were the most significant influential factors. This regression also shows that C/N ratio of litter displayed a greater effect on α than W1 because C/N ratio entered the model earlier than W1.

shows the relationships between α and lg(C/N), lignin content, cellulose content and soil water of 0–5 cm layer.
OLS estimation is based on the data with characteristics of normal distribution and can describe the influence of independent variables on the mean dependent variable. For the data with characteristics of non-normal distribution, OLS no longer has the advantage of the best linear unbiased estimation (BLUE). Consequently, it is not able to effectively describe the influence of independent variables on the minimum and maximum of range of dependent variables. Instead, quantile regression overcomes the limitations of the OSL estimation and can more precisely reflect the effects of independent variables on dependent variables at different quantiles. Thus, in this study quantile regressions were run to exam the dependence of α on environmental and biotic factors in detail. It was also based on the consideration of close relationships (collinearity) among variables, in the quantile regression only three variables, C/N, W1 and MAT were taken as the predictors. The results showed the superiority of the quantile regression (Table 1). This provided more information about the influence of environmental and biotic factors on fractionation factor α compared to the multiple regression based on OSL estimation. For example, the multiple regression shows that W1 exerted a significant impact on α variance; but the quantile regression pointed out that the significant impact did not occur at quantiles 75 and 90. In addition, the multiple regression shows that fractionation factor α was independent of MAT; however, the quantile regression demonstrates that it is not always the case. For example, MAT played a significant role in fractionation factor α at quantile 10. Furthermore, although the influence of MAT on α was slight in the most cases, the influence changed with quantiles. MAT’s impact was positive at quantiles 10 50, 75 and 90 and negative at quantiles 25 (Table 1).
Discussion
This study showed that α values tend to decrease with higher C/N ratios, higher lignin contents and lower cellulose contents. This suggested that an increase in α is associated with higher litter quality (lower C/N ratios, lower lignin content and higher cellulose content). Although at present no direct observation of a correlation between α and litter quality has been reported, two previous studies may indirectly support our finding. Ågren et al. (1996)3 compiled the data from Balesdent et al. (1993)2 and observed that less 13C enrichment occurred in soil where evergreen trees were located, which presumably had a lower litter quality. They further reported that a great 13C enrichment occurred in deciduous forests, which were expected to have a higher litter quality. Garten et al. (2000)25 measured δ13CSOM in the Southern Appalachian Mountains, USA and found that less vertical change in δ13CSOM was associated with poorer litter quality (higher C/N ratios). The 13C enrichment and fractionation factor are not the same parameter, but generally, 13C enrichment is positively proportional to α. Thus, the two previous investigations also suggest increasing α with litter quality.
During decomposition, microbes assimilate a part of the existing organic carbon to build their bodies and remaining organic carbon is oxidated into CO2 and H2O12,13,38. In the meantime, the reaction releases some energy. N is a main element of protein used for building a microbial body and a microbe also needs to absorb N while decomposing organic matter19,38. Thus, a low C/N ratio of litter helps a microbe to grow and decompose matter23,24,39. Lignin is generally thought to be the slowest decomposing component, although some observations contradicting this idea have been reported22,40,41,42. Low lignin contents and high cellulose contents both benefit microbial growth and decomposition of organic matter. Decomposition of organic matter is a very complex biochemical reaction. The carbon isotope fractionation factor α of the reaction is shown as follows:

where k and k* are the rate constants of the decomposition reaction involving 12C-substituted organic matter (or molecule) and 13C-substituted organic matter (or molecule), respectively. Since the decomposition rate of organic matter increases with decreasing C/N ratio and lignin content and increasing cellulose content, both the rate of decomposition involving 13C-substituted organic matter and the rate of decomposition involving 12C-substituted organic matter also increase. However, because 13C-substituted organic matter (or molecule) has a higher activation energy than 12C-substituted organic matter (or molecule), the decomposition reaction involving 13C-substituted organic matter increases its rate less than the reaction involving 12C-substituted organic matter. This feature causes α to increase with decreasing C/N ratio and lignin content and with increasing cellulose content of litter.
The second explanation for a change in α with litter quality is that the properties and compositions of microbial decomposer communities are associated with litter quality43,44,45. Different microbes have different metabolic pathways even when they decompose the same organic compound43,45,46 and the extent of isotope fractionation during decomposition may be tightly related to the metabolic pathways of microbes43. For example, Morasch et al. (2001) observed a greater hydrogen isotope fractionation for toluene degradation in growth experiments with the aerobic bacterium P. putida mt-2 and a less fractionation in toluene degradation by the anaerobic bacteria47.
High soil water content could lead to the formation of an anaerobic environment that limits microbial growth and decomposition of organic matter. However, this study shows that higher soil water content is related to a greater carbon isotope fractionation. The effect of water availability on isotope fractionation is also associated with compositions of microbial communities. Microbial communities in aerobic environments differ from those in anaerobic environments. As mentioned above, different microbes could use different metabolic pathways to decompose the same organic compound, thus, aerobic microbes could produce a different isotope fractionation from that produced by anaerobic microbes. For example, Griebler et al. (2004) did not observe a significant carbon isotope fractionation during mineralization of 1,2,4-trichlorobenzene by the aerobic strain Pseudomonas sp. P51, which used a dioxygenase for the initial enzymatic reaction48. In contrast, carbon isotope enrichment factors were between −3.1‰ and −3.7‰ for the degradation of 1,2,3- and 1,2,4-trichlorobenzene by the anaerobic strain Dehalococcoide sp.
For a pure chemical reaction, the magnitude of the kinetic fractionation factor α is dependent on temperature and difference of activation energy (ΔQ) between heavy and light isotopically substituted molecules of the reactant49. Parameter α decreases with an increase in temperature when ΔQ remains constant47,49. However, for isotope fractionation in a biochemical reaction, in addition to being affected by temperature and the ΔQ, it also depends on the activity of enzymes and microbes because an increase in activity of enzymes and microbes benefits enhanced decay rate, causing a greater isotopic fractionation during decomposition50,51. Rising temperature generally leads to elevated enzyme and microbe activities44,45,46, leading to increase in isotope fractionation. Coleman et al. (1981) found that undefined cultures of methane-oxidizing bacteria displayed greater carbon isotope fractionation at 30 °C than 11.5 °C52. However, current study found that temperature had no effect on α in most cases. The potential mechanism is that with decreasing temperature, the α induced by a pure chemical process would increase whereas the α induced by enzyme and microbe would decrease owing to a decrease in enzyme and microbe activities. Most probably the α induced by enzyme and microbe offset the α induced by a pure chemical process. This eventually rendered temperature to exhibit no impact on isotope fractionation.
The quantile regression showed that fractionation factor α at high quantiles was independent of soil water and MAT, suggesting that a great isotope fractionation during SOM decomposition was associated with high litter quality which positively influenced α variance during decomposition (Table 1). Organic matter with high quality often maintains high decay rate, consequently, a great α is produced. Table 1 shows that α variance at low quantiles was affected by both litter quality and environmental factors, indicating that the decomposition of organic matter with low quality depended on environmental conditions, especially soil water status. Quantile regression further demonstrates that the impact of temperature on fractionation was positive in most cases although it was not slight. The finding suggests that the α induced by enzyme and microbe was slightly bigger than the α induced by a pure chemical process.
Although the multiple regression of α against 7 variables including C/N, lignin, cellulose, W1, W2, MAT and MST is highly significant, only 36% of the α variance can be explained by these environmental and biotic factors. Three potential reasons were responsible for such a low amount of explanation. 1) In this study, we calculated fractionation factor α of each soil profile based on the approach of Poage and Feng (2004)13. However, they explained the 13C profile solely by fractionation of organic matter during decomposition and ignored the other hypotheses such as different isotopic signature of root litter compared to surface litter or temporal changes of isotope composition of vegetal inputs progressively incorporated into the soil13. 2) Soil layer is too thin, even less than 20 cm, at some sampling sites, thus, the obtained α values in these profiles with thin soil layer may be not very reliable due to limited δ13CSOM data. 3) The soil water contents used in this study were measured in the dry season, while fractionation during organic matter decomposition is dependent of long-term soil water conditions. Thus, further studies are needed in the regard.
Although there were some sorts of limitations mentioned above, we are fairly confident that the α values obtained in current study reflect the actual values of the isotope fractionation factor during organic matter decomposition in the ecosystem studied. This confidence is based on the following two facts: 1) the average δ13C values did not display differences among the bulk individual, leaf, stem and root in our other study conducted recently on 22 C3 plants and 6 C4 plants (unpublished). 2) A previous investigation conducted in the same study area showed that the mean δ13C difference between 0–5 cm soil layer and vegetation was very big, 2.87‰, while the mean δ13C differences between 0–5 cm and 5–10 cm and 10–20 cm soil layer were very small, 0.17‰ and 0.62‰, respectively36. The 2.87‰ difference between 0–5 cm layer and vegetation showed the combined contributions of carbon isotope fractionation during organic matter decomposition and the temporal changes of plants δ13C induced mainly by the δ13C decrease in atmospheric CO2 since the industrial revolution. In present study, Δδ13CSOM in Eqn. 4 is the δ13C difference between mineral soil samples and the surface mineral soil (0–5 cm depth), thus, the effect of δ13C temporal changes of plant inputs on the α calculation should be small and could be neglected. On the other hand, all α values obtained were very small with an average of 1.00118. If the temporal changes of plants δ13C contained in the α calculation, the α values obtained will be probably negative, which would be inconsistent with the actual situation.
In conclusion, this study shows that the magnitude of isotope fractionation during SOM decomposition was related to biotic and environmental factors. Litter quality and soil water content both had positive impact on α whereas temperature displayed no effect.
Methods
Study site
Mount Gongga is located in the southeastern side of the Tibetan Plateau (101°30΄ ~ 102°10΄E, 29°20΄ ~ 30°00΄N). There are remarkable differences in terrain and climate between the eastern and western slopes of this area. We selected the eastern slope of Mount Gongga as a study site because it consists of many climate types, diverse ecological systems and stable vegetation types ranging from tropical, subtropical to cold zone and relatively little human disturbance. The eastern slope belongs to an alpine gorge landform. The altitude of the eastern slope of Mount Gongga varies from 1100 m a.s.l. (Dadu River valley) to 7600 m a.s.l. and its climate is warm and dry at low elevations and cold and moist at high elevations. On the slope, temperature decreases and precipitation may increase with increasing altitude; this feature is based on the records of two meteorological observatories on the slope53.
An intact and continuous vertical vegetation spectrum can be observed along the eastern slope of Mount Gongga. It consists of subtropical evergreen broad-leaved vegetation (1100–2200 m, including a semi-arid valley with shrubs and grasses (<1500 m), evergreen broad-leaved forests and deciduous broad-leaved forests), temperate coniferous and broad-leaved mixed forests (2200–2800 m), frigid dark coniferous forests (2800–3600 m), alpine subfrigid shrub and meadow vegetation (3600–4200 m), alpine frigid meadow vegetation (4200–4600 m), alpine frigid sparse grasses and a desert zone (4600–4800 m) and a high-altitude alpine ice-and-snow zone (>4900 m). The vertical distribution of soil on the eastern slope of Mount Gongga is also very pronounced and a continuous soil sequence occurs from 1100 m to 4900 m. It consists of yellow-red soil (luvisols) (<1500 m), yellow-brown soil (luvisols) (1500–1800 m), brown soil (1800–2200 m) (luvisols), dark-brown soil (luvisols) (2200–2800 m), dark-brown forest soil (luvisols) (2800–3600 m), black mattic soil (cambisols) (3600–4200 m), mattic soil (luvisols) (4200–4600 m) and chilly desert soil (cryosols) (>4600 m)54.
Sample collection
A vertical transect spanning from 1200 m a.s.l. to 4500 m a.s.l., across five vegetation types and seven soil types, was set on the eastern slope of Mount Gongga. In August 2004, samples (including plant leaves, litter and soil) were collected along the transect at intervals of about 100 m. The method of plant sampling was described in previous papers54,55. At the most sampling sites, we set three plots (0.5 m × 0.5 m) within a 200 m2 area. All aboveground litters within a plot were collected and then, a soil profile was dug to the weathered rock. The depth of a soil profile depended on the depth of the weathered rock and most profiles had the depth of 40 cm to 50 cm. In total, 27 sites with 75 plots and 75 soil profiles were sampled along the transect. Organic layers above mineral soil were defined as “litter.” Depth zero refers to the top of the mineral horizon. Litter samples (0.25 m2) were separated into one to four layers, depending on the humus type. Layers were separated and defined by visual aspect according to Kubiena (1953)56. The first layer (L1) contains entire leaves remaining from the last fall. The second layer (L2) consists of partial leaves and partially decomposed small wood. The third layer (F) consists of small pieces (<10 mm) of decomposed leaves and small wood. The fourth layer is the dark, fine, moders and mors. Mineral soil was collected at 5 cm intervals down to a 10 cm depth, after which it was collected at 10 cm intervals down to the bottom of the soil profile.
Measurements of soil water content
Soil water content for each of three layers (0–5 cm, 5–10 cm and 10–20 cm) was determined by comparing the weight of wet and dry soils. Wet soil, the intact natural soil, was oven-dried at 105 °C until the weight did not change anymore.
Measurements of δ13C and C concentration of soil organic matter
Soil was oven-dried at 50 °C for 24 h; afterward, stones and plant residues in soil were removed; finally, soil was ground and filtered through a 2 mm sieve. About 3 g of soil consisting of particles less than 2 mm in diameter was immersed by excessive HCl (1 mol/l) for 24 h to remove carbonate and then washed to neutrality by distilled water57. Finally, the soil was oven-dried at 50 °C and ground into a fine powder. Measurements of δ13C and C concentration of SOM were determined on a DeltaPlusXP mass spectrometer (Thermo Scientific, Bremen, Germany) that was coupled with an elemental analyzer in continuous flow mode. The elemental analyzer (FlashEA 1112; CE Instruments,Wigan, UK) combustion temperature was 1020 °C.
The carbon isotopic ratios are reported in standard notation, relative to the V-PDB standard. The standard deviations for measurements of soil δ13C and soil C concentrations were less than 0.2‰ and 0.1%, respectively.
Measurements of C and N concentrations in litter
The litter samples (L1) were oven-dried at 65 °C and ground into a fine powder. The measurements of C and N concentrations were conducted on an elemental analyzer (FlashEA 1112; CE Instruments, Wigan, UK). The standard deviations for measurements of litter C and N both were less than 0.1%.
Measurements of fiber and lignin in litter
Fiber is the insoluble residue in litter after removing fat, starch, protein and sugar by acid detergent. It includes cellulose and lignin. Lignin is the insoluble residue after dissolving fiber by sulfuric acid.
Fiber was obtained through the following steps. The first step was to pour 100 ml of hot acid detergent in a beaker with 1 g of litter (previously ground and sieved through a 2 mm sieve), covered the condensing ball, opened the cooling water, quick heated the beaker to a boiling state and then maintained boiling for 60 min. The second step was to pour the solution into a filter crucible and then vacuumed and filtered the solution so that all acid was removed. The third step was to wash the residues left in the filter crucible two times with 40 ml acetone and then filter the solution until the filtrate was transparent. Each wash lasted 3–5 min. The final step was to place the filter crucible with residues into a ventilate cabinet until all of the acetone evaporated, dried the filter crucible with residues for 4 h at 105 °C and then weighed the filter crucible with residues and recorded the mass. We denoted the recorded mass as m2. The fiber content (%) was calculated by the following equation (1):

where m is the sample mass (1 g in this measurement), m1 is the crucible mass and m2 is the total mass of the crucible and residue.
Lignin was obtained through the following steps. The first step was to place the above residue into a 50 ml beaker, then 12.0 mol L−1 sulfuric acid was poured into the beaker and let the acid digest for 3 h at 20–25 °C. The second step was to pour the solution into a filter crucible, vacuum and filter the solution so that all of the acid was removed and then repeatedly washed the residual material with hot water until its pH equaled 7. The residue was lignin and the amount of cellulose was the difference between the fiber and lignin amounts.
Definitions and basic equations
The isotope fractionation factor α indicates the degree of isotope fractionation; a larger α value means a greater fractionation. In Poage and Feng (2004)13, the carbon isotope fractionation factor α during decomposition of organic carbon was defined as

where RSOM and RCO2 are the 13C/12C ratios of organic carbon (substrate) and respired CO2 (product), respectively.
In this study, we calculated the carbon isotope fractionation factor α of decomposing organic matter by using the data of δ13CSOM of soil profiles. It must be noted that only the soil profiles from sites with constant C3 or C4 vegetation can be used to do the calculation. The first step to obtaining α value of each soil profile was to calculate the δ13C difference (Δδ13CSOM) between mineral soil samples and the surface mineral soil (0–5 cm depth). The second step was to calculate ln(ρ/ρ0), where ρ0 is the C density of the surface mineral soil and ρ denotes the C density of mineral soil samples13. We used C concentration instead of C density in this calculation and generally, the alternative calculation does not cause big errors (personal communication with Feng). The third step was to plot Δδ13CSOM and ln(ρ/ρ0). Finally, α is calculated from the slope of the following linear equation13:

Figure 4 shows an example of obtaining α for a soil profile located at 2700 m a.s.l. on the eastern slope of Mount Gongga.

shows how to obtain α value of a soil profile located at 2700 m a.s.l. on the eastern slope, Mount Gongga.
The α value was estimated indirectly as the slope of the linear equation between Δδ13C and ln(C/C0).
Ecologists often use isotope discrimination, ε, to describe fractionation of a biochemical process:

where Rp is the 13C/12C ratio of the product (CO2) and RS the source (substrate) ratio. Comparing Eqns. 3 and 5, one can obtain Eqn. 6:

Eqn. 6 shows that a larger α value means a greater absolute ε value, which indicates a greater isotope fractionation.
Statistical analyses
In this study, a series of bivariate correlation analyses were first conducted to preliminarily exam the effects of different environmental and biotic factors (including C/N ratio, lignin, cellulose, W1, W2, MAT and MST) on fractionation factor α, then a stepwise regression of α vs. the 7 variables was run with forward direction (criteria: probability of F to enter r ≤ 0.05, probability of F to remove r ≥ 0.100) so that the most important influential factors could be determined. In order to effectively describe the influence of environmental and biotic variables on the minimum and maximum of range of fractionation factors, quantile regression of α vs. C/N, W1 and MAT was carried out. We used the statistical software SPSS 11.0 (SPSS Inc., Chicago, IL, USA) for the analyses of correlation and stepwise regression and the Stata/SE120 for Windows (StataCorp LP, USA) for the quantile regression.
Additional Information
How to cite this article: Wang, G. et al. Effects of environmental and biotic factors on carbon isotopic fractionation during decomposition of soil organic matter. Sci. Rep. 5, 11043; doi: 10.1038/srep11043 (2015).
References
Natelhoffer, K. J. & Fry, B. Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter. Soil Sci. Soc. Am. J. 52, 1633–1640. 10.2136/sssaj1988.03615995005200060024x (1988).
Balesdent, J., Cirardin, C. & Mariotti, A. Site-related δ13C of tree leaves and soil organic matter in a temperate forest. Ecology 74, 1713–1721 (1993).
Ågren, G. I. & Bosatta, E. Quality: A bridge between theory and experiment in soil organic matter studies. Oikos 76, 522–528 (1996).
Ågren, G. I., Bosatta, E. & Balesdent. J. Isotope discrimination during decomposition of organic matter: A theoretical analysis. Soil Sci. Soc. Am. J. 60, 1121–1126 (1996).
Fernandez, I., Mahieu, N. & Cadisch, G. Carbon isotopic fractionation during decomposition of plant materials of different quality. Glo. Biogeochem. Cyc. 17, 1075, 10. 1029/2001GB001834 (2003).
Wang, G. A., Feng, X., Han, J. & Zhou, L. P. Paleovegetation reconstruction using δ13C of soil organic matter. Biogeosciences 5, 1325–1337(2008).
Risk, D., Nickerson, N., Phillips, C. L., Kellman, L. & Moroni, M. Drought alters respired δ13CO2 from autotrophic, but not heterotrophic soil respiration. Soil Biol. Biochem. 50, 26–32. 10.1016/j.soilbio.2012.01.025 (2012).
Ciais, P., Tans, P. P., Trolier, M., White, J. W. C. & Francey, R. J. A large northern hemisphere terrestrial CO2 sinks indicated by the 13C/12C ratio of atmospheric CO2 . Science 269, 1098–1102. 10.1126/science.269.5227.1098 (1995).
Battle, M., Bender, M. L., Tans, P. P., White, J. W. C. & Eily, J. T. Global carbon sinks and their variability inferred from atmospheric O2 and δ13C. Science 287, 2467–2470 (2000).
Rayner, P. J., Law, R. M., Allison, C. E., Francey, R. J., Trudinger, C. M. & Pickett-Heaps, C. Interannual variability of the global carbon cycle (1992–2005) inferred by inversion of atmospheric CO2 and δ13CO2 measurements. Glob Biogeochem. Cyc. 22, GB3008, 10.1029/2007GB003068 (2008).
Alden, C. B., Miller, J. B. & White, J. W. C. Can bottom-up ocean CO2 fluxes be reconciled with atmospheric 13C observations? Tellus 62B, 369–388 (2010).
Feng, X. A theoretical analysis of carbon isotope evolution of decomposing plant litters and soil organic matter. Glob. Biogeochem. Cyc. 16, 1119, 2002, 10.1029/2002GB001867 (2002).
Poage, M. A. & Feng, X. A theoretical analysis of steady state δ13C profiles of soil organic matter. Glob. Biogeochem. Cyc, Vol.18, GB2016, 10.1029/2003GB002195 (2004).
Cerling, T. E., Quade, J., Wang, Y. & Bowman, J. R. Carbon isotopes in soils and palaeosols as ecology and palaeoecology indicators. Nature 341, 138–139 (1989).
Hatté, C. et al. δ13C of loess organic matter as a potential proxy for paleoprecipitation. Quat. Res. 55, 33–38 (2001).
Liu, L., Zhou, X., Yu, Y. Y. & Guo, Z. T. The Natural vegetation on the Chinese Loess Plateau: the evidence of soil organic carbon isotope. Quat. Sci. 31, 506–513 (2011).
Rao, Z. G. et al. High-resolution summer precipitation variations in the western Chinese Loess Plateau during the last glacial. Scientific Reports 3, 2785. 10.1038/srep02785 (2013).
Wang, G. A., Li, J. Z., Liu, X. Z. & Li, X. Y. Variations in carbon isotope ratios of plants across a temperature gradient along the 400 mm isoline of annual precipitation in north China and relevance to paleovegetation reconstruction. Quat. Sci. Rev. 63, 83–90 (2013).
Magill, A. H. & Aber, J. D. Long-term effects of experimental nitrogen additions on foliar litter decay and humus formation in forest ecosystems. Plant Soil 203, 301–311 (1998).
Trumbore, S. E., Chadwick, O. & Amundson, A. Rapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change. Science 272, 393–396 (1996).
Buchmann, N. Biotic and abiotic factors controlling soil respiration rate in Picea abies stands. Soil Bio. Biochem. 32, 1625–1635 (2000).
Schmidt, M. W. I. et al. Persistence of soil organic matter as an ecosystem property. Nature 478, 49–56. 10.1038/nature10386 (2011).
Sinsabaugh, R. L., Carreiro, M. M. & Repert, D. A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition and mass loss. Biogeochem. 60, 1–24 (2002).
Saiya-Cork, K. R., Sinsabaugh, R. L. & ZaK, D. R. The effects of long-term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Bio. Biochem. 34, 1309–1315 (2002).
Garten, C. T. Jr, Cooper, L. W., Post, W. M. III. & Hanson, P. J. Climate controls on forest soil C isotope ratios in the southern Appalachian Mountains. Ecology 81, 1108–1119 (2000).
Melillo, J. M. et al. Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. Plant Soil 115, 189–198 (1989).
Giardina, C. P. & Ryan, M. G. Evidence that decomposition rates of organic carbon inmineral soil do not vary with temperature. Nature 404, 858–861 (2000).
Trumbore, S. E. & Czimczik, C. I. An uncertain future for soil carbon. Science 321, 455–1456. 10.1126/science.1160232 (2008).
Kirschbaum, M. U. F. The temperature dependence of organic-matter decomposition—still a topic of debate. Soil Biol. Biochem. 38, 2510–2518. 10.1016/j.soilbio.2006.01.030 (2006).
Von Lützow, M. & Kögel-Knabner, I. Temperature sensitivity of soil organic matter decomposition—what do we know? Bio. Fert. Soils 46, 1–15. 10.1007/s00374-009-0413-8 (2009).
Knicker, H. Soil organic N - An under-rated player for C sequestration in soils? Soil Bio. Biochem. 43, 1118–1422. 10.1016/j.soilbio.2011.02.020 (2011).
Cahoon, S. M. P., Sullivan, P. F., Shaver, G. R., Welker, J. M. & Post, E. Interactions among shrub cover and the soil microclimate may determine future Arctic carbon budgets. Ecol. Lett. 15, 1415–1422. 10.1111/j.1461-0248.2012.01865.x (2012).
Freschet, G. T. et al. Linking litter decomposition of above- and below-ground organs to plant–soil feedbacks worldwide. J. Ecol. 101, 943–952 (2013).
Wang, G., Zhou Y., Xu, X., Ruan, H. & Wang, J. Temperature Sensitivity of Soil Organic Carbon Mineralization along an Elevation Gradient in the Wuyi Mountains, China. PLoS ONE 8: e53914. 10.1371/journal.pone.0053914 (2013).
Feng, X. et al. Distribution, accumulation and fluxes of soil carbon in four monoculture lysimeters at Sam Dimas Experimental forest, California. Geochim Cosmochim. Acta 63, 1319–1333 (1999).
Chen, P. N., Wang, G. A., Han, J. M., Liu, M. & Liu. X. Q. δ13C difference between plants and soils on the eastern slope of Mount Gongga. Chin. Sci. Bull. 55, 55–62, 10.1007/s11434-009-0405-y (2010).
Li, J. Z. et al. Variations in carbon isotope ratios of C3 plants and distribution of C4 plants along an altitudinal transect on the eastern slope of Mount Gongga. Sci. China Ser. D-Earth Sci. 52, 1714–1723, 10.1007/s11430-009-0170-4 (2009).
Luo, Y. Q. & Zhou, X. H. Soil respiration and the Environment. (Elsevier, Inc. 2006).
Berg, B., Wessen, B. & Ekbohm, G. Nitrogen level and lignin decomposition in Scots pine needle litter. Oikos 38, 291–296 (1982).
Amelung, W., Brodowski, S., Sandhage-Hofmann, A. & Bol. R. Combining biomarker with stable isotope analysis for assessing the transformation and turnover of soil organic matter. Adv. in Agronomy 100, 155–250 (2008).
Grandy, A. S. & Neff, J. C. Molecular C dynamics downstream: the biochemical decomposition sequence and its impact on soil organic matter structure and function. Sci. Total Environ. 404, 297–307. 10.1016/j.scitotenv.2007.11.013 (2008).
Marschner, B. et al. How relevant is recalcitrance for the stabilization of organic matter in soils? J. Plant Nut. Soil Sci. 171, 91–110. 10.1002/jpln.200700049 (2008).
Macko, S. A. & Estep, M. L. F. Microbial alteration of stable nitrogen and carbon isotopic composition of organic matter. Org. Geochem. 6, 787–790 (1984).
Atlas, R. M. & Bartha, R. Microbial Ecology: Fundamentals and Applications. (Benjamin/Cummings Science Publishing, 1998).
He, W. X. & Hong, J. P. Environmental Microbiology. (China Agricultural University Press, 2006).
Xu, X. H. General Microbiology. (China Agricultural University Press, 1992).
Morasch, B., Richnow, H. H., Schink, B. & Meckenstock, R. U. Stable hydrogen and carbon isotope fractionation during microbial toluence degradation: mechanistic and environmental aspects. App. Environ. Microbiol. 67, 4842–4849 (2001).
Griebler, C., Adrian, L., Meckenstock, R. U. & Richnow, H. H. Stable carbon isotope fractionation during aerobic and anaerobic transformation of trichlorobenzene. FEMS Microbiol. Ecol. 48, 313–321 (2004).
Guo, Z. Y. Stable Isotopic Chemistry. (Science Press, 1984).
Körner, C., Farquhar, G. D. & Roksandic, Z. A global survey of carbon isotope discrimination in plants from high altitude. Oecologia 74, 623e632 (1988).
Körner, C., Farquhar, G. D. & Wong, S. C. Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 88, 30e40 (1991).
Coleman, D. D., Risatti, B. & Schoell, M. Fractionation of carbon and hydrogen isotopes by methane-oxidizing bacteria. Geochim. Cosmochim. Acta 45, 1033–1037 (1981).
Zhong, X. H. et al. Researches of the Forest Ecosystems on Mount Gongga. (Chengdu University of Science and Technology Press, 1997).
Liu, X. Z. & Wang, G. A. Measurements of nitrogen isotope composition of plants and surface soils along the altitudinal transect of the eastern slope of Mount Gongga in southwest China. Rap. Commun. Mass Spectrom. 24, 3063–3071 (2010).
Shi, W. Q., Wang, G. A. & Han, W. X. Altitudinal variation in leaf nitrogen concentration on the eastern slope of Mount Gongga on the Tibetan Plateau, China. PLoS ONE 7: e44628. 10.1371/journal.pone.0044628 (2012).
Kubiena, W. L. The soils of Europe. Illustrated diagnosis and systematics. London: Thomas Murphy (1953).
Brodie, C. R. et al. Evidence for bias in C and N concentrations and δ13C composition of terrestrial and aquatic organic materials due to pre-analysis acid preparation methods. Chem. Geol. 282, 67–83 (2011).
Acknowledgements
This research was supported by grant from the National Natural Science Foundation of China (No.41272193) and the Chinese National Basic Research Program (No. 2014CB954202). We would like to thank Ma Yan for analyzing carbon isotopes at the Stable Isotope Laboratory of the College of Resources and Environment, China Agricultural University.
Author information
Authors and Affiliations
Contributions
W. G. A conceived the study. W.G.A. and J.Y.F carried out the experiments and performed the data analyses. W. G. A. wrote the manuscript. J. Y. F and L.W. reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, G., Jia, Y. & Li, W. Effects of environmental and biotic factors on carbon isotopic fractionation during decomposition of soil organic matter. Sci Rep 5, 11043 (2015). https://doi.org/10.1038/srep11043
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep11043
This article is cited by
-
The Fate of Soil Organic Carbon from Compost: A Pot Test Study Using Labile Carbon and 13c Natural Abundance
Journal of Soil Science and Plant Nutrition (2024)
-
Palaeobotanical evidence reveals the living conditions of Miocene Lufengpithecus in East Asia
BMC Plant Biology (2023)
-
Preferential substrate use decreases priming effects in contrasting treeline soils
Biogeochemistry (2023)
-
A study on benthic molluscs and stable isotopes from Kutch, western India reveals early Eocene hyperthermals and pronounced transgression during ETM2 and H2 events
Swiss Journal of Palaeontology (2022)
-
The linkage of 13C and 15N soil depth gradients with C:N and O:C stoichiometry reveals tree species effects on organic matter turnover in soil
Biogeochemistry (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
