
Abstract
The dynamics of association between pathogens and vectors can strongly influence epidemiology. It has been proposed that wilt disease epidemics in cucurbit populations are sustained by persistent colonization of beetle vectors (Acalymma vittatum) by the bacterial phytopathogen Erwinia tracheiphila. We developed a qPCR method to quantify E. tracheiphila in whole beetles and frass and used it to assess pathogen acquisition and retention following variable exposure to infected plants. We found that (i) E. tracheiphila is present in frass in as little as three hours after feeding on infected plants and can be transmitted with no incubation period by vectors given brief exposure to infected plants, but also by persistently colonized vectors several weeks following exposure; (ii) duration of exposure influences rates of long-term colonization; (iii) frass infectivity (assessed via inoculation experiments) reflects bacterial levels in frass samples across time; and (iv) vectors rarely clear E. tracheiphila infections, but suffer no apparent loss of fitness. These results describe a pattern conducive to the effective maintenance of E. tracheiphila within cucurbit populations.
Similar content being viewed by others


Introduction
Many plant pathogens rely on arthropod vectors to carry them from one host plant to the next. While some of these pathogens form only transitory associations with their vectors1, others form more durable relationships in which an individual infectious vector is capable of transmitting the pathogen to multiple hosts over extended periods of time2,3,4. The ability of pathogens to form such long-lasting associations with vectors—in some cases colonizing the vector as a secondary host—may facilitate dispersal across heterogeneous landscapes and pathogen persistence in vector reservoirs independent from the lifecycle of the host plant. Previous studies, most from plant virus systems, show that diverse aspects of the pathogen-vector interaction (e.g., pathogen adherence to internal structures, the duration of latency periods and pathogen propagation within the vector) can significantly influence pathogen transmission, with implications for disease ecology and evolution5,6,7. Transmission can furthermore be influenced by pathogen effects on vector health and behavior, which may be either direct or mediated by pathogen-induced changes in the host plant6,8. To date, the detailed interactions of bacterial plant pathogens with their arthropod vectors have been explored less extensively than those of plant viruses, even though a number of ecologically and economically important plant diseases are caused by obligately vectored bacteria3,9,10.
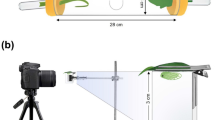
Erwinia tracheiphila Smith (Enterobacteriaceae), the causative agent of a bacterial wilt disease that afflicts wild and cultivated cucurbit plants, is obligately transmitted by spotted and striped cucumber beetles (Coleoptera: Chrysomelidae: Luperini)11,12. Beetles acquire the pathogen by feeding on infected plant tissue and transmit it by depositing infective frass at sites of recent (foliar) feeding damage (Figure 1) or onto floral nectaries13. The necessity for the pathogen to access host-plant vasculature via these specific routes in order to successfully infect, coupled with the haphazard deposition of frass by feeding beetles (Figure 1), suggests that transmission via frass is relatively inefficient. Once bacterial cells successfully enter the host plant, they replicate in the xylem (Cucurbita spp. and Cucumis spp.) and secrete an extracellular polysaccharide matrix that prevents the passage of xylem sap, causing vessel occlusion and characteristic wilting symptoms14. Bacterial wilt disease is typically fatal once symptoms appear15 and causes significant economic losses in cultivated cucurbit crops12. It has previously been suggested that E. tracheiphila colonization of cucumber beetles, as secondary hosts, may play an important role in maintaining epidemics within host populations and in the initiation of new epidemics across growing seasons16. However, relatively little is known about the quantitative and temporal dynamics of interactions between E.tracheiphila and its vectors or their implications for host-plant exposure to the pathogen.

Frass deposition on leaves of C. pepo ssp. texana after cucumber beetle feeding.
Previous work has produced conflicting results regarding the dynamics of long-term colonization of cucumber beetles by E. tracheiphila17,18 and how long beetles continue to produce infective frass after they have been exposed to the pathogen18. An initial attempt to quantify E. tracheiphila within striped cucumber beetles (Acalymma vittatum) via fuschin staining reported so few bacteria in the beetle intestinal tract that the authors hypothesized there must be an alternative environmental reservoir for the pathogen19, but this was subsequently shown not to be the case16. Later work using bacteria-smeared cotyledon disks as inoculation sources for beetles and DAS-ELISA and immunoperoxidase localization as a detection method revealed stronger fluorescence in the hindgut at 10 and 30 days post acquisition compared to 3 days post acquisition, suggesting bacterial replication within the vector17, but another study failed to detect E. tracheiphila in beetle frass with standard PCR at 6 days post acquisition18.
To achieve a better understanding of the dynamics of E. tracheiphila association with its insect vector, we developed a qPCR assay to measure relative amounts of E. tracheiphila in whole beetles and beetle frass at multiple time points following exposure to infected host plants (0, 5 and 28 days) and also assessed rates of Erwinia tracheiphila acquisition and retention over time as a function of the exposure period (the length of time that beetles spent feeding on infected plants). In addition, we performed plant inoculation experiments with beetle frass to determine how the likelihood of pathogen transmission to new hosts (through leaf wounds) was influenced by time since exposure. Finally, we examined the effects of E. tracheiphila on beetle fitness by measuring oviposition and survival in exposed and non-exposed beetles. The results from these assays, taken together with other recent findings, provide important insights into the disease dynamics of Erwinia tracheiphila within cucurbit populations.
Results
Acquisition and retention: frass
Seven of 131 frass samples failed to amplify either 18S or EtOMP and were omitted from the analyses. Two frass samples that amplified EtOMP but not 18S were included in counts for acquisition and retention rates, but were not used for quantitative analyses (because while they could be positively identified as E. tracheiphila exposed, the lack of 18S prevented its use for total DNA normalization). Furthermore, three beetles did not produce frass at the 5-day time point and three did not produce frass at the 28-day time point and these were also omitted. To assess the acquisition and retention rates (number of E. tracheiphila positive beetles out of all beetles in the trial), a chi-square test revealed that the probability of frass containing E. tracheiphila immediately after beetles were removed from infected plants was not significantly influenced by exposure period (3 hr vs. 24 hr) (χ2 = 1.583, df = 1, P = 0.208) (Figure 2a, b). However, at 5 days post exposure significantly more beetles in the 24 hr exposure group were E. tracheiphila-positive compared to the 3 hr exposure group (χ2 = 4.014, df = 1, P = 0.045) and this difference persisted at 28 days post exposure (χ2 = 5.743, df = 1, P = 0.017) (Figure 2a, b). These data thus indicate that the length of the exposure period influences the likelihood of the pathogen forming a long-term association with the beetle.

Percentages of frass samples and whole beetles that were positive for Erwinia tracheiphila immediately (0 days), 5 days and 28 days post exposure.
(2a) Frass samples following 3 h exposure; (2b) Frass samples following 24 h exposure; (2c) Whole-beetle samples following 24 h exposure.
Quantitative dynamics: frass
Because the amount of bacteria shed in frass is an important factor influencing the level of exposure of healthy plants to the wilt pathogen, we also sought to understand the quantitative and temporal dynamics of bacterial levels in beetle frass. E. tracheiphila form localized blockages in plant xylem vessels and are thought to be heterogeneously distributed throughout the vasculature. To assess whether this might affect the amount of bacteria that beetle vectors are exposed to through feeding on symptomatic plants, Bartlett's test for equal variances was applied to untransformed EtOMP/18S ratios to compare the ranges of bacterial levels present in frass 0, 5 and 28 days post acquisition. The variance of relative bacterial levels was significantly larger at 0 and 28 days compared to 5 days (Bartlett's K2 = 100.97, df = 2, P < 0.005) (Figure 3a).

Distribution (variance) of relative E. tracheiphila/totalDNA ratios in frass (3a) and whole beetles (3b) immediately (0 days), 5 days and 28 days post exposure.
To determine the effects of exposure period (3 hr vs. 24 hr) and time since exposure (0, 5, 28 days) on the amount of E. tracheiphila relative to total plant DNA in the frass, the EtOMP/18S ratios were first log + 1 transformed for normality. Because frass was collected from the same beetles at three time points, a repeated measures model was used in SAS PROC MIXED20 on the log + 1 transformed EtOMP/18S ratios. The results indicated that Exposure (3 hr or 24 hr) did not significantly affect EtOMP/18S ratios (F1,32 = 0.2, P = 0.876); therefore, the 3 hr and 24 hr exposure groups were pooled to test the effect of time post exposure (0, 5, or 28 days). We found that while the ratio of E. tracheiphila shed in frass was not significantly different between acquisition groups (3 hr or 24 hr), time post exposure was a significant factor (F2,21 = 5.86, P = 0.0095). Individual comparisons revealed significantly more E. tracheiphila in frass both immediately following exposure (0 days) and at 28 days after exposure compared to 5 days after exposure. The difference between EtOMP/18S ratios immediately (0 days) and 28 days after an acquisition period was not significant (Figure 4). The repeated measures model statement was SAS PROX MIXED log (1 + EtOMP/18S) = Exposure + Time post exposure + (Exposure*Time post exposure); Repeated Time/Subject = Beetle (SAS 2011).

Relative ratios of EtOMP/18S for whole beetles (left) and beetle frass (right) immediately (0 days), 5 days and 28 days post exposure.
Acquisition and retention: whole beetles
One of the 74 beetles in whole-insect assays was not scored due to failed DNA extraction determined by lack of product for both 18S and EtOMP. The remaining 73 beetles were unambiguously scored as positive or negative for E. tracheiphila after feeding for 24 hrs on E. tracheiphila infected plants. A moderately higher percentage of beetles were positive for E. tracheiphila immediately (84%) and at 5 days (83%) following exposure than at 28 days (64%) following exposure, but the proportion of E. tracheiphila-positive beetles was not significantly different between any time points (0 days vs. 5 days χ2 = 0, df = 1, P = 1; 0 days vs. 28 days χ2 = 1.906, df = 1, P = 0.168; 5 days vs. 28days χ2 = 1.689, df = 1, P = 0.194) (Figure 2c). Furthermore, the frequency of E. tracheiphila positive beetles from the whole beetle experiment was statistically indistinguishable from the proportion of E. tracheiphila positive beetles in the 24 hr acquisition group in the frass experiment (0 days frass vs. 0 days beetle χ2 = 3.149, df = 1, P = 0.076; 5 days frass vs. 5 days beetle χ2 = 2.573, df = 1, P = 0.1089; 28 days frass vs. 28 days beetle χ2 = 0, df = 1, P = 1). In both the whole-beetle and frass assays, more than 60% of beetles exposed for 24 h retained the pathogen 28 days after exposure (Figure 2).
Quantitative dynamics: whole beetles
Bartlett's test of variances of EtOMP/18S ratios was used to examine changes in the variance of ratios at different time points post exposure. For untransformed data, Bartlett's test of EtOMP/18S ratios rejects the null hypothesis of equal variances (Bartlett's K2 = 175.93; df = 2, P < 0.005). Immediately after acquisition, there was a wide variance in relative levels of E. tracheiphila present (Figure 3b). By 5 days and continuing at 28 days post acquisition, the median amount of bacteria present remained unchanged compared to the mean at 0 days, but the variance was significantly narrower at both the 5 day and 28 day time points compared to the variance at 0 days.
To compare changes in the mean EtOMP/18S ratios at different time points, the ratios were log + 1 transformed for normality and a 1-way ANOVA showed no significant effect of time post-exposure on EtOMP/18S ratios (F2,50 = 0.80; P = 0.454) (Figure 4). Model statement was SAS PROC MIXED (log + 1 EtOMP/18S) = Time Post Exposure. We note that the four observations with the largest EtOMP/18S ratios occurred in beetles analyzed immediately following exposure (Figure 3b).
Transmission assays
We performed a series of transmission experiments to examine how changes in the amount of bacteria in frass over time following exposure affects transmission potential and inoculation success. Chi-square tests for difference in two proportions in R21 were used to analyze differences in disease development on C. pepo ssp. texana seedlings inoculated with a homogenate of frass collected from groups of 100 A. vittatum at 0, 5 and 28 days post exposure. In total, 44 out of 84 plants became infected when inoculated with frass collected immediately (0 days) after feeding, 3 out of 76 plants became infected when inoculated with frass collected five days after feeding and 22 out of 54 plants became infected when inoculated with frass collected 28 days after feeding on wilting plants. We observed significantly higher inoculation success with frass collected 0 days and 28 days post acquisition compared to frass collected 5 days post acquisition (0:5 days χ2 = 42.812, df = 1, P < 0.005; 5days:28 days χ2 = 25.967, df = 1, P < 0.005), matching the pattern observed for levels of bacteria shed in frass over time. The final proportion of infected plants after inoculation with frass collected 0 days and 28 days post acquisition was not significantly different (χ2 = 1.348, df = 1, P = 0.245) (Figure 5).

Occurrence and progression of bacterial wilt symptoms in C. pepo ssp. texana inoculated with frass collected from beetles immediately (0 days), 5 days and 28 days post exposure.
Effects of E. tracheiphila exposure on Acalymma vittatum fitness
Prolonged acquisition access periods on wilting E. tracheiphila infected plants did not significantly affect striped cucumber beetle survival, oviposition rates, or rates of adult emergence. Ten of 13 females that fed on the mock-inoculated plants and 10 of 12 females that fed on the E. tracheiphila infected plants survived for the 32-day duration of the experiment. One female from the mock-inoculated control group died 9 days post feeding and two died 20 days post feeding. In the E. tracheiphila exposure group, one beetle died 2 days after exposure and another died 23 days after exposure. Females that oviposited fewer than 20 total eggs (two from the mock group and one from the E. tracheiphila group) were excluded from the statistical analysis of oviposition rate but not survival. The total number of eggs oviposited by females in each group was not significantly different (1-way ANOVA; F1,22 = 0.32; P = 0.726). In the egg development experiment, there was no difference in the percentage of adult emergence from eggs produced by females fed on either healthy or wilting seedlings (1-way ANOVA, F1,22 = 0.50, P = 0.485).
Discussion
For bacterial phytopathogens that are obligately insect transmitted, direct interactions with vectors can determine dispersal efficiency and rates of pathogen exposure experienced by healthy plants, but these interactions are often poorly understood. The data presented here demonstrate that E. tracheiphila can persistently colonize its striped cucumber beetle vector; that exposure period to infected plants affects the probability of beetle colonization by the bacteria; and that temporal colonization dynamics affect the probability of transmission through leaf wound exposure to infective frass. Our results further demonstrate that E. tracheiphila can be transmitted via frass immediately (i.e., with no incubation period) from beetles exposed to infected plants for 24 h (Figure 5), presumably by bacterial cells carried in plant material passing through the digestive tract of the beetle, but also that E. tracheiphila can be effectively transmitted from persistently colonized beetles several weeks after exposure (Figure 4, 5).
The large variance of bacterial levels observed in whole beetles and in frass immediately following exposure to infected plants (Figure 3) likely reflects similar variation in levels of bacteria ingested during herbivory, perhaps owing in part to the heterogeneous distribution of E. tracheiphila in host plant tissues. In whole beetles, the mean EtOMP/18S ratio remains stable while the variance narrows significantly by 5 days (and continuing through 28 days), suggesting that bacterial population levels in the gut lumen converge toward an equilibrium population range in beetles that have been successfully colonized and that E. tracheiphila population levels do not continue increasing in population size as a function of time post exposure. In frass, there was a significantly wider range of relative bacterial levels at 0 and 28 days compared to 5 days post exposure, suggesting that the amount of E. tracheiphila shed in frass at 5 days post acquisition is uniformly low, but that by 28 days there is significantly more bacteria and a significantly wider range of bacterial levels, being shed in frass (Figure 3a). The contrast between quantitative changes in EtOMP/18S ratios in whole beetles and in frass, combined with the absence of latency, is consistent with the hypothesis that E. tracheiphila colonizes the surface of the digestive tract but does not cross into the hemolymph and suggests that bacterial shedding in frass is the predominant source of E. tracheiphila exposure for healthy plants (i.e., rather than regurgitation or contaminated mouthparts).
Furthermore, the length of vector exposure to diseased plants significantly affects the probability of persistent E. tracheiphila infection (Figure 2). Overall, it appears that the initial colonization process is inefficient—as indicated by the relatively low rates of persistent infection observed in beetles given short-term exposure to infected plants (Figure 2a) despite abundant levels of the pathogen passing through the digestive tracts of these beetles (Figure 3a)—but that once colonization occurs E. tracheiphila population growth within the insect's digestive tract follows a similar trajectory regardless of the length of the exposure period.
Our inoculation experiments with frass homogenates yielded maximum infection rates of ~50%. This is higher than the rates previously reported for experiments in which single beetles were caged on plants22,23,24, likely reflecting the relatively haphazard deposition of frass by feeding beetles (Figure 1) compared to the direct inoculation of homogenate into leaf wounds employed here. Differences in host-plant age may also be a contributing factor, as Cucurbita ssp. individuals become more resistant to wilt infection via leaf damage as they mature25. We recently demonstrated that transmission can also occur via floral nectaries13—cucumber beetles frequently aggregate in cucurbit flowers and deposit frass onto or near the nectaries—and it is possible that both levels of exposure and infection efficiency differ for foliar and floral transmission. In a large-scale field experiment, more than 95% of the flowers on healthy wild gourd plants were found to have E. tracheiphila on or near the nectaries, but fewer than 20% of plants in the field developed symptoms of wilt disease during the same month13. Thus, infection appears to be relatively inefficient through either foliar or floral infection routes and we hypothesize that repeated contact with frass from multiple infective vectors is likely to be necessary to achieve a high probability of transmission.
Transmission is further constrained by the high virulence of E. tracheiphila in plant hosts, which typically die within 7–14 d following the onset of wilt symptoms, providing only a limited window for acquisition of the pathogen by the vector. However, we have previously found that A. vittatum cucumber beetles are preferentially attracted to the odors of wilting plants and also exhibit a feeding preference for E. tracheiphila-infected versus healthy foliage26. Consequently, beetles are probably more likely to acquire the wilt pathogen than would otherwise be predicted based on the frequency of wilt diseased plants in the population at a given time. The current results demonstrate that beetles that do acquire the pathogen can transmit it throughout the 32 day experimental period and likely throughout the remainder of the current growing season (Figures 2–5), long after the host plant from which the pathogen was initially acquired will (almost invariably) have died from bacterial wilt disease.
Our investigation of the direct impacts of E. tracheiphila on A. vittatum survival and reproduction revealed no evidence of adverse fitness consequences for beetles exposed to infected plants. This result is compatible with our previous observations regarding beetle attraction to and feeding preferences for infected compared to healthy host plants26, as we would expect beetles to discriminate against salient cues from infected plants if feeding on those plants entailed significant fitness costs. It is furthermore compatible with the observation that obligately vector-transmitted pathogens do not typically impact their vectors in ways that might compromise transmission to the next primary host, although there are some exceptions to this general trend27,28,29,30. However, while our findings suggest that E. tracheiphila infection does not significantly impact beetle survival and reproduction, further work is needed to understand the detailed effects of the pathogen on the insect, particularly under field conditions where pathogen-induced changes in behavior or physiological efficiency might have greater consequence than is observed in our laboratory assays6. For example, the microbial communities inhabiting the guts of insect herbivores are often poorly described, though their composition can have important implications for pathogen virulence31,32, or acquisition33 and the A. vittatum gut microbiome might well be influenced by E. tracheiphila colonization.
In conclusion, these findings describe novel aspects of the dynamic relationship between E. tracheiphila and A. vittatum, the most important vector of E. tracheiphila in the northeastern and midwestern United States. They reveal that the beetles given brief exposure to infected plants can transmit the pathogen even when persistent vector colonization does not occur, in as little as one day and probably as soon as the infected plant material moves through the beetles' gut and also demonstrate that the bacteria is capable of establishing persistent colonizations that last throughout the month-long study period employed here and presumably throughout most or all of the life-span of the adult vector. Together with our previous work describing patterns of vector attraction to healthy and infected plants26, these results describe a pattern consistent with the effective spread of E. tracheiphila within cucurbit populations despite the relative inefficiency of transmission to new host plants and the high virulence of E. tracheiphila in the primary plant host, which results in rapid plant death and a limited window of time for transmission from infected plants to vectors. These findings provide new insight into the interactions of bacterial plant pathogens with their insect vectors and may have relevance for pathogen management in natural and agricultural ecosystems. Improved understanding of the ecology of pathogen-vector-host interactions can inform efforts to predict and manage the outcomes of such interactions. The current work—including development of a quantitative PCR assay for detecting E. tracheiphila—also lays a foundation for more detailed studies aimed at elucidating the detailed mechanisms of bacterial interaction with its primary and secondary hosts, which could facilitate the development of management techniques aimed at disrupting colonization or transmission34,35.
Methods
The study system
Striped cucumber beetles are specialists on cucurbits in both larval and adult stages. The larvae are rootworms, while adults feed on the foliage, flowers and fruits of mature plants36. Cucumber beetles feed compulsively on cucurbitacins, inducible triterpenoid compounds characteristic of Cucurbitaceae that are toxic to most herbivores37. Beetles are preferentially attracted to the odors of wilting leaves on E. tracheiphila infected plants and aggregate on wilting foliage in the field26, likely in part because reduced turgor pressure facilitates feeding38. Current agricultural management strategies for bacterial wilt rely on pesticide application to suppress beetle populations12 or expensive and labor-intensive use of row covers to prevent beetle contact with plants39.
Beetle rearing
An E.tracheiphila-free striped cucumber beetle colony was maintained following Mitchell 200918: Field captured adult beetles were kept in 1 ft × 1 ft Bioquip mesh cages (cat #1466A). Pro-Mix potting soil (Premier Horticulture Reviere Du Loup, Quebec, Canada) was provided as an oviposition substrate. Beetles were fed 3× per week with leaves and flowers of C. pepo ‘Raven’. Potting soil with eggs was transferred weekly to a 6 L plastic rearing container. Root-feeding larvae were continuously supplied with sprouted C. pepo ‘Raven’ seedlings until pupation. Newly eclosed adults were removed from the rearing container and placed with adults from the same weekly age cohort in 1 ft × 1 ft × 1 ft Bioquip mesh cages.
Quantitative real-time PCR
To achieve a quantitative understanding of changes in bacterial levels during long-term colonization of A. vittatum by E. tracheiphila (acquisition and retention assays described below), we used the single-copy gene EtOMP to assess the amount of E. tracheiphila present and 18S as a normalizing gene for the amount of total DNA present. This approach assumes that bacterial DNA contributes negligibly to total DNA present in our samples and that dead E. tracheiphila contribute negligible amounts of EtOMP to total bacterial DNA. 5 μl of DNA was used as template in triplicate 20 μl RealTimePCR reactions with primers and probes at 10 mM and ABI TaqMan® Gene Expression MasterMix (cat #4369016). Primers and probe were designed to anneal to conserved regions of a single-copy gene encoding the outer membrane protein A (EtOMP) of E. tracheiphila, such that all published strains of E. tracheiphila are indiscriminately detected (Primers: Et73: GGCGATCACGACACAGTTGT, Et140: CAGTTTTTGGTCAGGGCATACTCP Probe: TCCCCTCTGGCAGCCATAGGTGC). 18S was used as a normalizing gene for the total amount of DNA in each reaction (18S F: CGGCTACCACATCCAAGGAA; 18SR: TGCTGGCACCAGACTTGCCCTC; 18S Probe: CGCAAATTACCCACTCCCGGCAC)40. The cycling program comprised an initial denaturation step at 95°C for 3 minutes, 40 cycles of (i) denaturation/activation at 95°C for 10 seconds and (ii) annealing/extension at 55°C for 30 seconds; this program was carried out on an ABI Prism 7900 HT sequence detector (Applied Biosystems, Foster City, CA). Fluorescence reads were taken at the end of each annealing/extension step. A measure of the amount of E. tracheiphila in each sample was computed as the copy numbers of EtOMP relative to the copy numbers of 18S in each DNA sample. The respective copy numbers were determined via standard curves for EtOMP and 18S, using 5 serial 10-fold dilutions of DNA extracted from pure bacterial culture for the EtOMP curve and DNA extracted from clean (unexposed) colony beetles for 18S. The identity of the amplified fragments was verified by sequencing the respective PCR products.
Acquisition and retention assays
Beetle frass assays
Two Cucurbita pepo ssp. texana plants were infected with a local field isolate of E. tracheiphila using the colony-stab procedure18. Both plants were placed together in a 1 ft × 1 ft × 1 ft Bioquip mesh cage (cat #1466A) when most leaves showed wilt disease development. Seventy-five 5-day-old A. vittatum beetles from an E. tracheiphila-free beetle colony that had been starved for 48 hours were placed in the same cage with the infected plants. Groups of 23 beetles were removed after 3 hr and 24 hr exposure periods and placed individually into 0.7 ml Eppendorf tubes for frass collection. Once frass accumulation was visible inside the tube (~2 hours), each beetle was transferred to its own petri dish with water-soaked cotton and one healthy (E. tracheiphila-free) cucumber leaf as food (and subsequently transferred 4× weekly to new petri dishes). Frass was again collected from each beetle (via the same method) at 5 and 28 days post exposure. Following each frass collection, DNA was immediately extracted using prepGem Bacteria extraction kits (ZyGEM Corp Ltd. Hamilton, New Zealand), following label instructions scaled down from 100 ul to 55 ul extractions.
Whole-beetle assays
In a separate experiment, 75 beetles from an infection-free lab colony that had been starved for 48 hr were allowed to feed for 24 hr on symptomatic E. tracheiphila-infected plants and then maintained as described above. At three time points post-acquisition (0 days, 5 days, 28 days) ~25 whole beetles were ground in a Genogrinder (OPS Diagnostics, LLC) under liquid N2. DNA was extracted using 100 μl volume prepGem Bacteria following label instructions. 5 μl of this DNA was used in triplicate qPCR reactions for 18S and EtOMP as described above.
Transmission assays
To determine whether E. tracheiphila levels in frass affect transmission potential, 150 beetles were fed on two infected plants for 24 hours as described above. At 0, 5 and 28 days post exposure, frass from 100 randomly selected beetles was collected, pooled and homogenized in tap water. We applied 10 μl of this homogenate to a small incision (0.5 cm) in the leaf petioles of 14-day-old C. pepo ssp. texana seedlings with two fully expanded leaves (n = 84 seedlings at 0 days; n = 76 seedlings at 5 days, n = 54 seedlings at 28 days). Symptom development was recorded 3× weekly.
Effects of E. tracheiphila exposure on Acalymma vittatum fitness
Oviposition and survival
Seventy-five adult beetles that had eclosed within 3 days of one another were separated by sex. Half of the female beetles were placed into a mesh 1 ft × 1 ft × 1 ft Bioquip cage (cat #1466A) containing a symptomatic, E. tracheiphila-infected plant and the other half were placed in a similar cage with a healthy, mock-inoculated plant. All males fed on mock-infected (healthy) plants. In each cage, beetles were allowed to feed on infected or healthy plants for 96 hr. Individual male and female beetles were then placed together in petri dishes and allowed to mate freely over 48 hours. Following mating, females were kept individually in petri dishes with one teaspoon of moistened potting soil as an oviposition substrate and a 3 × 3 cm cucumber leaf as a food source (and transferred to new, clean petri dishes 3× weekly). Females were re-mated after 14 days to account for potential deterioration in the quality of stored sperm41. Proportion of of female survivorship and the number of eggs oviposited were recorded. Experimental insects were kept in an incubator set at 25 C with a 14:10 L/D cycle for an additional 26 days after mating.
Egg hatch rate and developmental time
In a separate experiment, newly-eclosed beetles were divided by sex and randomly assigned to feed on healthy (mock-inoculated) or infected C. pepo ssp. texana seedlings for 96 hours (as above). Individual females were then placed together with two male beetles in petri dishes and allowed to mate freely over 48 hours. Females were then kept individually in petri dishes for oviposition, as above. Thirteen females were used from each feeding treatment. All thirteen beetles that fed on symptomatic, E. tracheiphila – infected plants oviposited the first week after mating and 11 of 13 females that fed on healthy (mock – infected) wild gourd oviposited in the same period. The first 15–29 eggs laid by each female were collected from the soil in the petri dish and placed in a 1-gallon black pot in which four C. pepo ‘Raven’ seedlings had been planted as a food source; seedlings were replenished as needed. Adult emergence from each pot was recorded 3–4× weekly. Pots with seedlings and eggs were kept in a growth chamber at 25 C/22 C on a 14:10 day:night cycle.
Data deposition
Unique primer and TaqMan probe sequences developed for this project are provided above. Experimental data are archived online and available at Dryad Digital Repository (doi:10.5061/dryad.fq127).
References
Ng, J. C. K. & Falk, B. W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Ann. Rev. Phytopathol. 44, 183–212 (2006).
Purcell, A. H. & Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Ann. Rev. Phytopathol. 34, 131–151 (1996).
Garcia-Salazar, C., Gildow, F. E., Fleischer, S. J., Cox-Foster, D. & Lukezic, F. L. Alimentary canal of adult Acalymma vittata (Coleoptera: Chrysomelidae): morphology and potential role in survival of Erwinia tracheiphila (Enterobacteriaceae). Can J of Ent 132, 1–13 (2000).
Hogenhout, S. A., Ammar, E. D., Whitfield, A. E. & Redinbaugh, M. G. Insect vector interactions with persistently transmitted viruses. Annual Review Phytopathol 46, 327–359 (2008).
Madden, L. V., Jeger, M. J. & Van den Bosch, F. A theoretical assessment of the effects of vector-virus transmission mechanism on plant virus disease epidemics. Phytopathology 90, 576–594 (2000).
Hogenhout, S. A., Oshima, K., Ammar, E. L. D., Kakizawa, S. & Namba, S. Phytoplasmas: bacteria that manipulate plants and insects. Mol. Plant Pathol. 9, 403–423 (2008).
Chatterjee, S., Almeida, R. P. P. & Lindow, S. Living in two worlds: The plant and insect lifestyles of Xylella fastidiosa. Ann. Rev. Phytopathol. 46, 243–271 10.1146/annurev.phyto.45.062806.094342 (2008).
Sisterson, M. S. Effects of insect-vector preference for healthy or infected plants on pathogen spread: insights from a model. J. Econ. Entomol. 101, 1–8 (2008).
Ammar, E. D., Shatters Jr, R. G., Lynch, C. & Hall, D. G. Detection and relative titer of Candidatus Liberibacter asiaticus in the salivary glands and alimentary canal of Diaphorina citri (Hemiptera: Psyllidae) vector of citrus huanglongbing disease. Ann. Entomol. Soc. Am. 104, 526–533 (2011).
Suzuki, S. et al. Interaction between the membrane protein of a pathogen and insect microfilament complex determines insect-vector specificity. Proc. Nat. Acad. Sci. USA 103, 4252–4257 10.1073/pnas.0508668103 (2006).
Rand, F. V. & Enlows, E. M. A. Transmission and control of bacterial wilt of cucurbits. J. Agri. Res. 6, 7–434 (1916).
Brust, G. E. New economic threshold for striped cucumber beetle (Coleoptera: Chrysomelidae) in cantaloupe in the Midwest. J. Econ. Entomol. 92, 936 (1999).
Sasu, M., Seidl-Adams, I., Wall, K., Winsor, J. & Stephenson, A. Floral transmission of Erwinia tracheiphila by cucumber beetles in a wild Cucurbita pepo. Env. Entomol. 39, 140–148 (2010).
Watterson, J. C. Cucurbit responses to Erwinia tracheiphila and the multiplication and movement of the bacterium in selected resistant and susceptible cucurbits University of Wisconsin - Madison, (1971).
Sasu, M. A., Ferrari, M. J., Du, D., Winsor, J. A. & Stephenson, A. G. Indirect costs of a nontarget pathogen mitigate the direct benefits of a virus-resistant transgene in wild Cucurbita. Proc. Nat. Acad. Sci. USA 106, 19067–19071 10.1073/pnas.0905106106 (2009).
de Mackiewicz, D., Gildow, F. E., Blua, M., Fleischer, S. J. & Lukezic, F. L. Herbaceous Weeds Are Not Ecologically Important Reservoirs of Erwinia tracheiphila. Plant Dis. 82, 521–529 (1998).
Garcia-Salazar, C., Gildow, F. E., Fleischer, S. J., Cox-Foster, D. & Lukezic, F. L. ELISA versus immunolocalization to determine the association of Erwinia tracheiphila in Acalymma vittatum (Coleoptera: Chrysomelidae). Env. Entomol. 29, 542–550 (2000).
Mitchell, R. F. & Hanks, L. M. Insect frass as a pathway for transmission of bacterial wilt of cucurbits. Env. Entomol. 38, 395–403 (2009).
Leach, J. G. Observations on cucumber beetles as vectors of cucurbit wilt. Readings in insect plant-disease relationships.Vol. 54, 188–191 (1964).
SAS. Base SAS®9.3 Procedures Guides (2011).
R: A language and environment for statistical computing (R Foundation for Statistical Computing, Vienna, Austria, 2011).
Rand, F. V. & Enlows, E. M. A. Bacterial wilt of cucurbits. Vol. 828, 1–33 (US Dept. of Agriculture, 1920).
Brust, G. Seasonal variation in percentage of striped cucumber beetles (Coleoptera: Chrysomelidae) that vector Erwinia tracheiphila. Env. Entomol. 26, 580–584 (1997).
Fleischer, S. J., de Mackiewicz, D., Gildow, F. E. & Lukezic, F. L. Serological estimates of the seasonal dynamics of Erwinia tracheiphila in Acalymma vittata (Coleoptera: Chrysomelidae). Env. Entomol. 28, 470–476 (1999).
Brust, G. E. Differential susceptibility of pumpkins to bacterial wilt related to plant growth stage and cultivar. Crop Prot. 16, 411–414 (1997).
Shapiro, L., De Moraes, C. M., Stephenson, A. G. & Mescher, M. C. Pathogen effects on vegetative and floral odours mediate vector attraction and host exposure in a complex pathosystem. Ecol. Lett. 15, 1430–1438 (2012).
Elliot, S. L., Adler, F. R. & Sabelis, M. W. How virulent should a parasite be to its vector? Ecol. 84, 2568–2574 (2003).
Mauck, K., Bosque-Pérez, N. A., Eigenbrode, S. D., De Moraes, C. M. & Mescher, M. C. Transmission mechanisms shape pathogen effects on host–vector interactions: Evidence from plant viruses. Funct. Ecol. 26, 1162–1175 (2012).
Beanland, L., Hoy, C. W., Miller, S. A. & Nault, L. R. Influence of aster yellows phytoplasma on the fitness of aster leafhopper (Homoptera: Cicadellidae). Ann. Ent. Soc. Am 93, 271–276 (2000).
Mauck, K. E., De Moraes, C. M. & Mescher, M. C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Nat. Acad. Sci. USA 107, 3600–3605 10.1073/pnas.0907191107 (2010).
Broderick, N. A., Raffa, K. F. & Handelsman, J. Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proc. Nat. Acad. Sci. USA 103, 15196–15199 (2006).
Douglas, A. E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 23, 38–47 (2009).
Weiss, B. L., Wang, J., Maltz, M. A., Wu, Y. & Aksoy, S. Trypanosome infection establishment in the tsetse fly gut is influenced by microbiome-regulated host immune barriers. PLoS Pathog. 9, e1003318 (2013).
Verschuere, L., Rombaut, G., Sorgeloos, P. & Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Micro. Mol. Biol. Rev. 64, 655–671 10.1128/mmbr.64.4.655-671.2000 (2000).
Bando, H. et al. Intra-specific diversity of Serratia marcescens in Anopheles mosquito midgut defines Plasmodium transmission capacity. Sci. Rep. 3, 1641; 10.1038/srep01641 (2013).
Metcalf, R. L. & Lampman, R. L. The chemical ecology of diabroticites and cucurbitaceae. Cell. Mol. life sci. 45, 240–247 (1989).
Tallamy, D. W., Stull, J., Ehresman, N. P., Gorski, P. M. & Mason, C. E. Cucurbitacins as feeding and oviposition deterrents to insects. Env. Entomol. 26, 678–683 (1997).
McCloud, E. S., Tallamy, D. W. & Halaweish, F. T. Squash beetle trenching behaviour: avoidance of cucurbitacin induction or mucilaginous plant sap? Ecol. Entomol. 20, 51–59 (1995).
Rojas, E. S., Gleason, M. L. & Batzer, J. C. Feasibility of delaying removal of row covers to suppress bacterial wilt of muskmelon (Cucumis melo). Plant Dis. 95, 729–734 (2011).
Broackes-Carter, F. C. et al. Temporal regulation of CFTR expression during ovine lung development: implications for CF gene therapy. Hum. Mol. Gen. 11, 125–131 (2002).
Skelton, T. E. Laboratory rearing and reproduction of the spotted cucumber beetle. J. Econ. Ent. 63, 948–950 (1970).
Acknowledgements
We thank Haskell Sie and Allshine Chen for assistance with statistical analysis; Christina Grozinger for use of a qPCR machine; Dawn Luthe for use of a Genogrinder; Erin Scully for review of the manuscript and experimental advice; Erica Smyers, Janet Saunders and Christopher Balough for laboratory assistance; Nick Sloff for photography and graphic design assistance. Funding for this research was provided by Grants 2008-35302-04577 and 2009-33120-20093 from the USDA National Institute of Food and Agriculture, the David and Lucile Packard Foundation. LRS received support from a Sigma-Xi Grant-in-Aid of research, an NSF Graduate Research Fellowship and a USDA-AFRI Microbial Genomics Training grant (2010-65110-20488).
Author information
Authors and Affiliations
Contributions
L.R.S. and I.S.A. designed and executed the technical assays. L.R.S., I.S.A., C.D.M., A.G.S. and M.C.M. contributed to planning the experiments, the interpretation of the data and writing and editing the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Shapiro, L., Seidl-Adams, I., De Moraes, C. et al. Dynamics of short- and long-term association between a bacterial plant pathogen and its arthropod vector. Sci Rep 4, 4155 (2014). https://doi.org/10.1038/srep04155
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep04155
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
