
Abstract
For many years, the rise of stratified societies along the Central Andean coast, known as the birthplace of Andean civilization, has been closely linked to a marine-oriented economy. This hypothesis has recently been challenged by increasing evidence of plant management and cultivation among Andean populations long before the emergence of complex societies and monumental architecture. The extent to which marine and plant-based economies were integrated and their contributions to early sedentism, population growth, and intra-community stratification, however, remain subjects of ongoing and contentious debate. Using Bayesian Mixing Models we reanalyze the previously published stable isotopes (δ15Ncollagen, δ13Ccollagen, δ13Capatite) values of 572 human individuals from 39 archaeological sites in the Central Andes dated between ca. 7000 BCE and 200 CE to reconstruct dietary regimes in probabilistic terms. Our results reveal that fish, terrestrial fauna, and cultivated plants variably contributed to the diet of prehistoric Andean populations; in coastal and middle valley settlements plant cultivation, not fishing, fueled the development of the earliest complex societies during the Formative Period (from 3000 BCE). Similarly, in the highlands the societies that built ceremonial centers show a plant-based economy. Our findings also show that maize only became a staple food (> 25% dietary contribution) in more recent phases of Andean prehistory, around 500 BCE.
Similar content being viewed by others

Introduction
Over the past three decades, archaeological investigations have revolutionized our comprehension of the timelines and processes involved in early plant domestication along the Central Andean coast, a region renowned as the cradle of Andean civilization1,2,3,4. This transformation owes much to the meticulous analysis of plant micro- and macro-remains, which have provided invaluable insights into the chronological sequence and possible routes through which domesticated crops were disseminated throughout the region5,6,7,8. Studies indicate that during the Late Pleistocene, Early Holocene, and Middle Holocene periods, prehistoric populations in the Central Andes subsisted by gathering a diverse array of terrestrial and marine fauna. Concurrently, they initiated the management and cultivation of semi-domesticated and domesticated plants as early as 8350 BCE9,10. These food production systems were complemented by the development of irrigation technologies as early as 4550 BCE11, and camelid domestication (between 6500 and 3500 BCE)12, leading to deep economic and social changes.
Remarkably, a comprehensive review and calibration of available radiocarbon dates associated with food remains spanning from the Late Pleistocene to the Inca period have unveiled that among the 51 plants used during the time of European contact, at least 40 were under cultivation during the preceramic era, predating approximately 1800 BCE12. Overall, the emerging evidence suggests that the temporal gap between the early adoption of cultivated plants and the emergence of “full-fledged agriculture” (sensu Zeder13, an economic system primarily based on the production and reliance upon domesticates) may not have been as protracted as previously assumed,14,15, indicating that preceramic societies responsible for the early monumental centers likely practiced agriculture.
Other lines of evidence also support this hypothesis. Studies of demographic dynamics in precontact South America detected a period of exponential population growth starting around 3000–2000 BCE, linked to environmental changes and the expansion of farming systems. In the Central Andes, an increase in settlement density suggests that population growth rates were particularly high between 2000–1 BCE16. This is consistent with other empirical evidence for population increases during the Formative period (3000–1 BCE), including larger size and agglutinated distribution of settlements, as well as changes in domestic architecture17,18,19.
For the Peruvian Coast, however, the role of fishing and farming in the processes of social complexity, reflected in monumental architecture and ceremonial centers, sites hierarchization, and intracommunity stratification, which in turn may reflect the existence of large scale (regional) social cooperation or control of labor, and possibly the existence of centralized institutions1,2,15,19,20 has been widely debated15,19,20,21,22. Seminal works emphasized the marine economy as the key factor in the development of sedentism, population growth, and social stratification in the region, suggesting that social divisions would have arisen around an early centralization and strategic distribution of marine production, which would have “pre-adapted” coastal groups to a later increasingly hierarchical development based on relatively late agriculture. In this scenario, agriculture was introduced long after the initial complexity, approximately 1800 BCE, as a way of maintaining the earlier institutionalized social structures15.
Later discoveries of older and even larger monumental sites in the middle valleys have, however, reinforced the hypothesis that civilization arose inland based on an agricultural economy, rather than on the coastline. At least 35 planned sites integrated into intra-valley hierarchical systems in the North-Central Coast of Peru, with over 200 radiocarbon dates between 4100–1800 BCE, have led archaeologists to rethink the origins of urbanization processes and re-examine the chronology of the Central Andes as a whole1,23. Additionally, evidence of macro- and micro-botanical remains shows that coastal economies were more diverse and richer in domesticated plants than previously considered24,25,26. Even in models that minimize material factors as “triggers” of social stratification and emphasize behavioral causes, such as power accumulation strategies and manipulation of belief systems, the existence of subsistence systems able to support increasing populations in sedentary settlements would have played a critical role27,28.
According to recent discoveries, since at least the Initial Formative Period (3000–1 BCE) the communities of the Pacific basin valleys would have been organized around symbiotic farming-fishing economies, with trade networks along transversal littoral-inland corridors that would not typically exceed 80–100 km29. Coastal societies would have relied primarily on fishing with some dependence on cultivated products, while communities in the middle and higher sectors of the valleys cultivated semi-domesticated or domesticated plants supplemented by marine products such as dry-salted fish and mollusks11,29,30. In the highlands, the archaeological record of ceremonial centers suggests that subsistence was largely based on terrestrial food items31,32,33. However, the relative contribution of plants and terrestrial and marine protein sources to diets at different elevations and latitudes is not well understood, and their importance to the process of social complexity remains controversial29,30,34,35.
In addition, the paths of the agricultural intensification of certain plants and their role in sustaining dense populations and social structures during different stages of the process is still in discussion26,30,36. Among these, maize (Zea mays) is of particular interest. Maize began its domestication process around 9000 years ago in the Mexican lowlands37 and spread into South America around 6000 BCE where it evolved isolated from the wild teosinte gene pool38. It reached the Peruvian North Coast as early as 4800–4500 BCE8, possibly through multiple waves of colonization39,40. Until recently, the Central Andes was considered a “2nd center of domestication for maize”36, but currently is recognized as an “improvement center” where local adaptations of partially domesticated maize occurred38. While maize’s importance as a staple food has been demonstrated in later periods, linked to state and empire domination strategies (e.g., as maize beer offered during community work, feasts, and ceremonies)12,41,42, its significance as a prime mover of the earliest complex societies remains unclear30,31,43,44.
Available evidence suggests that its consumption may have been limited in the region, possibly only in ceremonial contexts or as a seasonal crop, until later times45,46. Stable carbon and nitrogen isotope analyses of collagen (δ13Ccoll and δ15N) and bioapatite (δ13Cap) from exhumed human bones and teeth have proven to be an unparalleled method to study subsistence in past populations47. The δ13Ccoll is a proxy for dietary protein sources and photosynthetic pathways of plants in the food web (C3, C4, CAM). The δ13Cap instead reflects the isotopic composition of all macronutrients (carbohydrates, lipids, proteins) in an individual’s diet, offering insights into the energetic constituents48. The analysis of δ13Ccoll and δ13Cap can effectively discriminate diets based on C3 from C4 (i.e., maize and amaranth, a high altitude plant12) and CAM plants (cactuses with isotope values that overlap C4 in the Andean case) plants due to fundamental differences in their isotopic composition49,50; however, they are not suitable for distinguishing between diets based on marine resources and those based on C4 plants, due to overlapping values51. This limitation is usually addressed by analyzing the δ15N value since it is affected by relatively well-established trophic fractionations (3–6‰), which are larger in aquatic systems due to longer and more complex food webs compared to terrestrial systems52. High δ15N values, however, can be also related to aridity53 and fertilization practices (e.g., camelid dung, seabird guano)54,55, two factors to be considered in isotopic analyses in the Central Andes.
Stable isotope analyses have been successfully employed to reconstruct diets in prehistoric Andean populations, particularly for later periods, during which the main dietary trends and shifts have been identified17. However, the evidence for earlier periods, when the first complex societies arose, is more limited30,34,44,46. Despite the excellent morphological preservation of human remains and the growing interest in the relationship between subsistence systems and social changes, efforts to understand dietary shifts at the individual or population level have been hindered due to the lack of viable collagen from prehistoric populations beyond the first millennium CE30,56,57,58. Furthermore, until recently, the analytical methods used only allowed for a qualitative approach to the dietary constituents, which did not clarify in quantitative terms the contribution of C3 plants (e.g., tubers, legumes) and C4 plants such as maize, nor the contribution of marine and terrestrial proteins in these early diets.
Here, we reassess the stable carbon and nitrogen isotope values of 572 pre-Columbian human individuals from Peru. These individuals represent 61 populations dated between ca. 7000 BCE and 200 CE, through the Archaic (7000–3000 BCE), Formative (3000–1 BCE) and Early Intermediate (1–600 BCE) periods, and were recovered from 39 archaeological sites located along the coast, middle valleys, and highlands of the three distinct latitudinal regions (i.e., North, Central, and South) of the Central Andes, characterized by contrasting environmental conditions (Fig. 1, see Supporting Information 1). We also included isotope values from 408 faunal and 404 plant specimens discriminated by subregion, to characterize regional and local food webs (Supporting Information 2).

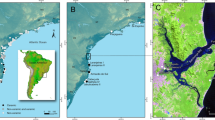
Location of the archaeological sites mentioned in the text and tables. Map generated with QGIS v. 3.2.8 (https://www.qgis.org) with images of Google Earth Pro® (2023 Google LLC. All rights reserved) and a raster basemap from https://www.naturalearthdata.com/.
A Bayesian stable isotope mixing models (BSIMMs) approach was used to estimate the proportional caloric contributions of marine and terrestrial fauna, as well as C3 and C4 plants, to the diets of individuals from different sites and periods30,59,60,61 to assess their economic importance for Andean communities over the Archaic and Formative periods and refine the chronology of major dietary shifts. Out of 572 individuals, 293 individuals with confident parameters for collagen preservation62,63 and a complete set of isotope values (i.e., δ13Ccoll, δ15N, and δ13Cap) were included in the BSIMMs reconstructions (see Material and Methods and Supporting Information 3).
Our aim is to determine whether major social changes, demographic growth, and political-ideological transitions, which usually define the archaeological periodization, are linked to the intensification of key crops, intended as the substantial increased intake of domestic plants. While our primary focus is to elucidate the role of marine and terrestrial resources on the Andean coast, highland diets have been included to document different trajectories in crop adoption along this vast and heterogeneous environment.
Results
Stable isotope analysis and Bayesian dietary reconstruction
In the available dataset, human individuals had δ13Ccoll values (n = 317) ranging from − 23.8‰ to − 9.1‰, δ15N values (n = 294) from + 2.8 ‰ to + 24.9 ‰, and δ13Cap values (n = 400) from − 15.7 ‰ to − 2.4‰. The δ13Ccoll and δ15N values of coastal populations fall within the range of expected values for marine based diets, notably during the Archaic period (7000–3000 BCE), whereas inland and highland populations show values consistent with diets based on C3 plant ecosystems (Fig. 2a). Some populations, however, show wide standard deviations suggesting quite diverse diets. As expected, human δ15N values decrease significantly with increasing distance from the sea (n = 292; Spearman’s ρ = − 0.693; p < 0.01).

(a) Average δ13Ccoll and δ15N values from prehistoric populations of the Central Andes according to region. (b) Average δ13Ccoll and δ13Cap values from human individuals in the workspace of Kellner and Schoeninger48, discriminated by region for comparative purposes. In this model, the regression lines represent the source of protein (C3, marine, and C4) and the location of the values along the lines indicate the relative proportion of energy provided by the source. Red dots represent highland populations. Dots represent averages and crosses are standard deviations (1σ).
The δ13Ccoll and δ13Cap values (Fig. 2b), which track the sources of protein and energy components of the diet, respectively, show mixed subsistence strategies and three different dietary regimes through time: 1) the diets of the Initial, Early and Middle Formative populations (3000–600 BCE) with a significant contribution of C3 plants (i.e., legumes, tubers, and cucurbits) and a more discrete contribution of C4 plants (maize and potentially amaranth, depending on ecological setting); 2) the diets of Late Formative populations (600–1 BCE) based on C3 plants with a growing consumption of C4; and 3) diets based on a higher intake of C4 plants and less C3 plants, mostly linked to populations dated between 200 BCE and the first centuries of the current era.
It is worth noting that while C4 plant intake varied considerably among early populations (for instance, an early dependence on C4 plants appears evident in the North Coast since at least 4500 BCE44), their consumption increased significantly in later populations of the North and Central coasts, especially during the last half of the first millennium BCE (p < 0.05 with Kruskal–Wallis-H for δ13Cap; see post-hoc tests in Supporting Information 1: SI1 T4).
In conclusion, estimates from BSIMMs indicate that, in general, C3 plants were the main sources of dietary calories for most of the populations during the Formative periods, and notably in the highlands. Marine resources provided most proteins to coastal populations, decreasing inland and in the highlands. Calories from maize, instead, increased over time, notably among coastal and inland populations from the North and Central regions during the Late Formative and the Early Intermediate (Fig. 3; see also Supporting Information 4). However, we acknowledge that his interpretation is mostly based on five outlier populations, thus, further studies are required to confirm this hypothesis.

Dietary caloric estimations of food sources in Andean populations grouped by geographic regions, and archaeological periods: Archaic (Arch); Formative; Early Intermediate. The boxes represent a 68% confidence interval (between the 16th and 84th percentiles) and the whiskers a 95% confidence interval (corresponding to the 2.5th and 97.5th percentiles). The horizontal discontinuous line indicates the median (50th percentile). The horizontal continuous line indicates the mean. Sites’ IDs in black refer to sites or phases with monumental architecture or hierarchical distribution of settlements. IDs in red refer to non-complex sites (see also SI1 and SI4 F1and F2).
Discussion
Regional dietary trends
In the Central Andes, the role of agriculture and fisheries as economic drivers for sedentism, population growth and social stratification has been widely debated, and a wide variability in diets across time periods is observed in different subregions. Coastal and highland populations show more reliance on marine products and C3 plants, respectively. However, coastal diets show also high reliance on C3 plants. Among inland groups, a notable overlap of isotope values suggests more diverse diets based on plants.
On the North Coast, maize was an important dietary item at the site of Paredones as early as 4500 to 4000 BCE, whereas in the neighboring site of Huaca Prieta people relied on marine foods complemented with terrestrial items and a little maize around 3000–2500 BCE8,10,44,64. From the Early Formative (1200–1000 BCE) to the Salinar period (400–200 BCE), communities flourished with a combination of C3 plants, marine and terrestrial protein, with low C4 contribution (< 15%). Archaeological inventories and coprolite analyses indicate diets of C3 plants (i.e., tubers, beans, peanuts, and carob), fish (e.g., Squatina armata, Mustelus sp., Paralonchurus sp., Cynoscion sp., Sciaena deliciosa), marine mammals (Otaria sp.) and shellfish (e.g., Mesodesma donacium, Choromytilus chorus, Polinices uber, Thais sp.), and some maize57,65. Increasing maize contribution, making up more than 60% of the dietary calories, is clear in inland sites (Viru 11, Viru 59) around 200 BCE–200 CE, during the Gallinazo period, between the Late Formative and the Early Intermediate57,66.
In the Northern highlands, diets were dominated by C3 plants over the evaluated periods (Fig. 3). This scenario of C3 preference (i.e., quinoa, beans, and various tubers) can be extended regionally to other Formative sites with monumental architecture, such as Huaricoto (~ 2200–200 BCE), Huacaloma (1500–550 BCE), Pacopampa (900–600 BCE), Chavin (850–450 BCE), and Kuntur Wasi (800–1 BCE). Maize increases slightly only at the beginning of the current era31,32,33,46,56. Terrestrial protein was potentially more important at early sites, like La Galgada, possibly due to easily accessible cervids or camelids32. Later, diets were low protein, despite being herders, like in Pacopampa33.
On the Central Coast, Archaic populations from La Paloma and Chilca I show the greatest dependence on marine and terrestrial fauna among all the compared groups34,67 (Fig. 3). Although macro- and micro-botanical remains (phytoliths, starches, and pollen) from Fortaleza, Pativilca, and Supe valleys dated between 2900 and 2100 BCE, confirm maize consumption during the Initial Formative6,25,26, isotopic analyses suggest that its consumption was limited and sporadic, with Formative sites instead showing a reliance on C3 plants. For instance, Caral individuals during the Initial Formative showed dietary calorie contributions of 62% C3 plants and less than 7% maize, while diets from Áspero and Bandurria30,68, two early settlements with monumental architecture located on the shoreline, had contributions of 50–60% C3 plants. C3 plant diets predominated the Early and Middle Formative, but these preferences changed in the Late Formative with maize contributions increasing by 10% to 25%30. For example, in Tablada de Lurin (200 BCE–100 CE), based on combination of C4 plants and marine resources, maize provided around 40–50% of dietary calories56,58.
In the Southern region, during the Late Formative period, individuals from Paracas communities (550–100 BCE), had a diet composed of C3 plants (56–57% of dietary calories), mainly tubers (manioc, jíquima, achira, among others69), marine resources (19–21% of dietary calories), and maize (11–14% of dietary calories). Individuals from Samaca cemeteries (100 BCE–100 CE) in the lower Ica Valley, show dietary preference for C3 plants (i.e., peanuts, squash, manioc, and pacay) and low amounts of C4 plants (i.e., maize, and possibly CAM plants), and restricted marine foods despite a relatively short distance to the sea70. In this region, maize intensification seems to have been delayed despite the proximity to Ayacucho and Cusco in the South Highlands, where maize cultivation was hegemonic at least since 800 BCE71,72.
The compiled isotope data reveals dietary trends during the Archaic period (7000–3000 BCE), showing that coastal individuals had diverse diets primarily composed of marine resources and C3 plants (e.g., tubers, cucurbits, and legumes), alongside terrestrial mammals, which aligns with faunal remains30,67. In contrast, highland individuals primarily relied on C3 plants and terrestrial mammals during this period. Moving into the Formative Period (3000–1 BCE), coastal, inland, and highland populations increasingly incorporated C3 plants into their diets, although coastal communities continued to consume fish. Maize played a limited role during the Formative Period, with notable exceptions like Paredones. Maize consumption saw a notable rise during the first millennium BCE, a period linked to increasing population pressure16,17,30, contributing to approximately 25% of calories in some inland sites by 500 BCE. Its intensification was more marked on the North Coast (between 400–100 BCE) and the Central Coast (~ 200–1 BCE), and slower on the South Coast (between 750–450 BCE)73. It eventually became a staple between 200 BCE and 200 CE in the North and Central coasts. This increase in maize production and consumption during the last half of the first millennium BCE is probably linked to chicha production for ceremonial activities74 and is the precursor to the extensive production and use of chicha in Early Intermediate Period and Middle Horizon contexts75.
While archaeological and botanical evidence strongly supports maize as the primary C4 plant in the Andes, it is important to note that maize was not the exclusive source of 13C-enriched values in Andean diets. The overlapping δ13C values of maize, amaranth, and CAM plants (i.e., cactuses) is a confusing factor that should be considered in the interpretation of maize adoption in arid and high-altitude environments. However, the available evidence suggests that cactuses were more important for Archaic populations, and amaranth, as staple, possibly was restricted to highlands or lower latitudes12,70,76.
This extended process of farming intensification among Andean communities involved the implementation of productive strategies that were well-suited to the specific crops, soils, and limited water resources in these arid and challenging landscapes (i.e., subterranean water management, increasing irrigation technology, sunken fields, flooding agriculture, consortium cultivation etc.77), modulated by changing environmental conditions16,78,79,80. These technological improvements probably did not occur at the same time and not necessarily were related to the same factors in each subregion. Social and political structures could have variably affected the way that crops were adopted and used by populations from different altitudes, characterizing particular processes for each region and site.
The factors that conditioned maize intensification by a few “maize-dependent” populations and preclude its adoption by most Formative communities are possibly related to ecological (e.g., water availability, land features, climate, and altitude restrictions) and cultural (e.g., identity) circumstances. For instance, the wider valleys of the North Coast, especially those with permanent water supply are more appropriate for maize cultivation with insipient technology20.
To conclude, this analysis helps address three foundational questions about the diet in the Archaic and Formative periods of the Central Andes: 1) the diet that led to the development of societies linked to monumental architecture, presumably complex societies, was not predominantly marine in caloric terms; 2) these developments were based mainly in C3 plants at any altitude, including coastal sites; and 3) most diets were not based on maize, which, in turn, was a crop of marginal importance at least until the first millennium BCE. Thus, in accordance with previous arguments, for coastal settings, our results challenge the prevailing theory that marine resources were the primary economic driver for social complexity. Our results, instead, align well with the emerging view that political development in most inland and highland sites relied on productive systems based on C3 crops. It emphasizes the significance of plant cultivation in propelling population growth, social stratification, and the development of political institutions in one of the cradles of civilization.
Methods
Archaeological sites and individuals
This is an in-silico study developed using previously published stable isotope data hosted in the South American Archeological Isotopic Database (SAAID_V.2.0_2023, hosted in CORA Platform—https://doi.org/10.34810/data602). A total of 572 human individuals from 39 archaeological sites of the Central Andes, which represent the totality of the available data before the current era, were included in our analysis.
The database with raw isotope values of individuals (consumers’ values) is reported in the Supporting Information 1. Sex and age estimates are reported as in the original references. Most of the sites are in the Peruvian valleys that transversally cross the Pacific Coastal Desert to drain in the Pacific Ocean, at coastal (i.e., less than 3 km from the sea; n = 10), and inland (i.e., lower, and middle valleys < 600 m above the sea level, located at any distance from the coast; n = 16), locations. Other sites (n = 13) are in the Andean highlands (i.e., upper valleys of the Pacific Basin with altitude > 600 masl, and interandean valleys at any altitude). Because some of the sites have long occupational sequences, a total of 61 discrete populations were analyzed (see Supporting Information 1). Sites and phases were classified by social complexity in two categories: 1) sites with monumental architecture or ceremonial centers, which correspond to more or less complex societies; and 2) other type of sites (camps, towns, open sites, rock shelters), that possibly correspond to non-complex or egalitarian societies. This classification was contrasted with the archaeological interpretations for each site based in several markers (i.e., architectural features, construction volume, intravalley settlement patterns, mortuary evidence, and archaeological inventories) and corrected when necessary (e.g., Paracas Peninsula sites, which are tombs linked to a complex settlement network and rich mortuary offerings).
The chronological framework of the sites and cultural phases were based on re-calibrated radiocarbon dates provided in the references (see details in Supporting Information 3). The time span of examined individuals comprises the Archaic (7000–3000 BCE), Formative (3000–1 BCE), and Early Intermediate (1–600 BCE) periods.
Our analysis also included isotope values from 812 specimens of fauna (n = 408) and plants (n = 404) for food web reconstructions (see Supporting Information 2). These values were discriminated by latitudinal region to generate more reliable inputs for Bayesian models and visual interpretations. An isotopic baseline of potential dietary sources by each sub-region was generated with rKin using Standard Ellipse Areas81 (see Supporting Information 3). All the isotope values were extracted from SAAID.
To prevent the potentially confusing effect of taphonomy issues in our analysis, only individuals with reliable δ13Ccoll and δ15N values from bone and dentine collagen, according to accepted preservation criteria62,63, were included. For δ13Cap values, although the volume of published data is higher than collagen, preservation criteria have not been used extensively, especially in older studies, and the assumption of reliability has been largely based on the extraction protocols82. In this research, we used bone δ13Cap values only if a sample’s collagen was well-preserved. We assumed that if collagen, an organic biomolecule that forms the biological structure housing inorganic crystals, is well-preserved, then bioapatite is also well-preserved. However, apart from data consistency (the values in apatite are typically 13C-enriched compared to collagen), there are no other objective markers to assess apatite preservation83. Because dental enamel preserves better than bone, all available enamel δ13Cap values from selected individuals were employed.
To avoid confounding factors of breastfeeding and weaning, just adults with acceptable collagen atomic composition were included in our analyses. Infants and toddlers (subadults < 4 years) were not included84. According to these criteria, 293 human individuals with confident parameters for collagen preservation and a complete set of isotope values (i.e., δ13Ccoll, δ15N, and δ13Cap) were selected for BSIMMs analyses.
Isotopic analysis and BSIMMs
First, we used descriptive statistics and conventional isotope scatterplots as diagnostic tools to identify major dietary trends and shifts for each population and region. BSIMMs were then run for each group. Scatterplots of δ15N vs. δ13Ccoll provide information about the type of protein consumed, terrestrial (nitrogen below + 11 ‰ and carbon below − 12.5 ‰), or marine if both values are very high52. Scatterplots of δ13Ccoll vs. δ13Cap in a workspace of three regression lines derived from the experimental feeding studies, allows for discrimination of the protein and carbohydrate (C3 or C4) sources. The distribution of an individual’s values on the axis of the apatite indicates if the energy of the diet is C3 or C4, and in the axis of collagen, if the protein source is C3 or C4/marine. Populations concentrated between the two lines would have complex diets, combining C3 and C4 proteins. The distance to the end lines indicates the approximate proportion of C3 or C4 energy consumed48.
Descriptive statistics (mean, range, standard deviation, standard error of the mean) and comparisons of isotope values between groups, were used to detect diagnostic differences over time and space. These analyses were performed using Kruskal–Wallis tests (α = 0.05), after evaluation for normal distribution with Kolmogorov–Smirnov test for normality (α = 0.05). Mann–Whitney U (α = 0.05) was used as a post-hoc test, accordingly. Spearman’s correlation was used to test the relationships between isotope values and distance to the sea. All the tests were performed in SPSS v.21 (Microsoft®).
The wide range of dietary variations in macronutrient composition, isotopic values of consumed items, fractionation (diet-to-consumer offsets), and nutrient routing factors, along with other sources of uncertainty, often hinder accurate dietary reconstructions using graphic or linear methods alone. In contrast, BSIMMs allow for the inclusion of multiple proxies and sources of variation, enabling more reliable estimates of dietary composition as probability distributions59,60,61.
The software Food Reconstruction Using Isotopic Transferred Signals (FRUITS v.2.1.1 Beta program), was used to estimate the proportional calorie contribution of different food sources to human diets, based on stable isotope values as dietary proxies59. A weighted model with fractionation factors (offsets) based on nutrient concentrations in the respective food groups was applied60,61.
The consumer data for individuals consist of their isotopic values with an uncertainty of ± 0.560. For groups, we used the average isotope values of each period/site and the standard error of the mean. After a careful screening of preservation criteria for collagen in the SAAID database, only individuals with valid collagen isotope values from each period/site were included in the analysis. When available, a complete set of δ13Ccoll, δ15Ncoll and δ13Cap values was included in models of three proxies. However, because some individuals/populations had only δ13Ccoll, δ15Ncoll values, we were required to implement alternative models with two proxies for comparative purposes. Two-proxy models can reflect the main trends in dietary compositions but may be less confident in the exact proportions of the analyzed food sources compared to three-proxy models30,59,60.
The potential assimilated food sources (Food groups), with their respective composition of macro-nutrients (Food fractions: bulk, protein, and energy) were grouped in four categories: terrestrial faunal (providing proteins and lipids), called “TF” in the model; marine faunal (providing proteins and lipids), called “MF”; and C3 and C4 plants (providing carbohydrates and proteins), called C3 and C4, respectively30,59,60,61.
The isotope values from each food group come from isotopic paleodietary studies focused on the Central Andes valleys classified in three latitudinal regions: the Central Coast and Central Highlands, the North Coast and Northern Highlands, and the South Coast and Southern Highlands. For consistency, each site was analyzed in accordance with isotopic data from regional or local species, paying special attention to a site’s archaeological inventories or previous isotope values from local modern or archaeological samples. Although this implies the omission of some valuable references, each region was characterized with its own values. If local or regional values were not available, values from neighboring regions were used. For comparisons with the archaeological values, δ13C values from modern specimens were corrected for the fossil fuel effect, adjusting the values by + 1.5 ‰85.
For marine fauna, a synthesis of several sources was used, mainly based on modern data from the Central Coast30. Only isotope values from modern specimens were used for plants to avoid issues related to preservation86,87. The average δ13C and δ15N of food groups, the number of specimens, and species included in the models, as well as the original reference of the values for the Central, North, and South regions, can all be found in the Supporting Information 2.
Isotope fractionation factors are consensual values derived from experimental studies. For δ13C, the fractionation factor was set at + 4.8 ± 0.5 ‰ between diet and collagen, and + 10.1 ± 0.5 ‰ between diet and apatite. The δ15N diet-collagen fractionation factor was set at + 5.5 ± 0.5 ‰59,60,61,88.
The weighted values of each fraction of macronutrients (lipids, carbohydrates, and proteins) were set based on previously published parameters30,59,60,61. The values of δ13Cap represent the total carbon mix in the diet. Therefore, we use the same bulk value for each food group56,57,58. Terrestrial and marine fauna δ13C bulk values were estimated as a weighted mean of lipid and protein δ13C values59,60 For collagen, the carbon of the protein and energy routed to the total collagen was established at 74 ± 4% and 26%, respectively89, although carbon can be routed from carbohydrates and lipids recycled during the synthesis of non-essential amino acids90. We assumed that nitrogen was derived exclusively from proteins (100%). Lipids and carbohydrates were added to the model as “energy”. In order to integrate this combinatory effect, we applied a “concentration-dependent” model59,60,61.
Following previous reconstructions using FRUITS30,59,60,61, the isotopic composition of each nutritional fraction (protein, carbohydrates, and lipids) was obtained from the mean values of δ13Ccoll and δ15Ncoll using the following fractionation factors: − 2‰ (∆13Cprotein-collagen), − 8‰ (∆13Clípids-collagen), and + 2‰ (∆15Nprotein-collagen) for terrestrial mammals, and − 1‰ (∆13Cprotein-collagen), − 7‰ (∆13Clípids-collagen), and + 2‰ (∆15Nprotein-collagen) for marine animals. For plants, the offsets were − 2‰ (∆13Cbulk-protein) and + 0.5‰ (∆13Cbulk-lipids), while for the δ15N value of plant protein, the known value of δ15N recorded for the plant was assumed. The carbon weight (concentrations) of each food fraction (protein and energy) from each food group was calculated according to its macronutrient composition30,59,60,61 (see details in Supporting Information 3).
Finally, the dietary estimates were parametrized by a physiological, conservative, and acceptable range of protein consumption stipulated between 5 and 45% of the total calories59. This input was charged as “prior”. Guided by our observations of the bivariate plots, alternative models were tested for each site and population. The estimates generated for FRUITS models reflect carbon content or equivalent calorie contributions and are expressed as relative contributions (adding up to 1 or 100%) with an associated 1σ uncertainty60. The estimations of food group contributions are reported in the Supporting Information 4 (see also Supporting Information 5).
Study limitations
It is necessary to objectively judge the limitations of this study and acknowledge the inherent complexity of economic and sociocultural processes in the Andean region. These processes go beyond what can be captured through dietary reconstructions alone. As new research emerges, the current understanding of the topic may evolve, and we must remain open to revising our interpretations based on additional findings.
In addition, this study uncovered some complicating factors that may have affected the overall data interpretations: 1) information to asses bone collagen preservation according to established collagen quality criteria62,63 were not always available in the surveyed literature; 2) several ancient Andean populations have poor collagen preservation, thus not providing sufficient bone collagen for dating or isotopic analyses, and considerable temporal and regional gaps were observed in the sampled isotope data; 3) the sites contained different numbers of human individuals, which may impact the observed variability; 4) some sites lack information on faunal and botanical remains, and the data preclude the construction of local isotope baselines therefore limiting the possibility of obtaining refined dietary estimates; 5) most of studies from the past three decades included in this research did not have access to the methods currently used for analyzing bioapatite integrity (i.e., Raman spectroscopy and FTIR)83, thus, enamel and bioapatite diagenesis cannot be estimated, additionally, there is a potential issue with the interlaboratory variation of δ13Cap analysis91; 7) despite the accuracy of the methods employed, we should consider isotopic equifinality, that is, varying combinations of food contributions that may result in the same consumer isotopic values; finally, 8) isotopic markers are unable to differentiate cultivated from non-cultivated plants, which is central in this discussion, however, based on available references2,4,7,10,12,26, we can assume, that most plants behind the isotopic values observed in this study were domesticated crops.
Data availability
All data that support paper’s conclusions are present in the main text and the Supporting Information files. The stable isotope data used in this research have been previously published and were obtained in compliance with the regulations of the Peruvian government (i.e., Ministerio de Cultura since 2010 and, previously, by the Instituto Nacional de Cultura). This research did not involve direct processing of human tissues, and there are no ethical issues related to the handling of published data.
References
Shady Solís, R., Haas, J. & Creamer, W. Dating Caral, a Preceramic urban center in the Supe Valley on the central coast of Peru. Science 292, 723–726 (2001).
Haas, J. & Creamer, W. Crucible of Andean Civilization: The Peruvian Coast from 3000 to 1800 BC. Curr. Anthropol. 47, 745–756 (2006).
Dillehay, T. D. Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru (University of Texas Press, 2017).
Dillehay, T. D. From Foraging to Farming in the Andes: New Perspectives on Food Production and Social Organization (Cambridge University Press, 2011).
Grobman, A. et al. Preceramic maize from Paredones and Huaca Prieta. Peru. Proc. Natl. Acad. Sci. USA 109, 1755–1759 (2012).
Haas, J. et al. Evidence for maize (Zea mays) in the Late Archaic (3000–1800 B.C.) in the Norte Chico region of Peru. Proc. Natl. Acad. Sci. USA 110, 4945–4949 (2013).
Bonavia, D., et al. Plant remains in Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru (ed. Dillehay, T. D.) 367–433 (University of Texas Press, Austin, TX, 2017).
Prieto, G. The fisherman’s garden: Horticultural practices in a second millennium maritime community of the North Coast of Peru in Maritime Communities of the Ancient Andes (eds. Prieto, G., & Sandweiss, D. H.) 218–246 (University Press of Florida, 2020).
Dillehay, T. D., Rossen, J., Andres, T. C. & Williams, D. E. Preceramic adoption of peanut, squash, and cotton in northern Peru. Science 316, 1890–1893 (2007).
Piperno, D. R. & Dillehay, T. D. Starch grains on human teeth reveal early broad crop diet in northern Peru. Proc. Natl. Acad. Sci. USA 105, 19622–19627 (2008).
Dillehay, T. D., Elling, H. & Rossen, J. Preceramic irrigation canals in the Peruvian Andes. Proc. Natl. Acad. Sci. USA 102, 17241–17244 (2005).
León, E. 14,000 años de alimentación en el Perú (Universidad de San Martin de Porres, 2013).
Zeder, M. A. Core questions in domestication research. Proc. Natl. Acad. Sci. USA 112, 3191–3198 (2015).
Dillehay, T. D. et al. Emergent consilience among coeval fishing and farming communities of the middle holocene on the North Peruvian coast. Front. Earth Sci. 10, 939214 (2022).
Moseley, M. E. The Maritime Foundations of Andean Civilization (Cummings, Menlo Park, CA, 1975).
Goldberg, A., Mychajliw, A. M. & Hadly, E. A. Post-invasion demography of prehistoric humans in South America. Nature 532, 232–235 (2016).
Wilson, K. M. et al. Climate and demography drive 7000 years of dietary change in the Central Andes. Sci. Rep. 12, 2026 (2022).
Willey, G. Prehistoric settlement patterns in the Virú valley, Peru (Bureau of American Ethnology Bulletin 155, Smithsonian Institution, Washington DC, 1953).
Sandweiss, D. H. Early fishing and inland monuments. In Andean Civilization. A tribute to Michael Moseley (eds. Marcus, J., & Williams, P.R.) 39–54 (Cotsen Institute of Archaeology, University of California, Los Angeles, CA, 2009).
Engel, F. A. De las Begonias al Maíz. Vida y Producción en el Perú Antiguo (Centro de Investigaciones de Zonas Áridas CIZA, Universidad Nacional Agraria La Molina, Lima, 1987).
Raymond, J. S. The maritime foundations of Andean civilization: A reconstruction of the evidence. Am. Antiq. 46, 806–821 (1981).
Makowski, K. Pre-Hispanic Andean Urbanism and its ‘Anti-Urban’ Peculiarities. J. Urban Archaeol. 8, 165–196 (2023).
Creamer, W., Ruiz, A., Perales, M. & Haas, J. The Fortaleza Valley, Peru: Archaeological Investigation of Late Archaic Sites (3000–1800 BC). Fieldiana Anthropol. 44, 1–108 (2013).
Vega-Centeno, R. Ritual and Architecture in a Context of Emergent Complexity: A Perspective from Cerro Lampay, a Late Archaic Site in the Central Andes (Ph.D. Dissertation, The University of Arizona, Tucson, AZ, 2005).
Shady, R. Los orígenes de la civilización y la formación del Estado en el Perú: Las evidencias arqueológicas de Caral-Supe. In La Ciudad Sagrada de Caral-Supe: Los orígenes de la civilización andina y la formación del Estado Prístino en el antiguo Perú (eds. Shady, R. & Leyva, L.) 93–105 (Instituto Nacional de Cultura, Lima, 2003).
Yseki, M., Pezo-Lanfranco, L., Machacuay, M., Novoa, P. & Shady, R. Plant consumption in Áspero, Peru, during the Initial Formative Period (3000–1800 BCE): New evidence from starch grain trapped in human dental calculus. Sci. Rep. 13, 14143 (2023).
Rick, J. W. The Evolution of Authority and Power at Chavín de Huántar, Peru. In Foundations of Power in the Prehispanic Andes (eds. Vaughn, K. J., Ogburn, D. & Conlee, C.A.) pp. 13–36 (Archeological Papers of the American Anthropological Association Vol. 14, Arlington, VA, 2005).
Moore, J. Power and Practice in the Prehispanic Andes: Final Comments. In Foundations of Power in the Prehispanic Andes (eds. Vaughn, K. J., Ogburn, D. & Conlee, C.A.) 261–274 (Archeological Papers of the American Anthropological Association Vol. 14, Arlington, VA, 2005).
Shady, R. El sistema social de Caral y su trascendencia: El manejo transversal del territorio; la complementariedad social y política; y la interacción intercultural, Nayra Kunan Pacha, 19–90 (2018).
Pezo-Lanfranco, L. et al. The diet at the onset of the Andean Civilization: New stable isotope data from Caral and Áspero, North-Central Coast of Peru. Am. J. Biol. Anthropol. 177, 402–424 (2022).
Seki, Y. & Yoneda, M. Cambios de manejo del poder en el Formativo: desde el análisis de la dieta alimenticia. Perspectivas Latinoamericanas 2, 110–131 (2005).
Washburn, E. et al. Maize and dietary change in early Peruvian civilization: Isotopic evidence from the Late Preceramic Period/Initial Period site of La Galgada, Peru. J. Arch. Sci. Rep. 31, 102309 (2020).
Takigami, M. et al. Isotopic study of maize exploitation during the Formative Period at Pacopampa. Peru. Anthropol. Sci. 129(2), 121–132 (2021).
Beresford-Jones, D. G. et al. Diet and lifestyle in the first villages of the middle preceramic: Insights from stable isotope and osteological analyses of human remains from Paloma, Chilca I, La Yerba III, and Morro I. Lat. Am. Antiq. 32, 741–759 (2021).
Beresford-Jones, D. G. et al. Refining the maritime foundations of Andean Civilization: How plant fiber technology drove social complexity during the preceramic period. J. Arch. Method Theory 25, 393–425 (2018).
Bonavia, D. E. maíz, su origen, su domesticación y el rol que ha cumplido en el desarrollo de la cultura (Universidad de San Martín de Porras, 2008).
Piperno, D. R., Ranere, A. J., Holst, I., Iriarte, J., & Dickau. R. Proc. Natl. Acad. Sci. USA 106, 5019–5024 (2009).
Kistler, L. et al. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 362, 1309–1313 (2018).
Moreno-Mayar, J. V. et al., Early human dispersals within the Americas. Science 362, eaav2621 (2018).
Nakatsuka, N. et al. A paleogenomic reconstruction of the deep population history of the Andes. Cell 181, 1–15 (2020).
Rostworowski, M. Pescadores, artesanos y mercaderes costeños en el Perú prehispánico in Obras Completas III (Instituto de Estudios Peruanos IEP, Lima, 2004).
Tung, T. A. & Knudson, K. J. Stable Isotope analysis of a pre-hispanic Andean Community: Reconstructing Pre-Wari and Wari Era diets in the Hinterland of the Wari Empire, Peru. Am. J. Phys. Anthropol. 165, 149–172 (2018).
Perry, L. et al. Early maize agriculture and interzonal interaction in southern Peru. Nature 440, 776–779 (2006).
Tung, T. A., Dillehay, T. D., Feranec, R. S. & DeSantis, L. R. G. Early specialized maritime and maize economies on the North Coast of Peru. Proc. Natl. Acad. Sci. USA 117, 32308–32319 (2020).
Shady, R. Caral-Supe and the north-central area of Peru: The history of maize in the land where civilization came into being in Histories of maize: Interdisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize (eds. Staller, J. E., Tykot, R. H., & Benz, B. F.) 381–402 (Elsevier, New York, 2006).
Burger, R. & van der Merwe, N. Maize and the origin of highland Chavín civilization: An isotopic perspective. Am. Anthropol. 92, 85–95 (1990).
Richards, M. Isotope Analysis for Diet Studies. In Archaeological Science: An Introduction (eds. Richards, M. & Britton, K.) 125–144 (Cambridge University Press, Cambridge, 2020).
Kellner, C. M. & Schoeninger, M. J. A simple carbon isotope model for reconstructing prehistoric human diet. Am. J. Phys. Anthropol. 133, 1112–1127 (2007).
Ambrose, S. H., & Norr, L. Experimental evidence for the relationship of carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate in Prehistoric Human Bone: Archaeology at the Molecular Level (eds. Lambert, J., & Grupe, G.) 1–37 (Springer, Berlin, 1993).
Lee-Thorp, J. A., Sealy, J. C. & van der Merwe, N. J. Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationship to diet. J. Archaeol. Sci. 16, 585–599 (1989).
Chisholm, B. S. Variation in diet reconstructions based on stable carbon isotopic evidence in The Chemistry of Prehistoric Human Bone (ed. Price, T. D.) 10–37 (Cambridge University Press. New York, 1989).
Schoeninger, M. J., DeNiro, M. J. & Tauber, H. Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220, 1381–1383 (1983).
Schwarcz, H. P., Dupras, T. & Fairgrieve, S. 15N enrichment in the Sahara: In search of a global relationship. J. Archaeol. Sci. 26, 629–636 (1999).
Szpak, P., Millaire, J. F., White, C. D. & Longstaffe, F. Influence of seabird guano and camelid dung fertilization on the nitrogen isotopic composition of field-grown maize (Zea mays). J. Archaeol. Sci. 39, 3721–3740 (2012).
Santana-Sagredo, F. et al. ‘White gold’ guano fertilizer drove agricultural intensification in the Atacama Desert from AD 1000. Nat. Plants 7, 152–158 (2021).
Tykot, R. H., Burger, R. L., & van der Merwe, N. J. Importance of Maize in Initial Period and Early Horizon Peru in Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize (eds. Staller, J. E., Tykot, R. H., & Benz, B. F.) 187–197 (Elsevier, New York, 2006).
Ericson, J.E., West, M., Sullivan, C., & Krueger, H. W. The development of maize agriculture in the Virú valley, Peru in The Chemistry of Prehistoric Human Bone, (ed. Price, T. D.) 68–104 (Cambridge University Press. New York, 1989).
Gerdau-Radonić, K., Goude, G., Castro de la Mata, P., André, G., Schutkowski, H., & Makowski, K. Diet in Peru's pre-Hispanic central coast. J. Arch. Sci. Rep. 4, 371–386 (2015).
Fernandes, R., Millard, A. R., Brabec, M., Nadeau, M. J. & Grootes, P. M. Food reconstruction using isotopic transferred signals (FRUITS): A Bayesian model for diet reconstruction. PLoS ONE 9(2), e87436 (2014).
Fernandes, R., Grootes, P. M., Nadeau, M. J. & Nehlich, O. Quantitative diet reconstruction of a neolithic population using a Bayesian mixing model (FRUITS): The case study of Ostorf (Germany). Am. J. Phys. Anthropol. 158, 325–340 (2015).
Colonese, A. C. et al. Stable isotope evidence for dietary diversification in the pre-Columbian Amazon. Sci. Rep. 10, 16560 (2020).
DeNiro, M. J. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317, 806–809 (1985).
Van Klinken, G. J. Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Arch. Sci. 26, 687–695 (1999).
Vázquez, V. F., Rosales Tham, T., Dillehay T. D., & Netherly, P. J. Faunal remains. In Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru (ed. Dillehay, T. D.) 197–366 (University of Texas, Austin, TX, 2017).
Elera, C. The Puémape site and the Cupisnique culture: a case study on the origins and development of complex society in the Central Andes, Peru (PhD. Dissertation. University of Calgary, Alberta, 1998).
Hyland, C., Millaire, J.-F. & Szpak, P. Migration and maize in the Virú Valley: Understanding life histories through multi-tissue carbon, nitrogen, sulfur, and strontium isotope analyses. Am. J. Phys. Anthropol. 176, 21–35 (2021).
Reitz, E. J. Resource Use through Time at Paloma Peru. Bull. Fla. Mus. Natl. Hist. 44, 65–80 (2003).
Coutts, K. H., Chu, A. & Krigbaum, J. Paleodiet in late preceramic Peru: Preliminary isotopic data from Bandurria. J. Island Coast. Archaeol. 6, 196–210 (2011).
Sotelo, C. (ed.) Cuaderno de Investigación del Archivo Tello nº9. Paracas WariKayan (Museo de Arqueología y Antropología de la Universidad Nacional Mayor de San Marcos, Lima, 2012).
Cadwallader, L. Investigating 1500 years of dietary change in the Lower Ica Valley, Peru, using an isotopic approach (PhD Dissertation, University of Cambridge, UK, 2013).
Finucane, B. C. Mummies, maize, and manure: Multi-tissue stable isotope analysis of late prehistoric human remains from the Ayacucho Valley, Peru. J. Arch. Sci. 34(12), 2115–2124 (2007).
Turner, B. L. et al. Diet and foodways across five Millennia in the Cusco Region of Peru. J. Arch. Sci. 98, 137–148 (2018).
Kellner, C. M., & Schoeninger, M. J. Dietary Correlates to the development of nasca social complexity (A.D. 1–750). Lat. Am. Antiq. 23, 490–508 (2012).
Ikehara, H. C., Paipay, J. F. & Shibata, K. Feasting with Zea Mays in the Middle and Late Formative North Coast of Peru. Lat. Am. Antiq. 24, 217–231 (2013).
Goldstein, D. J., Coleman, R. C., & Williams, P. R. You are what you drink: a sociocultural reconstruction of Pre-Hispanic fermented beverage use at Cerro Baúl, Moquegua, Peru in Drink, Power, and Society in the Andes (ed. Jennings, J. and Bowser, B. J.) 133–176 (University Press of Florida. 2009).
Gil, A. F. et al. Isotopic evidence on human bone for declining maize consumption during the little ice age in central western Argentina. J. Archaeol. Sci. 49, 213–227 (2014).
Antúnez de Mayolo, S. Ciencia agrícola en el Perú Pre-Colombino in Estudios de la Ciencia en el Perú (ed. Yepes, E.) (CONCYTEC, Lima, 1986).
Sandweiss, D. H., Shady, R., Moseley, M., Keefer, D. K. & Ortloff, C. R. Environmental change and economic development in coastal Peru between 5,800 and 3,600 years ago. Proc. Natl. Acad. Sci. USA 106, 1359–1363 (2009).
Ortloff, C. R. & Moseley, M. E. 2600–1800 BCE Caral: Environmental change at a Late Archaic Period Site on North Central Coast Peru Ñawpa Pacha. J. Andean Archaeol. 32, 189–206 (2012).
Guédron, S. et al. Holocene variations in Lake Titicaca water level and their implications for sociopolitical developments in the central Andes. Proc. Natl. Acad. Sci. USA 120, e2215882120 (2023).
Robinson, J. R. Investigating isotopic niche space: Using rKIN for stable isotope studies in archaeology. J. Archaeol. Method Theory 29, 831–861 (2022).
Koch, P. L., Tuross, N. & Fogel, M. L. The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxyapatite. J. Archaeol. Sci. 24, 417–429 (1997).
France, C. A. M., Sugiyama, N. & Aguayo, E. Establishing a preservation index for bone, dentin, and enamel bioapatite mineral using ATR-FTIR. J. Arch Sci. Rep. 33, 102551 (2020).
Tsutaya, T. & Yoneda, M. Reconstruction of breastfeeding and weaning practices using stable isotope and trace element analyses: A review. Am. J. Phys. Anthropol. 156, 2–21 (2015).
Marino, B. D. & McElroy, M. B. Isotopic composition of atmospheric CO2 inferred from carbon in C4 plant cellulose. Nature 349, 127–131 (1991).
DeNiro, M. J. & Hastorf, C. A, Alteration of 15N/14N and 13C/12C ratios of plant matter during the initial stages of diagenesis: Studies utilizing archaeological specimens from Peru. Geochim. Cosmochim. Acta 49, 97–115 (1985).
Szpak, P. & Chiou, K. L. A comparison of nitrogen isotope compositions of charred and desiccated botanical remains from northern Peru. Veg. Hist. Archaeobot. 29, 527–538 (2019).
Fernandes, R. A simple (R) model to predict the source of dietary carbon in individual consumers. Archaeometry 58, 500–512 (2016).
Fernandes, R., Nadeau, M. J. & Grootes, P. M. Macronutrient-based model for dietary carbon routing in bone collagen and bio-apatite. Arch. Anthropol. Sci. 4, 291–301 (2012).
Jim, S., Jones, V., Ambrose, S. H. & Evershed, R. P. Quantifying dietary macronutrient sources of carbon for bone collagen biosynthesis using natural abundance stable carbon isotope analysis. Br. J. Nutr. 95, 1055–1062 (2006).
Pestle, W. J., Crowley, B. E. & Weirauch, M. T. Quantifying inter-laboratory variability in stable isotope analysis of ancient skeletal remains. PLoS ONE 9(7), e102844 (2014).
Acknowledgements
The authors are grateful to Krista McGrath and Thiago Fossile for the review of the manuscript and their valuable comments on the initial draft.
Funding
This project has received funding from the European Union’s Horizon Europe research and innovation program under the Marie Sklodowska-Curie grant agreement No 101062179 (Paleodietary analyses of the first Andean cities: high-resolution assessment to macronutrients using a multiproxy approach—PACHAMAMA). This work was also funded by the ERC Consolidator project TRADITION, which has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program under Grant Agreement No 817911. This work contributes to the “ICTA-UAB María de Maeztu” Program for Units of Excellence of the Spanish Ministry of Science and Innovation (CEX2019-000940-M). This work also contributes to EarlyFoods (Evolution and impact of early food production systems), which has received funding from the Agència de Gestió d'Ajuts Universitaris i de Recerca de Catalunya (SGR-Cat-2021, 00527). The funders had no role in the study design, data collection and analysis, the decision to publish, or the preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
L.P.L. and A.C.C. designed research, L.P.L. performed research, L.P.L. and A.C.C. analyzed data and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pezo-Lanfranco, L., Colonese, A.C. The role of farming and fishing in the rise of social complexity in the Central Andes: a stable isotope perspective. Sci Rep 14, 4582 (2024). https://doi.org/10.1038/s41598-024-55436-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-55436-4
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
