
Abstract
Sugi (Cryptomeria japonica D. Don) is an economically important coniferous tree in Japan. However, abundant sugi pollen grains are dispersed and transported by the wind each spring and cause a severe pollen allergy syndrome (Japanese cedar pollinosis). The use of pollen-free sugi that cannot produce pollen has been thought as a countermeasure to Japanese cedar pollinosis. The sugi CjACOS5 gene is an ortholog of Arabidopsis ACOS5 and rice OsACOS12, which encode an acyl-CoA synthetase that is involved in the synthesis of sporopollenin in pollen walls. To generate pollen-free sugi, we mutated CjACOS5 using the CRISPR/Cas9 system. As a result of sugi transformation mediated by Agrobacterium tumefaciens harboring the CjACOS5-targeted CRISPR/Cas9 vector, 1 bp-deleted homo biallelic mutant lines were obtained. Chimeric mutant lines harboring both mutant and wild-type CjACOS5 genes were also generated. The homo biallelic mutant lines had no-pollen in male strobili, whereas chimeric mutant lines had male strobili with or without pollen grains. Our results suggest that CjACOS5 is essential for the production of pollen in sugi and that its disruption is useful for the generation of pollen-free sugi. In addition to conventional transgenic technology, genome editing technology, including CRISPR/Cas9, can confer new traits on sugi.
Similar content being viewed by others


Introduction
Sugi (Japanese cedar, Cryptomeria japonica D. Don) is a conifer species in the family Cupressaceae in gymnosperms1. It is one of the important domestic trees in Japan for forestry, industry, and the economy. Artificially sugi-planted areas had reached 4.44 million ha in 2017, covering about 12% of the land area of Japan2. The production of sugi roundwood was 12.28 million m3, which represented the largest volume (57% of the total volume) among the domestic tree species in 20173. Sugi wood is widely used as a structural, construction, and packaging material, as well as for flooring, ceiling boards, barrels, chopsticks, etc.
Conversely, sugi pollen allergy syndrome (Japanese cedar pollinosis) is a serious disease in Japan4. Sugi produces male strobili and female strobili, similar to other monoecious conifer trees. Sugi pollen is dispersed from male strobili in early spring and is carried far away by the wind. The pollen grain contains multiple allergen proteins: Cry J 1 (pectate lyase), Cry J 2 (polygalacturonase), Cry J 3 (thaumatin-like protein), CJP-4 (class IV chitinase), CJP-6 and CJP-8 (isoflavone reductase-like lipid transfer proteins), CPA9 (subtilisin-like serine protease), and CPA63 (aspartic protease)4. The pollen grains enter the human body and adhere to the nasal and ocular mucosa. They are burst by water absorption, release cytoplasmic components, including the allergens, and induce type II immunity. Japanese cedar pollinosis was first reported in 1964 in Nikko, Japan5. Thereafter, several nationwide surveys showed that the number of patients with Japanese cedar pollinosis had been increasing: the estimated prevalence was 11.7% in 19986, 13.1% in 20017, 26.5% in 20088, and 38.8% in 20199.
The reduction of the amount of sugi pollen has been thought as a countermeasure to Japanese cedar pollinosis. For this purpose, wild varieties with fewer male strobili and wild male-sterile mutants without pollen grains have been used for breeding. Male-sterile mutants were discovered in many plant species and the genetic patterns of male sterility are divided into genic male sterility (GMS) and cytoplasmic male sterility10,11. GMS is also called nuclear male sterility or Mendelian sterility because it depends exclusively on the nuclear genome and exhibits Mendelian inheritance. The development of genetic analyses has served to identify several key genes involved in GMS. For example, a transposon insertion mutant of the Arabidopsis thaliana acyl-CoA synthetase 5 (ACOS5) gene does not produce mature pollen in anthers12. The recombinant ACOS5 protein catalyzes oleic acid CoA-ester formation in vitro, and thus probably participates in the biosynthesis of sporopollenin, which is a constituent of the exine of pollen grains. Rice (Oryza sativa) carries the OsACOS12 gene, which is an ortholog of ACOS5, and the osacos12 mutant also shows a phenotype of absence of mature pollen13,14.
A male-sterile sugi without pollen was first identified in Japan in 199215,16; subsequently, various male-sterile mutant trees were discovered and analyzed17,18,19. To identify the genes that are responsible for male sterility in sugi, genetic marker analyses have been performed, leading to the identification of four recessive male sterility-linked independent loci (ms1, ms2, ms3 and ms4)20,21,22,23,24. Recently, one gene (MS1) at the ms1 locus was identified as a causative gene for male sterility25. MS1 encodes a lipid transfer protein and is expressed in male strobili, but its biochemical properties or physiological roles remain unknown. The appearance rate of pollen-free sugi trees is approximately 0.02% in a seed orchard in Japan26, and these trees are extremely rare in the field. In addition, the discovered mutant trees should be crossed to sugi plus trees with excellent properties for the generation of pollen-free plus trees, because most of the original male-sterile mutant trees exhibit slow growth or curved trunks. Therefore, the development of male-sterilization technology is valuable and needed for the generation of pollen-free sugi plus trees.
In this study, we aimed to generate pollen-free sugi trees using the CRISPR/Cas9 system. Genome editing technology including the CRISPR/Cas9 system has been widely utilized in plants and animals for the elucidation of gene function and for the breeding of new varieties. Because of the simplicity of its experimental procedures, the CRISPR/Cas9 system has been applied not only to herbaceous plants and crops, but also to woody plants such as sweet orange trees27, apple trees28, grape vines29, poplar trees30,31, sugi trees32, radiata pine33, and white spruce34. Although the use of genetically modified organisms (GMOs) including transgenic trees have been regulated by law in Japan, the products by the genome editing technology are less restricted by law, if they have no DNA from other species. Recently, γ-aminobutyric acid (GABA)-enriched tomato which had been generated by CRISPR/Cas9 started to be commercially sold without the regulation against GMOs35. The genome editing technology has the potential to be put the modified and improved forest trees to practical use in future. Here we report the genome editing of sugi by the CRISPR/Cas9 system, to mutate the CjACOS5 gene, which is an ortholog of Arabidopsis ACOS5 and rice OsACOS12. The regenerated sugi trees with mutated CjACOS5 genes flowered but did not produce pollen grains in male strobili. We demonstrated that the CRISPR/Cas9 system enables the generation of pollen-free sugi trees artificially, even though we had not been able to achieve this in the past.
Results
Isolation of the CjACOS5 genes from sugi
To induce male sterility in sugi, we searched for the target genes that are disrupted using the CRISPR/Cas9 system. We noticed that the A. thaliana acos5 mutant and the rice osacos12 mutant have been reported to exhibit male sterility and not to produce pollen12,13. Both the ACOS5 (TAIR ID: AT1g62940) and OsACOS12 (MSU ID: LOC_Os04g24530, RAP-DB ID: Os04g0310800) genes encode an acyl-CoA synthetase that is involved in sporopollenin synthesis during pollen formation. We searched for ACOS5-homologous genes in full-length cDNA libraries of sugi male strobili36, and identified one cDNA, CMFL003_A04 (DDBJ accession number: FX341539). The NCBI blastn program showed that CMFL003_A04 was homologous to Phoenix dactylifera 4-coumarate-CoA ligase-like 1 (GenBank accession: XM_008810877, e-value: 1e−81), Amborella trichopoda 4-coumarate-CoA ligase-like 1 (XM_006850938, 7e−55), A. thaliana ACOS5 (AY250836, 3e−53), Capsella rubella 4-coumarate-CoA ligase-like 1 (XM_006301215, 4e−52), etc.
Furthermore, we cloned cDNAs homologous to CMFL003_A04 from male strobili of sugi #13-8-2 line and sequenced. As a result, we identified two cDNAs, CjACOS5a (accession number: LC726337) and CjACOS5b (LC726338). The predicted protein encoded by CjACOS5a and CjACOS5b comprises 557 amino acid residues and their 551 residues (98.9%) are equal to each other (Fig. 1). The CjACOS5a protein has 61.9% identity to ACOS5 and 59.8% identity to OsACOS12. The CjACOS5 proteins are composed of two domains; one is an AMP-dependent synthetase/ligase domain (InterPro ID: IPR000873), from Glu41 to Tyr454, and the other domain is a C-terminal domain in AMP-binding enzymes (IPR025110), from Glu463 to Lys538 (Fig. 1). Both domains also exist in ACOS5 and OsACOS12. In the AMP-dependent synthetase/ligase domain, the CjACOS5 proteins have a conserved AMP-binding site (IPR020845), from Leu197 to Lys208 (LPYSSGTTGASK) (Fig. 1). This conserved AMP-binding site also appears in long-chain fatty acid-CoA ligases and 4-coumarate-CoA ligases, according to the InterPro web site. OsACOS12 also carries the conserved AMP-binding site (197-LPYSSGTTGVSK-208), whereas ACOS5 carries a similar amino acid sequence (186-LPFSSGTTGLQK-197), which was not determined to be IPR020845. As the result of these genetic analyses, CjACOS5a and CjACOS5b were thought to be sugi orthologs of ACOS5 and OsACOS12 and encode an acyl-CoA synthetase. The expression of the CjACOS5a gene in male strobili was about 280-fold higher than that detected in stems, whereas that in the leaves and female strobili was not significantly different from that in stems (Supplementary Fig. S1). Similarly, CjACOS5b was expressed most highly in male strobili, and their mRNA level was about 150-fold higher than that in stems. Accordingly, we expected that the CjACOS5 genes mainly worked in sugi male strobili and was involved in pollen formation, similar to Arabidopsis ACOS5 and rice OsACOS12.

Comparison of the amino acid sequences of ACOS5 orthologs. CjACOS5a and CjACOS5b from sugi, ACOS5 from Arabidopsis thaliana, and OsACOS12 from rice were aligned using by the MAFFT program82. Identical amino acids to CjACOS5a are indicated in blue. The AMP-dependent synthetase/ligase domain (IPR000873), C-terminal domain in AMP-binding enzymes (IPR025110), and conserved AMP-binding site (IPR020845) are shown in green, pink, and red colors, respectively.
CRISPR/Cas9 vector construction and sugi transformation
To induce loss of function of CjACOS5 in sugi, we planned the target sites for DNA breaks by the CRISPR/Cas9 system. The recruitment of the complex of guide RNA and Streptococcus pyogenes Cas9 requires 5′-NGG-3′ as a protospacer adjacent motif (PAM) and the upstream target sequence. The AGG sequence located downstream of the start codon of CjACOS5 was choose as a PAM (Fig. 2a). If the CjACOS5 gene is broken at the predicted site, at 3 base pairs upstream of the PAM, short CjACOS5 proteins with about 34 or 24 amino acid residues would probably be synthesized by a flame shift. Two allelic CjACOS5 genes, CjACOS5a and CjACOS5b, were distinguished by a single nucleotide polymorphism (T or G) located upstream of the start codon (Fig. 2a).

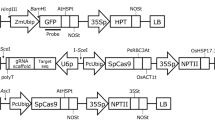
Design of the binary vector for the CRISPR/Cas9 system. (a) CjACOS5 cDNA sequence neighboring the DNA break site for the CRISPR/Cas9 system. The target sequence and PAM are shown in red and underlined blue, respectively. The arrow head is located at a predicted DNA break site. The reverse-typed K is a single nucleotide polymorphism and indicates thymine in the CjACOS5a gene or guanine in the CjACOS5b gene. Amino acid residues are indicated under the cDNA sequence. (b) Schematic representation of the constructed CRISPR/Cas9 binary vector. This vector was named pBFGE1 and was derived from pZK_gYSA_FFCas937. Twenty base pairs upstream of the PAM of CjACOS5 were inserted as the target sequence. AtU6, Arabidopsis thaliana U6 promoter; gRNA scaffold, guide RNA; PcUbi, Petroselinum crispum ubiquitin promoter; At-optimized Cas9, A. thaliana-optimized Streptococcus pyogenes Cas9; NLS, simian virus 40 nuclear localization signal; Tpea3A, Pisum sativum rbcS3A terminator; Tact, Oryza sativa actin terminator; P35S, cauliflower mosaic virus 35S promoter; ZmUbi, Zea mays ubiquitin promoter, NPTII, neomycin phosphotransferase II; Thsp, O. sativa heat shock protein terminator; RB, right border; LB, left border.
A CRISPR/Cas9 vector was constructed for transformation of sugi. The original pZK_gYSA_FFCas9 vector was generated for dicots and is composed of the A. thaliana U6 promoter-driven guide RNA, the parsley ubiquitin promoter-driven Cas9, and the cauliflower mosaic virus (CaMV) 35S promoter-driven NPTII gene37. We modified pZK_gYSA_FFCas9 to generate the pBFGE1 vector, in which the Zea mays ubiquitin promoter was inserted between the CaMV 35S promoter and the NPTII gene, to further express the NPTII gene (Fig. 2b). The ubiquitin promoter has been reported to work well in sugi38. The 20-bp target sequence of CjACOS5 was integrated into the guide RNA gene of the pBFGE1 vector, to break the CjACOS5 genes (Fig. 2b).
The sugi embryogenic cell line #13-8-2 was infected with Agrobacterium tumefaciens GV3101 harboring the constructed pBFGE1 vector to generate transgenic sugi trees. After infection, the cells were cultivated in selection medium containing kanamycin and formed calli harboring the vector. We picked six proliferated calli and investigated whether they had the NPTII gene by PCR. Among the six calli, three kanamycin-resistant calli lines were obtained and designated as GE#1, GE#2 and GE#3 (Supplementary Fig. S2a). These kanamycin-resistant calli and the non-transgenic #13-8-2 calli were transferred to kanamycin-free maturation medium, and then all of the calli formed somatic embryos in about 2 months. Subsequently, the somatic embryos were moved onto germination media, germinated, and grown. The regenerated transgenic plantlets showed normal shape with needles and roots as same as regenerated non-transgenic plantlets. The rooted transgenic sugi plantlets were planted in a pot and grown in a closed glass greenhouse (phytotron). All sugi plantlets regenerated from the GE#1, GE#2 and GE#3 calli possessed the NPTII gene in their leaves, whereas non-transgenic sugi plantlets obtained from the #13-8-2 cells did not (Supplementary Fig. S2b).
Induced mutation of CjACOS5
To confirm the mutation of CjACOS5 in the transgenic trees regenerated from the GE#1, GE#2 and GE#3 calli, genomic DNA was isolated from the leaves of each transgenic sugi plantlet, then DNA fragments including the target site of CjACOS5 were amplified using PCR and cloned into a plasmid vector in E. coli. The plasmids were individually sequenced and analyzed. As a result, several deletion or insertion mutants were detected in the target DNA sequence of CjACOS5a and CjACOS5b in the transgenic sugi trees, whereas non-transgenic sugi trees that were regenerated from embryogenic cells had no mutation in the two genes (Supplementary Table S2).
The two GE#1 callus-derived transgenic sugi lines, GE#1-17 and GE#1-18, were chimeric plants (Fig. 3). The GE#1-17 sugi tree carried a 3-bp deletion and a 1-bp deletion of CjACOS5a and two types of 1-bp deletion of CjACOS5b. GE#1-18 contained wild-type and 1 bp-deleted CjACOS5a and a 1-bp deletion of CjACOS5b. In contrast, all eight transgenic sugi lines derived from GE#2 calli (GE#2-1 to GE#2-22) had the same biallelic mutation with a 1-bp deletion at the same position of CjACOS5a and CjACOS5b (Fig. 3 and Supplementary Table S2). These deletions of a single base pair (G:C) suggested the production of a truncated and mutated CjACOS5 protein of 34 amino acid residues (MADQNAVREKRRSFSAAPFQLFWFLRTLVYRNSF). In this mutant CjACOS5 protein, the nine underlined-N-terminal amino acid residues alone are in accordance with the original CjACOS5 protein. Conversely, GE#3-derived transgenic trees include several types of mutation. Five transgenic sugi lines (GE#3-7, GE#3-8, GE#3-9, GE#3-11, and GE#3-12) were chimeric plants including wild-type and deleted CjACOS5 genes (Fig. 3 and Supplementary Table S2). There were not only 1-bp deletions in those plants, but also a 45-bp deletion in GE#3-9, and a new DNA sequence comprising a 17-bp deletion and a 24-bp insertion in GE#3-12. GE#3-10 alone did not contain the wild-type CjACOS5a and CjACOS5b genes; thus, it was a biallelic mutant, similar to the GE#2 lines (Fig. 3).

Mutations of CjACOS5 genes by CRISPR/Cas9. DNA sequence of PCR-amplified DNAs neighboring the break site of CjACOS5 in a non-transgenic sugi tree (WT) and in transgenic tree (GE#1, GE#2, and GE#3) lines. The asterisk shows a single nucleotide polymorphism in CjACOS5a and CjACOS5b. The target sequence and PAM are shown in reversed black and underlined blue, respectively. The gaps (−) represent deleted nucleotides. Reads mean the number of sequenced DNAs after cloning of the PCR products amplified using the leaf DNA.
We investigated off-target mutagenesis of sugi genes which had similar sequences to the CjACOS5 target sequence and the PAM. Because the sugi genome is huge and has never been clearly elucidated yet, three genes, CJt056454, CJt031386, and CJt113843, were chosen from the sugi transcriptome assembly (CJ3006NRE)39 using the blastn analysis. In conclusion, the PCR products of the three genes from the investigated transgenic sugi lines, GE#1-17, GE#1-18, GE#2-1, and GE#3-7 had only normal sequences and showed no off-target mutagenesis (Supplementary Fig. S7). These results confirmed that the CRISPR/Cas9 system worked in sugi cells and induced mutations of the CjACOS5a and CjACOS5b genes.
Morphogenesis and pollen production in CjACOS5-mutataed sugi trees
The transgenic sugi trees were planted in a pot and grown under the same conditions of outside temperature and sunshine in the phytotron (Supplementary Fig. S3a). The shape of the transgenic sugi trees from GE#2 lines and GE#3 lines appeared to be the same as that of non-transgenic sugi trees (Supplementary Fig. S3b, e–i). The initial height of trees from GE#2 lines and GE#3 lines was similar to that of non-transgenic sugi trees at 6 months, whereas GE#1-17 and GE#1-18 trees were significantly shorter than non-transgenic trees (Supplementary Fig. S3c, d, j). Unfortunately, GE#1-17 and GE#1-18 trees withered and died in the winter of 2017, probably because of severe growth suppression.
Flowering of sugi can be induced by spraying a gibberellin A3 (GA3) solution onto the twigs with leaves40. At the end of July/beginning of August of 2017, the young transgenic sugi trees with the mutation of CjACOS5 and non-transgenic sugi trees were treated twice with GA3 and cultivated in the phytotron. Visible flower buds appeared in October and developed to male strobili. In January of 2018, yellow-powdered pollen was packed in the male strobili of non-transgenic sugi trees (Fig. 4a). Spherical pollen grains in the male strobili were detected under a microscope.

Male strobili of genome-edited sugi trees in 2018. (a) Non-transgenic sugi (WT) male strobili (left; scale bar, 1 cm), magnified male strobili (middle; scale bar, 1 mm), and razor-cutting suspension of male strobili (right; scale bar, 100 μm). The magnified strobili are shown as an outside (top) and a vertical section (bottom). Spherical pollen grains and the suspension were stained with Calberla’s fuchsin staining solution (right). (b) Male strobili of the genome-edited GE#2-1 sugi tree, a biallelic mutant of CjACOS5. (c) GE#2-2, a biallelic mutant. (d) GE#3-7, a chimeric transgenic sugi tree. Normal male strobili and no-pollen male strobili are indicated. (e) GE#3-8, a chimeric transgenic sugi tree with pollen. (f) GE#3-10, a biallelic mutant without pollen. The photographs were taken in January, 2018.
In contrast, GE#2 sugi trees from GE#2-1 to GE#2-6, which were biallelic mutants of CjACOS5a and CjACOS5b, produced male strobili without pollen grains (Fig. 4b, c, and Supplementary Table S2). The male strobili were finely sliced using a razor blade in water, and the suspension was stained and observed under a microscope, however, no-pollen grains were detected and small cells were observed. Conversely, GE#3 sugi trees resulted in three types, as follows. The chimeric trees with wild-type and deleted genes, GE#3-7, GE#3-9, GE#3-11, and GE#3-12, had pollen-including male strobili and no-pollen male strobili (Fig. 4d and Supplementary Table S2). GE#3-8 was also a chimeric tree but had male strobili with pollen (Fig. 4e). GE#3-10, the sole biallelic mutant among the GE#3 lines, had no-pollen at all, similar to the GE#2 lines (Fig. 4f). Therefore, disruption of both CjACOS5a and CjACOS5b led to the loss of the ability of pollen production in male strobili. Moreover, the pollen productivity was shown to be maintained exclusively by one allele gene, CjACOS5a or CjACOS5b.
A sterile male strobilus of GE#2-1 and GE#2-2 tree carried only deleted CjACOS5 genes as same as their leaves (Supplementary Fig. S4). Conversely, a sterile male strobilus of GE#3-7 carried not only deleted CjACOS5 genes but also wild-type genes similar to a fertile male strobilus of GE#3-7. That is probably because a male strobilus had been constructed with not only mutated reproductive cells but also somatic cells without mutation. Additionally, the male strobilus of GE#3-7 possessed different types of mutations, a 2-bp deletion and a 4-bp deletion, which were not been observed in the leaves (Supplementary Fig. S4 and Fig. 3). The plausible reason is that GE#3-7 is a chimeric tree carrying the active Cas9 gene which has been inducing the mutation of the CjACOS5 genes.
To determine the reproducibility of the no-pollen flowers, the mutated sugi trees were again treated with GA3 in the summer of 2018. They were grown in the phytotron and in a special netted-house. Biallelic mutant sugi trees, such as GE#2-1 and GE#3-10, grown in the phytotron had male strobili but did not produce pollen in 2019 (Supplementary Fig. S5b, c, f). The chimeric mutants (GE#3-7, GE#3-9, GE#3-11, and GE#3-12) produced two types of strobili, i.e., with or without pollen (Supplementary Fig. S5d). GE#3-8 produced only fertile male strobili similar to the result of the preceding year (Supplementary Fig. S5e). The special netted-house is bigger than the phytotron and has a glass roof and a metal mesh for ventilation. That did not have an air conditioner, so that its temperature depended on the outside temperature and sunlight but often reached around 40 °C in the summer. The sugi trees suffered no damage under high temperature conditions. In the special netted-house, the biallelic mutant trees (GE#2-21 and GE#2-22) did not produce pollen in their male strobili, whereas the non-transgenic trees had pollen (Supplementary Fig. S5g–i). During the third experimental period, from 2019 to 2020, CjACOS5 biallelic mutant GE#2 lines also had male strobili but did not produce pollen in the special netted-house (Supplementary Fig. S6). Accordingly, it was suggested that CRISPR/Cas9-induced mutation of CjACOS5 genes stably maintained the no-pollen phenotype over a few years in sugi trees.
Discussion
Pollen-free sugi trees have been expected to mitigate Japanese cedar pollinosis. For that reason, male-sterile pollen-free sugi trees have been the goal of time-consuming and laborious research, and are rarely found in the field or in the test forests. In the present study, we accomplished the artificial generation of pollen-free sugi using genome editing technology. Sugi CjACOS5 genes were targeted and mutated by a CRISPR/Cas9 vector via Agrobacterium-mediated transformation. The regenerated sugi trees with biallelic CjACOS5 deletions could not produce pollen in their male strobili (Fig. 4). Although small dots were observed in the suspension of no-pollen male strobili (Fig. 4), we were not sure that they were pieces of cell debris or developing microspores which were progenitor cells of pollen grains. Even if the small dots were microspores, they would be expected to be finally degraded and not to be dispersed from male strobili, because there are many reports that male sterile mutants do not release microspores outside. In the Arabidopsis acos5 mutant, microspores are lysed and completely degraded on the way of development12. Similarly, the rice osacos12 mutant degenerates microspores and does not have mature pollen at later stage of anther development13. Sugi male sterile 1 (ms1) mutants neither produce nor release pollen, because microspores are not separated in the tetrad stage and then are degenerated16,18. In other sugi male sterile mutants, ms2 and ms3, micropores adhere each other and collapse in anthers, and thus the microspores are not dispersed outside17. Accordingly, biallelic CjACOS5-mutated sugi trees are thought not to release small developing microspores outside male strobili.
Male sterility with no-pollen has been regarded as being important and useful for human health, the economy, biodiversity, and agriculture. For example, if plant species with a large amount of allergenic pollen, such as sugi, cypress, and birch trees and pasture grasses, cannot produce pollen, the number of pollen-allergic patients will decrease. As the result, the pollinosis-related health expenditures and economic loss will decrease41. Pollen-free plants can also avoid unnecessary gene flow, which causes problems of biodiversity in the case of invasive and GMO plants42. Male sterility in a variety of crops (maize, rice, wheat, etc.) has helped produce hybrid seeds more efficiently than before43,44,45. Using genome editing technology, it is possible to turn a plant species with pollen into pollen-free varieties, even if wild pollen-free mutants had never been found in the same plant species. Thus, the generation of male-sterile plants using the CRISPR/Cas9 system is innovative and greatly available.
The no-pollen phenotype in male strobili of the genome-edited sugi trees was thought to be caused by the mutation of CjACOS5a and CjACOS5b, as demonstrated by the results obtained for different mutation types. The GE#2 lines and the GE#3-10 line had biallelic mutation of both deleted CjACOS5a and CjACOS5b, and they showed a no-pollen phenotype in male strobili (Figs. 3 and 4). These results demonstrated that biallelic mutation of CjACOS5a and CjACOS5b was closely related to the no-pollen phenotype. GE#3-7 produced male strobili partly with pollen or without pollen. Because GE#3-7 was a chimeric tree carrying the deleted CjACOS5a gene and the normal and delated CjACOS5b gene, it was suggested that normal CjACOS5b functioned in pollen production. Male strobili without pollen and with pollen were thought to be derived from biallelic mutant cells and monoallelic mutant cells, respectively. Conversely, it was thought that CjACOS5a alone could lead to the production of pollen, because GE#3-8 had normal CjACOS5a, deleted CjACOS5b, and normal CjACOS5b and produced only male strobili with pollen. Therefore, it was concluded that the no-pollen phenotype observed in this study was caused by loss of function of both the CjACOS5a and CjACOS5b genes, and that one allelic gene, alone, either CjACOS5a or CjACOS5b, was essential to generate pollen in male strobili. If possible, to confirm that the no-pollen phenotype was not caused by off-target mutagenesis, we should elucidate the existence of off-target mutagenesis in the genome-edited sugi trees in the same way as that used for several model plants46. However, the off-target mutagenesis in sugi is difficult to investigate because its genomic DNA is very large (about 11 Gb)47 and has never been adequately read. Here, we investigated the off-target mutagenesis of only three genes similar to the CjACOS5 target sequence and verified they had no mutation (Supplementary Fig. S7). For the development of genome editing technology in sugi, more extensive survey of off-target mutagenesis is surely needed in the future.
We do not know the reason why GE#1-17 and GE#1-18 showed severe growth retardation. Both transgenic lines had no off-target mutagenesis in the investigated three genes (Supplementary Fig. S7), though other mutations might occur in important genes by CRISPR/Cas9. On the other hand, we cannot deny the possibility of somaclonal mutation in GE#1-17 and GE#1-18. Few studies have been reported on the somaclonal mutation of sugi during cell culture and somatic embryogenesis, however, there are reports of somaclonal mutation or abnormal growth in other gymnosperms such as Picea48,49,50, Pinus51, and Larix52,53. Therefore, severe growth inhibition and lethality in GE#1-17 and GE#1-18 may be attributed to somaclonal mutation.
The sugi CjACOS5 gene was revealed to be essential for pollen production in this study. The Arabidopsis ACOS5 protein is an acyl-CoA synthetase that is estimated to be linked to the biosynthesis of sporopollenin in the exine of pollen12. The rice OsACOS12 protein is the ortholog of ACOS5 and catalyzes the condensation of oleic acid and CoA in vitro14. Although the physiological and biochemical functions of CjACOS5 were not clarified in this study, CjACOS5 was deduced to function as an acyl-CoA synthetase based on the structural analogy to ACOS5 and OsACOS12 (Fig. 1). Sporopollenin biosynthesis requires not only acyl-CoA synthetase, but also several other enzymes encoded by the MALE STERILITY 2 (MS2)54,55, CYP703A256, CYP704B157, LESS ADHESIVE POLLEN 3 (LAP3)58, LAP5/POLYKETIDE SYNTHASE B (PKSB)59,60, LAP6/PKSA59,60, and DIHYDROFLAVONOL 4-REDUCTASE-LIKE 1 (DRL1)/TETRAKETIDE α-PYRONE REDUCTASE 1 (TKPR1) genes61,62. The Arabidopsis ms2 mutant, the drl1/tkpr1 mutant, the lap3 mutant, and the double mutant of lap5 and lap6 (pksb and pksa) show complete male sterility. Similarly, the rice defective pollen wall (dpw, the ortholog of MS2)63, Ospks164, Ospks265, and Ostkpr166 mutants exhibit a male-sterile phenotype individually. The wheat TaNP1 gene (encoding a putative glucose-methanol-choline oxidoreductase) is an ortholog of OsNP1 that is involved in tapetum degeneration and pollen exine formation67. As the result of TaNP1 disruption by CRISPR/Cas9, triple homozygous mutants of TaNP1 showed complete male sterility. The tomato SlSTR1 gene (a stamen-specific expressed strictosidine synthase) is an ortholog of Arabidopsis LAP3, and CRISPR/Cas9-mediated mutagenesis of SlSTR1 confers no viability on pollen68. Those male sterility-related gene orthologs of sugi probably can be utilized in addition to CjACOS5 to generate new varieties with male sterility by CRISPR/Cas9.
Our study established that the genome editing technology is available as a method for generating pollen-free sugi trees. So far, conventional crossbreeding programs using wild male-sterile sugi mutants have been useful for the generation of new pollen-free varieties of sugi. Because these crossed varieties can be freely handled, unlike the case of GMO trees, the new pollen-free sugi varieties that are the progeny of sugi plus trees and wild pollen-free mutants have been gradually planted in Japan3. In contrast, the CjACOS5-disrupted pollen-free sugi trees obtained in this study are GMO trees and, thus, are currently restricted for handling and planting by law. However, the genome editing technology, including CRISPR/Cas9, has several advantages compared with conventional tree breeding, as follows. First, if we could identify target genes involved in excellent traits, genome editing technology would be able to directly modify the target genes and generate their mutant trees in cases in which mutants of the target genes are not found in nature. Second, CRISPR/Cas9 can edit several target genes simultaneously and, thus, can modify multiple traits. Considering the combination of expected traits, such as no-pollen, fast growth, quality of wood, straightness, and stress tolerance, multiple values can be conferred to sugi breeding using multiplex gene editing technology. Last, the genome editing technology can not affect every gene other than the target genes, if off-target mutagenesis can be controlled fully. Therefore, the genome editing technology is difficult to perturb the genetic composition of well-established varieties unlike crossbreeding. In case of using genome-edited trees in forestry, transgene-free null segregants are actually needed. To generate null segregants, we have been crossing the CjACOS5-mutated sugi trees with non-transgenic sugi trees. By genetic recombination and segregation, in future, we expect to obtain null segregant sugi progenies without the transgenes such as NPTII and Cas9. Recently, several methods have been developed for the generation of non-transgenic genome-edited plants69,70,71. Consequently, it is expected that they will be utilized in combination with the advantage of genome editing technology and conventional breeding methods for tree breeding.
Methods
Vector construction
pZK_gYSA_FFCas9 vector37 was digested with XbaI, the recognition site of which was located between the cauliflower mosaic virus 35S promoter and the NPTII gene, and blunted with Klenow fragment. About 2 kb of Zea mays ubiquitin promoter was inserted into the blunted pZK_gYSA_FFCas9 vector72. The resulting binary vector was designated as pBFGE1. Two synthetic DNAs (Supplementary Table S1) were annealed and ligated into the BbsI-digested pUC19_AtU6oligo vector73. A derivative vector was digested with I-SceI, and DNA fragments including the oligonucleotides were excised. The DNA fragments were inserted into I-SceI digested pBFGE1.
Plant materials and sugi transformation
The sugi (Cryptomeria japonica D. Don) embryonic cell line #13-8-2 was induced from an immature seed using previously described methods74. The #13-8-2 cells were transformed using A. tumefaciens GV3101 harboring the constructed pBFGE1 vector as described previously75,76. Selected kanamycin-resistant calli were regenerated through somatic embryogenesis77. Regenerated sugi plantlets were acclimatized in moist peat moss and Kanuma pumice, then transferred to moist vermiculite in a pot. They were cultivated under natural-day-length conditions in a closed glass greenhouse (phytotron) or a special netted-house. The temperature inside the phytotron was controlled by an air conditioner following the outdoor temperature. The special netted-house was a greenhouse with metal mesh windows that passed the outside air but impeded invasion by insects.
DNA analysis
Orthologs of the CMFL003_A04 cDNA were searched using the blastn program in the latest NCBI nucleotide database78,79. The CjACOS5 cDNAs were cloned as described in Supplementary Information. A functional domain analysis of ACOS5-related protein sequences was performed on the InterPro website80. Genomic DNA was isolated from embryogenic cells and from leaves using the DNeasy Plant Mini Kit (Qiagen). The NPTII and CjACOS5 genes were confirmed using PCR (Supplementary Information). To elucidate the mutation induced by genome editing, a DNA region including the predicted DNA break site for the CRISPR/Cas9 system was amplified using PCR (Supplementary Information). The amplified DNA fragments were cloned into the pBlueScript II vector and sequenced.
Induction and observation of male strobili
To induce flowering, 100 mg/L of a gibberellin A3 (GA3) aqueous solution was sprayed on sugi plantlets. In the first experiment, GA3 treatments were performed twice, on July 24 and August 3, 2017. Male strobili were observed in January, 2018. Pollen and the suspension of sliced male strobili were stained with Calberla’s fuchsin staining solution, for microscopy81. In the second and third experiments, GA3 treatments were performed in late July, 2018 and 2019, respectively. Thereafter, male strobili and pollen were analyzed in the following year in the same way as that described for first experiment.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Christenhusz, M. J. M. et al. A new classification and linear sequence of extant gymnosperms. Phytotaxa 19, 55–70 (2011).
Forestry Agency. Statistical Handbook of Forest and Forestry. (Japan Forest Foundation, 2017) (in Japanese).
Forestry Agency. Annual Report on Forest and Forestry in Japan. Fiscal Year 2018 (Summary). (Ministry of Agriculture, Forestry, and Fisheries, 2019).
Fujimura, T. & Kawamoto, S. Spectrum of allergens for Japanese cedar pollinosis and impact of component-resolved diagnosis on allergen-specific immunotherapy. Allergol. Int. 64, 312–320 (2015).
Horiguchi, S. & Saito, Y. Japanese cedar pollinosis in Nikko, Japan. Arerugi 13, 16–18 (1964) (in Japanese with English abstract).
Nakamura, A., Asai, T., Yoshida, K., Baba, K. & Nakae, K. Allergic rhinitis epidemiology in Japan. J. Otolaryngol. Jpn. 105, 215–224 (2002) (in Japanese with English abstract).
Okuda, M. Epidemiology of Japanese cedar pollinosis throughout Japan. Ann. Allergy Asthma Immunol. 91, 288–296 (2003).
Baba, K. & Nakae, K. Epidemiology of nasal allergy through Japan in 2008. Prog. Med. 28, 2001–2012 (2008) (in Japanese).
Matsubara, A. et al. Epidemiological survey of allergic rhinitis in Japan 2019. J. Otolaryngol. Jpn. 123, 485–490 (2020) (in Japanese with English abstract).
Vedel, F. et al. Molecular basis of nuclear and cytoplasmic male sterility in higher plants. Plant Physiol. Biochem. 32, 601–618 (1994).
Horner, H. & Palmer, R. Mechanisms of genic male sterility. Crop Sci. 35, 1527–1535 (1995).
de Azevedo Souza, C. et al. A novel fatty acyl-CoA synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis. Plant Cell 21, 507–525 (2009).
Li, Y. et al. OsACOS12, an orthologue of Arabidopsis acyl-CoA synthetase5, plays an important role in pollen exine formation and anther development in rice. BMC Plant Biol. 16, 256 (2016).
Yang, X. et al. Rice fatty acyl-CoA synthetase OsACOS12 is required for tapetum programmed cell death and male fertility. Planta 246, 105–122 (2017).
Taira, H., Teranishi, H. & Kenda, Y. A case study of male sterility in sugi (Cryptomeria japonica). J. Jpn. For. Soc. 75, 377–379 (1993) (in Japanese with English abstract).
Saito, M., Taira, H. & Furuta, Y. Cytological and genetical studies on male sterility in Cryptomeria japonica D. Don. J. For. Res. 3, 167–173 (1998).
Yoshii, E. & Taira, H. Cytological and genetical studies on male sterile sugi (Cryptomeria japonica D. Don), Shindai 1 and Shindai 5. J. Jpn. For. Soc. 89, 26–30 (2007) (in Japanese with English abstract).
Ueuma, H., Yoshii, E., Hosoo, Y. & Taira, H. Cytological study of a male-sterile Cryptomeria japonica that does not release microspores from tetrads. J. For. Res. 14, 123–126 (2009).
Miyajima, D., Yoshii, E., Hosoo, Y. & Taira, H. Cytological and genetic studies on male sterility in Cryptomeria japonica D. Don (Shindai 8). J. Jpn. For. Soc. 92, 106–109 (2010) (in Japanese with English abstract).
Moriguchi, Y. et al. The construction of a high-density linkage map for identifying SNP markers that are tightly linked to a nuclear-recessive major gene for male sterility in Cryptomeria japonica D. Don. BMC Genomics 13, 95 (2012).
Moriguchi, Y. et al. Establishment of a microsatellite panel covering the sugi (Cryptomeria japonica) genome, and its application for localization of a male-sterile gene (ms-2). Mol. Breed. 33, 315–325 (2014).
Moriguchi, Y. et al. A high-density linkage map with 2560 markers and its application for the localization of the male-sterile genes ms3 and ms4 in Cryptomeria japonica D. Don. Tree Genet. Genomes 12, 57 (2016).
Mishima, K. et al. Identification of novel putative causative genes and genetic marker for male sterility in Japanese cedar (Cryptomeria japonica D. Don). BMC Genomics 19, 277 (2018).
Hasegawa, Y. et al. Fine mapping of the male-sterile genes (MS1, MS2, MS3, and MS4) and development of SNP markers for marker-assisted selection in Japanese cedar (Cryptomeria japonica D. Don). PLoS ONE 13, e0206695 (2018).
Hasegawa, Y. et al. Identification and genetic diversity analysis of a male-sterile gene (MS1) in Japanese cedar (Cryptomeria japonica D. Don). Sci. Rep. 11, 1496 (2021).
Saito, M., Koga, Y., Furuta, Y. & Taira, H. Selections of male sterile sugi (Cryptomeria japonica D. Don) trees from open pollinated seedlings in a seed orchard. J. Jpn. For. Soc. 87, 1–7 (2005) (in Japanese with English abstract).
Jia, H. & Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 9, e93806 (2014).
Nishitani, C. et al. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 6, 31481 (2016).
Ren, C. et al. CRISPR/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 6, 32289 (2016).
Fan, D. et al. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 5, 12217 (2015).
Zhou, X., Jacobs, T. B., Xue, L. J., Harding, S. A. & Tsai, C. J. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate:CoA ligase specificity and redundancy. New Phytol. 208, 298–301 (2015).
Nanasato, Y. et al. CRISPR/Cas9-mediated targeted mutagenesis in Japanese cedar (Cryptomeria japonica D. Don). Sci. Rep. 11, 16186 (2021).
Poovaiah, C. et al. Genome editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 21, 363 (2021).
Cui, Y. et al. Efficient multi-sites genome editing and plant regeneration via somatic embryogenesis in Picea glauca. Front. Plant Sci. 12, 751891 (2021).
Nagamine, A. & Ezura, H. Genome editing for improving crop nutrition. Front. Genome Ed. 4, 850104 (2022).
Futamura, N. et al. Characterization of expressed sequence tags from a full-length enriched cDNA library of Cryptomeria japonica male strobili. BMC Genomics 9, 383 (2008).
Mikami, M., Toki, S. & Endo, M. Comparison of CRISPR/Cas9 expression constructs for efficient targeted mutagenesis in rice. Plant Mol. Biol. 88, 561–572 (2015).
Taniguchi, T., Ohmiya, Y., Kurita, M., Tsubomura, M. & Kondo, T. Regeneration of transgenic Cryptomeria japonica D. Don after Agrobacterium tumefaciens-mediated transformation of embryogenic tissue. Plant Cell Rep. 27, 1461–1466 (2008).
Wei, F. J. et al. Construction of a reference transcriptome for the analysis of male sterility in sugi (Cryptomeria japonica D. Don) focusing on MALE STERILITY 1 (MS1). PLoS ONE 16, e0247180 (2021).
Nagao, A., Sasaki, S. & Pharis, R. P. Cryptomeria japonica. in CRC Handbook of Flowering (ed. Halevy, A. H.) Vol IV. 247–269 (CRC Press, 1989).
Blaiss, M. S. Allergic rhinitis: direct and indirect costs. Allergy. Asthma. Proc. 31, 375–380 (2010).
Fritsche, S., Klocko, A. L., Boron, A., Brunner, A. M. & Thorlby, G. Strategies for engineering reproductive sterility in plantation forests. Front. Plant Sci. 9, 1671 (2018).
Wan, X. et al. Maize genic male-sterility genes and their applications in hybrid breeding: progress and perspectives. Mol. Plant 12, 321–342 (2019).
Abbas, A. et al. Exploiting genic male sterility in rice: from molecular dissection to breeding applications. Front. Plant Sci. 12, 629314 (2021).
Singh, M., Albertsen, M. C. & Cigan, A. M. Male fertility genes in bread wheat (Triticum aestivum L.) and their utilization for hybrid seed production. Int. J. Mol. Sci. 22, 8157 (2021).
Graham, N. et al. Plant genome editing and the relevance of off-target changes. Plant Physiol. 183, 1453–1471 (2020).
Hizume, M., Kondo, T., Shibata, F. & Ishizuka, R. Flow cytometric determination of genome size in the Taxodiaceae, Cupressaceae sensu stricto and Sciadopityaceae. Cytologia 66, 307–311 (2001).
Isabel, N. et al. Occurrence of somaclonal variation among somatic embryo-derived white spruces (Picea glauca, Pinaceae). Am. J. Bot. 83, 1121–1130 (1996).
Fourré, J. L., Berger, P., Niquet, L. & André, P. Somatic embryogenesis and somaclonal variation in Norway spruce: morphogenetic, cytogenetic and molecular approaches. Theor. Appl. Genet. 94, 159–169 (1997).
Tremblay, L., Levasseur, C. & Tremblay, F. M. Frequency of somaclonal variation in plants of black spruce (Picea mariana, Pinaceae) and white spruce (P. glauca, Pinaceae) derived from somatic embryogenesis and identification of some factors involved in genetic instability. Am. J. Bot. 86, 1373–1381 (1999).
Cuesta, C., Ordás, R. J., Rodríguez, A. & Fernández, B. PCR-based molecular markers for assessment of somaclonal variation in Pinus pinea clones micropropagated in vitro. Biol. Plant. 54, 435–442 (2010).
Krutovsky, K. et al. Somaclonal variation of haploid in vitro tissue culture obtained from Siberian larch (Larix sibirica Ledeb.) megagametophytes for whole genome de novo sequencing. In Vitro Cell. Dev. Biol. Plant 50, 655–664 (2014).
Goryachkina, O. V., Park, M. E., Tretyakova, I. N., Badaeva, E. D. & Muratova, E. N. Cytogenetic stability of young and long-term embryogenic cultures of Larix sibirica. Cytologia 83, 323–329 (2018).
Aarts, M. G. et al. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. Plant J. 12, 615–623 (1997).
Chen, W. et al. Male Sterile2 encodes a plastid-localized fatty acyl carrier protein reductase required for pollen exine development in Arabidopsis. Plant Physiol. 157, 842–853 (2011).
Morant, M. et al. CYP703 is an ancient cytochrome P450 in land plants catalyzing in-chain hydroxylation of lauric acid to provide building blocks for sporopollenin synthesis in pollen. Plant Cell 19, 1473–1487 (2007).
Dobritsa, A. A. et al. CYP704B1 is a long-chain fatty acid ω-hydroxylase essential for sporopollenin synthesis in pollen of Arabidopsis. Plant Physiol. 151, 574–589 (2009).
Dobritsa, A. A. et al. LAP3, a novel plant protein required for pollen development, is essential for proper exine formation. Sex. Plant Reprod. 22, 167–177 (2009).
Dobritsa, A. A. et al. LAP5 and LAP6 encode anther-specific proteins with similarity to chalcone synthase essential for pollen exine development in Arabidopsis. Plant Physiol. 153, 937–955 (2010).
Kim, S. S. et al. LAP6/POLYKETIDE SYNTHASE A and LAP5/POLYKETIDE SYNTHASE B encode hydroxyalkyl α-pyrone synthases required for pollen development and sporopollenin biosynthesis in Arabidopsis thaliana. Plant Cell 22, 4045–4066 (2010).
Tang, L. K., Chu, H., Yip, W. K., Yeung, E. C. & Lo, C. An anther-specific dihydroflavonol 4-reductase-like gene (DRL1) is essential for male fertility in Arabidopsis. New Phytol. 181, 576–587 (2009).
Grienenberger, E. et al. Analysis of TETRAKETIDE α-PYRONE REDUCTASE function in Arabidopsis thaliana reveals a previously unknown, but conserved, biochemical pathway in sporopollenin monomer biosynthesis. Plant Cell 22, 4067–4083 (2010).
Shi, J. et al. DEFECTIVE POLLEN WALL is required for anther and microspore development in rice and encodes a fatty acyl carrier protein reductase. Plant Cell 23, 2225–2246 (2011).
Zou, T. et al. OsLAP6/OsPKS1, an orthologue of Arabidopsis PKSA/LAP6, is critical for proper pollen exine formation. Rice 10, 53 (2017).
Zhu, X. et al. The polyketide synthase OsPKS2 is essential for pollen exine and Ubisch body patterning in rice. J. Integr. Plant Biol. 59, 612–628 (2017).
Xu, D. et al. Ostkpr1 functions in anther cuticle development and pollen wall formation in rice. BMC Plant Biol. 19, 104 (2019).
Li, J., Wang, Z., He, G., Ma, L. & Deng, X. W. CRISPR/Cas9-mediated disruption of TaNP1 genes results in complete male sterility in bread wheat. J. Genet. Genomics 47, 263–272 (2020).
Du, M. et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant J. 102, 1090–1100 (2020).
Metje-Sprink, J., Menz, J., Modrzejewski, D. & Sprink, T. DNA-free genome editing: past, present and future. Front. Plant Sci. 9, 1957 (2018).
Wada, N., Ueta, R., Osakabe, Y. & Osakabe, K. Precision genome editing in plants: state-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 20, 234 (2020).
Imai, R. et al. In planta particle bombardment (iPB): a new method for plant transformation and genome editing. Plant Biotechnol. (Tokyo) 37, 171–176 (2020).
Christensen, A. H., Sharrock, R. A. & Quail, P. H. Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 18, 675–689 (1992).
Ito, Y., Nishizawa-Yokoi, A., Endo, M., Mikami, M. & Toki, S. CRISPR/Cas9-mediated mutagenesis of the RIN locus that regulates tomato fruit ripening. Biochem. Biophys. Res. Commun. 467, 76–82 (2015).
Taniguchi, T., Konagaya, K.-I. & Nanasato, Y. Somatic embryogenesis in artificially pollinated seed families of 2nd generation plus trees and cryopreservation of embryogenic tissue in Cryptomeria japonica D. Don (Sugi). Plant Biotechnol. (Tokyo) 37, 239–245 (2020).
Konagaya, K.-I., Kurita, M. & Taniguchi, T. High-efficiency Agrobacterium-mediated transformation of Cryptomeria japonica D. Don by co-cultivation on filter paper wicks followed by meropenem treatment to eliminate Agrobacterium. Plant Biotechnol. (Tokyo) 30, 523–528 (2013).
Konagaya, K.-I., Nanasato, Y. & Taniguchi, T. A protocol for Agrobacterium-mediated transformation of Japanese cedar, Sugi (Cryptomeria japonica D. Don) using embryogenic tissue explants. Plant Biotechnol. (Tokyo) 37, 147–156 (2020).
Maruyama, E. & Hosoi, Y. Polyethylene glycol enhance somatic embryo production in Japanese cedar (Cryptomeria japonica D. Don). Propag. Ornam. Plants 7, 57–61 (2007).
Zhang, Z., Schwartz, S., Wagner, L. & Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7, 203–214 (2000).
Sayers, E. W. et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 50, D20–D26 (2022).
Jones, P. et al. InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240 (2014).
Ogden, E. C. & Raynor, G. S. A new sampler for airborne pollen: the rotoslide. J. Allergy 40, 1–11 (1967).
Nakamura, T., Yamada, K. D., Tomii, K. & Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 34, 2490–2492 (2018).
Acknowledgements
The authors are grateful to Ms. K. Nemoto, Ms. S. Tanaka, and Ms. A. Hagiwara for helpful assistance. This work was partly supported by Council for Science, Technology and Innovation (CSTI), Cross-ministerial Strategic Innovation Promotion Program (SIP), “Technologies for creating next-generation agriculture, forestry and fisheries” (funding agency: Bio-oriented Technology Research Advancement Institution, NARO), and by JSPS KAKENHI Grant Number JP20H03037.
Author information
Authors and Affiliations
Contributions
N.F. and M.N. designed the study and constructed the vectors. M.E., M.M., S.T., and S.K. were concerned in the construction of the vectors. Y.O., K.K., Y.N., and T.T. were involved in the transformation of sugi. T.E.M. regenerated the transgenic trees and treated them with gibberellin. M.N. analyzed the transgenes, flowers and pollen, and wrote the manuscript. All authors provided critical feedback for the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nishiguchi, M., Futamura, N., Endo, M. et al. CRISPR/Cas9-mediated disruption of CjACOS5 confers no-pollen formation on sugi trees (Cryptomeria japonica D. Don). Sci Rep 13, 11779 (2023). https://doi.org/10.1038/s41598-023-38339-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-38339-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
