
Abstract
Grapevine is a well-known host plant of the invasive pest Drosophila suzukii, but its susceptibility to pest oviposition and development greatly depends on the cultivar. To address environmental sustainability during viticultural zoning planning, new vineyard plantation and Integrated Pest Management programmes, it is essential to take pest pressure and cultivar susceptibility into account. To determine the different grapevine cultivars susceptibility to D. suzukii, we tested twelve widely spread cultivars during the ripening period. We also tested three cultivars during the drying period for raisin wine production. The infestation and emergence rates were consequently related to chemical and texture features of the berries to explain the role of skin and pulp characteristics in determining the nature of the susceptibility. Our results showed that susceptibility to D. suzukii infestation varies across cultivars. On ripening grapes, infestation is primarily influenced by skin and pulp firmness, elasticity and consistency. Suitability for egg development resulted mainly related to skin and pulp deformation. In a drying loft, infestation may also occur in relation to skin and pulp consistency. Lastly, we discuss the practical implication of the underestimated role of berry texture in D. suzukii oviposition and emergence success, in both ripening and drying grapes.
Similar content being viewed by others

Introduction
Newly introduced and invasive pests represent important challenges for crop production, and the magnitude of their detrimental effects should be accurately evaluated when a cultivar has to be selected. For grapevine, several differences in pest and disease incidence are reported according to cultivars and ripening stages1,2,3,4. Cultivar susceptibility to pests and diseases should thus be implemented within viticultural zoning programmes and in targeted Integrated Pest Management (IPM) strategies5,6,7. Differences among cultivars become relevant in viticulture because of a wide range of intrinsic physico-chemical characteristics, different ripening stages, grape uses, and wine destinations8,9.
Among grape pests, Drosophila suzukii Matsumura (Diptera: Drosophilidae) is a widespread and harmful species for ripe berries, due to its ability to lay eggs in healthy fruit close to the harvest period, the large number of generations per year, high reproductive capacity and wide host range10,11. Grapevine represents a host fruit for D. suzukii12,13,14,15,16, although it is affected by low oviposition, slow developmental rates, and limited survival to the adult stage (maximum 9%)13,17,18,19. Together with the direct damage caused by the larvae trophic activity, D. suzukii can facilitate D. melanogaster Meigen infestation20 and vehicle a large number of microorganisms, including acetic acid bacteria, favouring sour rot outbreaks18,20,21,22.
Infestations during ripening have been described in cultivars with soft skin16,23,24 confirming D. suzukii preference for fruits with thin skin and low penetrating resistance18,25,26,27,28,29,30,31. However, the skin resistance is only one aspect of the entire berry which is also characterized by a complex of anatomical and physical properties (e.g. skin firmness, pulp and berry consistency and elasticity). These properties, measured as mechanical response to applied forces, are used to characterize the cultivar differences of grapes and their quality32,33,34,35,36. The present study aims to test which among the anatomical and physical properties of the berry determines the higher cultivar susceptibility to infestation during the ripening period for all the winemaking productions. Understanding which cultivar and ripening stage are the most susceptible to oviposition would improve IPM practices and drive specific insecticide treatments and their optimal timing13.
In detail, this study was intended to identify the susceptibility to the infestation and development of D. suzukii in twelve highly valuable grape cultivars during the last phases of the ripening process and covering the entire harvesting period, by measuring the oviposition ability and emergence rate. In particular, the study aims to integrate a vast array of physical and mechanical proprieties related to berry texture and chemical features to identify the main drivers of susceptibility. For this reason, a penetrometer texture analysis was chosen to mimic the pest oviposition activity and its development.
In addition, grapes devoted to drying process, as for the production of valuable straw wines, may form a further substrate for D. suzukii oviposition and development, with a consequent detriment to the oenological process37. This drying process, which can last up to 100–120 days, is characterized by major changes of berry physical and chemical parameters. An accurate evaluation of the texture changes during the drying phases and their relationship with the incidence of pest infestation is needed to assess the vulnerability of these particular renewed traditional procedures. Three cultivars, suitable for drying process and devoted to the production of renowned raisin wines, were also analysed during postharvest, in order to test if the physio-chemical modifications that occur during the drying process would affect the success of D. suzukii infestation and development.
The proper characterization of cultivar susceptibility and analyses of environmental conditions are pre-requisite to design a rational D. suzukii control strategy within a wide grape growing area.
Results
Ripening grapes
Grape susceptibility to D. suzukii infestation
To evaluate the level of infestation we compared the percentage of infested ripening berries with the number of eggs laid per berry. Since these two indicators resulted highly correlated (n = 32, df=382, p-value < 0.0001, r2 = 0.50), the percentage of infested berries was used in all analyses as proxy for infestation level.
Significant differences in infestation were found among the tested grape cultivars (ANOVA, Linear Model: F11:307 21.95, p < 0.001) but not with collection time (F1:5 0.36, p = 0.55) or the interaction between cultivar x time (F11:307 1.22, p = 0.27; Fig. 1). The progress of maturation was also tested using chemical proxies to line up and monitor the different cultivars decoupling the observation from the collection date. However the analyses on sugar content, pH and total acidity showed no significant effects on infestation (F1:82 0.03, p = 0.86 for the sugar content, F1:82 2.47, p = 0.12 for pH and F1:82 0.05, p = 0.11 for total acidity, see also supplementary Fig. S1) also considering the interaction between these parameters and cultivar (F11:60 0.62, p = 0.81; F11:60 0.62, p = 0.80; F11:60 0.76, p = 0.67 for sugar content, pH and total acidity respectively), and further supporting the lack of a clear effect of time on infestation rate.

Level of infestation (percentage of infested berries) in the twelve tested cultivars over the whole period. The box edges enclose the first and third quartiles of the observations, the ends of the whiskers represent the 5th/95th percentiles, the solid line and cross indicate the median and mean respectively, the dots are the outliers. The different box colours indicate white-yellow (white) and blue-black (grey) cultivars.
Emergence rate showed low values in all the tested cultivars (Table 1) reaching maximum values (over 10%) in Cabernet sauvignon, Garganega, Glera, and Sauvignon blanc. Five cultivars presented fewer than 4%. In Barbera and Trebbiano di Soave no adults were obtained.
Texture and chemical features
The physical and mechanical features of the berries, related to the texture analyses, varied over time among the twelve cultivars. Instead, the chemical features showed a more constant cultivar-specific signature during the ripening period (supplementary dataset).
Relationships among entomological-mechanical-chemical features
The correlation analyses revealed a sizeable influence of texture features of berries on the susceptibility of infestation and larval development (Fig. 2). In particular, infestation level declines when the maximum force increases (r = −0.66, P < 0.001; Fig. 2) and when deformations at both maximum and minimum force increase (r = −0.51 and r = −0.40 respectively, P < 0.001; Fig. 2). On this regard, the twelve cultivars accurately cluster along the curve of the correlation between maximum force and infestation level (supplementary Figure S2). Among the texture features, emergence rate is highly correlated with gradient, deformation at maximum force, deformation at minimum force, minimum force and final force (Fig. 2).

Correlation chart in the texture-entomological analysis for ripening grapes. The frequency distribution and kernel density estimation of each variable is shown on the diagonal. The value of the Pearson’s correlation (r) and the significance level (p-value) corrected for multiple comparisons as asterisks (***<0.001, **<0.01, *<0.05, <0.10) are shown at the top of the diagonal; the size of the numbers is an indicator of the correlation value. The data range of values and units of measurement are given at the bottom of the diagonal.
On the contrary, no significant effects of chemical parameters were discovered (supplementary Fig. S1).
The grouping of the twelve cultivars is evident in PCA 2D-plot (Fig. 3), performed to depict the investigated mechanical texture variables, which explains with the first and second components 85.8% of total variability, thus very high. The spatial distribution of the twelve cultivars is consistent with the variables orientation imposed on the PCA plot. Sauvignon blanc, Glera, Pinot blanc, and Pinot gris are plotted in the area related to low values of internal forces in general, mainly maximum force. Merlot and Trebbiano di Soave are in the area characterized by high levels of maximum force.

Spatial arrangement of mechanical texture features investigated in the PCA analysis for grapes in the ripening process. Ellipses group the individual values of the cultivars (95% confidence).
Barbera, Corvina, Garganega and Syrah are characterized by high values of deformation, both at maximum and minimum force. Cabernet sauvignon is characterized by higher levels of final force and gradient, thus higher elastic properties, plotting in the low rate of infestation quadrant, defined by PC1.
Berry skin and pulp characteristics, derived from the specific mechanical texture analyses, are listed in Table 2 in order to summarize the level of infestation and emergence of the different cultivars in relation to the resulting phenotypic berry description.
Drying grapes
Grape susceptibility to D. suzukii infestation
As for ripening berries, also for drying grapes the percentage of infested berries was used as proxy for infestation level, because both number of infested berries and number of eggs laid per berry resulted highly correlated (n = 12, df=46, p-value<0.0001, r2 = 0.38).
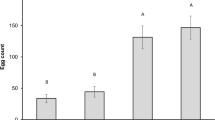
Merlot was never infested and discarded in the following analyses (Fig. 4). During the drying process, the grape cultivars presented differences in infestation over time (interaction time*cultivar F4:27 = 3.29, p = 0.025; Fig. 4). Garganega had low infestation levels at all the time points measured: less than 10% of berries infested on average, varying up to 20%. Corvina differed according to ripening level at harvest: overripe grapes (Or) were more susceptible to infestation than ripe ones, reaching values of 60% of infected berries (Fig. 4). At the end of the experiment, after 49 days of drying period, all cultivars resulted in low or zero infestation.

Infestation rate in the tested cultivars during the drying period. Corvina was also harvested overripe (Or). Error bars show standard error.
During the drying process, adults developed only from overripe grapes of Corvina cultivar with a high rate (Table 3).
Texture and chemical features
In the drying process, texture features varied among cultivars over time. It is interesting to note that maximum force remained stable during the whole drying period, while deformation at maximum force progressively increased with the dehydration of berries. Instead, the chemical features remained more constant during the drying period with the exception of sugar content that sizably increased (supplementary dataset).
Relationships among entomological-mechanical-chemical features
Similarly to the ripening process, the drying period was characterized by a significant effect of berry texture features impacting the susceptibility to infestation (Fig. 5).

Correlation chart in the texture-entomological analysis for grapes during the drying process. The frequency distribution and kernel density estimation of each variable is shown on the diagonal. The value of the Pearson’s correlation (r) and significance level (p-value) corrected for multiple comparisons as asterisks (***<0.001, **<0.01, *<0.05, <0.10) are shown at the top of the diagonal; the size of the numbers is an indicator of the correlation value. The data range of values and units of measurement are given at the bottom of the diagonal. The analyses of emergence percentage (separated from the other data by a bold line) were performed only in samples where more than 1 egg was laid (n = 6).
The infestation resulted correlated (P = 0.05) with deformation at maximum force and tended to inversely correlate with maximum force (P < 0.10; Fig. 5). The infestation percentage declined during the drying process as a result of the increase in deformation at maximum force. This pattern differed among the cultivars, ranging from the Corvina overripe grapes, characterized by low deformation and high susceptibility to infestation, to the Merlot which showed high deformation and no infestation (supplementary Fig. S3).
The emergence, analysed in a sub-sample where more than 1 egg was laid, showed no significant correlation with investigated features (Fig. 5).
Similarly to ripening grapes, none of the chemical parameters had any effect on infestation and larval development during drying period (supplementary Figure S4).
The grouping of the four harvested samples is evident in the PCA 2D-plot (Fig. 6) performed to depict the investigated mechanical texture proprieties. The spatial distribution of the samples appears consistent with the variable orientation imposed on the PCA plot. Merlot is plotted towards higher values of area, minimum force, maximum force and final force. The other three cultivars cluster together opposite to the high forces and area values.

Spatial arrangement of mechanical texture features investigated in the PCA analysis for grapes during the drying process. Ellipses group the individual values of the cultivars (95% confidence).
Discussion
The suitability of V. vinifera for infestation by D. suzukii is well established, while the driver in the susceptibility of different cultivars remains uncertain20. Grapevine can be a host plant for this pest12,13,14,15 and damage has been found in European countries on different white and black grape cultivars16,23,24. On the other hand, studies conducted by Lee et al.13, Bellamy et al.26 and Cai et al.38 found low attraction for grapes, suggesting that D. suzukii may prefer fruit with a thin skin. In addition, laboratory studies on grape berries reported low oviposition, slow developmental rates and limited survival to the adult stage13,17,18,19.
However, the infestation of D. suzukii in a vineyard determines economic losses and represents a sizable threat for quality wine production16,23,24,27. This apparent discrepancy can be explained by the different susceptibility to D. suzukii infestation of the various grape cultivars. It becomes pivotal to estimate such susceptibility to forecast economic repercussions of infestation and plan pest control strategies. Many aspects may contribute to the susceptibility for infestation in orchards such as fruit attractiveness (e.g. colour, odour and short-wavelength reflectance), microclimatic conditions, landscape characteristics and cultural practices (e.g. cultivar selection, plant nutrition, soil tillage, insecticide applications) and D. suzukii characteristics such as temperature-induced plasticity11,13,16,18,25,26,31,39,40,41,42,43,44,45,46,47. These aspects can also influence the transferability of laboratory results to the field situations22.
However, the essential element for infestation is reasonably the set of intrinsic characteristics of the berries since these features shape the fundamental micro-environment for egg oviposition and larval development. Thus, to determine whether the different susceptibility to infestation and adult emergence across grapevine cultivars is influenced by chemical and mechanical texture features of the berries, we tested twelve cultivars of renowned economic value during the ripening period useful for all winemaking productions. In addition, for three cultivars, the study was extended to the subsequent drying phase in a drying-loft for raisin wine production. The rates of infestation and emergence were related to chemical and mechanical texture features of the berries to explain the role of skin and pulp characteristics in determining the nature of the susceptibility.
The findings showed that the susceptibility to D. suzukii infestation, across the entire harvesting period, varies to a great extent among cultivars. Barbera, Garganega, Merlot and Trebbiano di Soave were infested only in a low percentage, thus showing the lowest level of susceptibility. Cabernet Sauvignon, Corvina and Syrah presented low levels of infestation (15% to 30% of berries infested on average). Chardonnay, Pinot blanc, and Pinot gris had medium-high infestation levels, up to 80% of infested berries. Glera and Sauvignon blanc presented higher levels, in some cases reaching complete berry infestation. Our results are partially comparable with those previously published on Chardonnay where 15.9% of infestation and 5.1% of emergence rate were found27 and on Cabernet sauvignon in open field conditions, characterized by average pest incidence of ~0.5 eggs/berry31. Differently from Ioriatti et al.27. where time showed an influence on susceptibility, in our experiment no significant changes in infestation were observed over time probably because our sampling period was placed in the last and more stable ripening phase, excluding, in particular the green stage of the fruit that might largely affect the variability of the forces, especially when it is measured as an end point value.
Furthermore, many studies underlined the role of chemical features of the grape berries in determining the rate of infestation27,29,30,48. However, these studies were addressed over the entire timespan of berry development from green to full ripening stage; thus, encompassing the major grapevine physiological transformations. Instead, our study focused specifically on the ripening period and reveals that the infestation and rate of emergence are not related to the analysed chemical factors (i.e. pH, sugar content and acidity) since these parameters remain stable and within the optimal range for D. suzukii development13,28. Other biochemical compounds, such us polyphenols, may have a role on larval development49.
On the other hand, we revealed that in the ripening period, characterized by the highest D. suzukii pressure, the texture features are primarily responsible for grape susceptibility to infestation. Indeed, berries with a firm and elastic skin and consistent pulp resulted less susceptible to D. suzukii oviposition. In addition, larval development is influenced by fruit texture, mostly due to the physical characteristics of the pulp and the consequent intrinsic forces.
Grape texture has been investigated in the last decade because of its importance in several contexts. Variety textural profile is important in grape blanching processes32, since its changes are closely related to cell-wall structure. Texture can affect processing events, like drying and fermentation50,51, but it also strongly influences sensory quality and consumers’ satisfaction when the fresh product is consumed52,53.
The rate of infestation resulted primarily influenced by toughness of the external tissue layers, like epidermis and hypodermis through the exocarp (expressed by maximum force), skin elasticity (gradient) and pulp consistency (deformation at minimum force); together these texture features have a sizable influence on cultivar susceptibility.
The most susceptible cultivar to D. suzukii infestation are Glera and Sauvignon blanc, which are characterized by low values of maximal force and consequently by soft berries. Chardonnay, Pinot blanc, and Pinot gris showed a medium-high infestation level. In detail, berries of these cultivars have a soft skin and low skin elasticity, that likely ease egg laying by D. suzukii. In Cabernet sauvignon, characterized by low skin elasticity, D. suzukii females lay eggs easily despite the relative high maximum force (firmer berry). On the contrary, Merlot berries, which present a tough skin, showed low infestation rate, similarly in Barbera which is characterized by high skin elasticity. In Corvina, Garganega, Syrah and Trebbiano di Soave the low infestation was related to mid-high or high skin firmness together with high skin elasticity.
The subsequent larval development is negatively correlated to skin and outer tissue layers elasticity and positively by pulp consistency, indicating that texture features influence the rate of success for the emergence of new adults. In detail, from our analysis the high deformation, as proxy for consistent pulp, is negatively correlated to the emergence, while high maximal force and high gradient, indices of low elastic skin, are positively correlated to pest development. Specifically, the highly deformable and elastic Barbera and Trebbiano of Soave likely prevent the development of eggs into new adults. The high values of deformation at maximum and minimum force, mainly related to the internal pulp structure recorded in Corvina and Syrah can explain the low emergence rate. Cabernet sauvignon, Chardonnay, Garganega, Glera and Sauvignon blanc, characterized by low values of these deformations, may grant high eggs development. Elastic skins and pulps can have a negative effect on the respiratory process of eggs and young larvae, possibly limiting the gas exchanges of preimaginal stages.
The cultivar selection operated by D. suzukii in the field can be also linked to other factors such as skin colour, with a clear preference for the red cultivars, and berries health condition18,22,27,44,46. In addition, biochemical molecules as those of odours released by the grape colonizing microorganisms may stimulate D. suzukii egg deposition20,21,22,44,54,55,56,57,58. Further studies should address whether the mechanical texture features of berries could influence the berries healthiness (e.g. development of damage or micro cracks) and the growth of such microorganisms.
Infestation may also occur in the drying loft, increasing the vulnerability of grape bunches to pathogens. Our results demonstrate that not all the tested cultivars are susceptible to D. suzukii infestation. In fact, oviposition never occurred on Merlot possibly because this cultivar is characterized by tough skin and consistent pulp. For the other cultivars, infestations during the drying process were lower than during ripening, reaching a maximum 60% of infested berries. Again, texture features contribute to the susceptibility of drying grapes to D. suzukii infestation. In particular, the maximum force and deformation at maximum force, indexes of skin and pulp consistency, represent the most effective obstacle to oviposition. Thus, firm but highly deformable berries tend to be less susceptible than those that were more turgid. Unlike in the ripening process, the duration of drying becomes a relevant factor in determining infestation rate. Indeed, higher infestation occurs during the early drying phase when the berries are swollen. The dehydration of berries associated with progressive drying decreases the D. suzukii capacity to lay eggs.
The effect of the softening of berries during drying is evident also on the low emergence rate since infested berries produced adults only in Corvina overripe sample.
In conclusion, our study demonstrates that the texture features of berries play a role in the susceptibility of grapevine to D. suzukii infestation and influence the D. suzukii oviposition and emergence success during the ripening process.
In addition to oenological and terroir facets, viticultural zoning programmes should include cultivar susceptibility to infestation in order to meet the level of pest pressure with the proper grapevine cultivar. It thus becomes worthwhile, when planning new vineyards to prefer less susceptible cultivars in vineyards subjected to major pest pressure, which are generally those exposed to high spillover from semi-natural habitats11,24,39,40. In addition, when implementing Integrated Pest Management strategies, chemical treatments can be limited and targeted to the vulnerable cultivars only41,59.
Regarding the drying process, the increase of skin elasticity, which also includes the sub-epidermis tissue layers, that progressively hampers D. suzukii oviposition, suggests adapting the environmental drying loft conditions in order to induce a rapid dehydration of berries. Even anticipating the grape picking, as in the case of the Corvina samples, can help in reducing the further development of D. suzukii in the drying loft. However, both these suggestions must be adapted to the particular grape characteristics and the oenological goals.
Methods
Collection of ripening grapes
Twelve highly valuable grape cultivars were investigated (Table 4). The grapevines, eleven years old, were growing in the experimental vineyard of the Soave wine cellar “Borgo Rocca Sveva” (45°25′10.4″N, 11°15′06.1″E) trained with Guyot system at a plant distance 2.5 m between rows and 0.8 within rows. Agronomic and phytosanitary practices were according to the local IPM programme (Veneto Region - Phytosanitary service)60; no insecticides were applied during the trial period.
Berries used in the bioassay were randomly collected from different vines and grapes. For each cultivar, samplings in the vineyard were performed weekly, from September 01st to October 13th, 2015 covering the typical harvesting periods (supplementary Table S1) and those for all winemaking productions (e.g. wine from late harvest, wines rich in tannins and spicy overtones, with complex, baked fruit or honeyed aroma) as suggested by the local vineries.
Collection of drying grapes
Boxes with grape bunches of three different cultivars (Corvina, Garganega and Merlot) were placed in a drying loft (located in Cazzano di Tramigna; 45°27′43.8″N, 11°12′20.6″E) to monitor infestation susceptibility and intrinsic characteristics during the drying process. Corvina, Garganega and Merlot grapes were harvested on September 28th. An additional overripe sample of Corvina was harvested on October 5th in order to encompass the variability of ripening stages that characterize the oenological uses of this cultivar. Samples of drying grapes were collected three times during the drying process: 21, 35, and 49 days after harvest.
Drosophila suzukii infestation
Experimental infestations of grape berries were artificially conducted in the laboratory at DAFNAE Department of the University of Padua, located in Legnaro (Italy). Experimental units were composed of five healthy and fully intact berries, randomly selected among more than 100, placed inside 7 × 7 × 7 cm net cages. The berries were sustained by their racemes to ensure the durability and integrity of fruit conditions. Each berry was previously checked under a stereo-microscope to exclude the presence of previous eggs, larvae and/or physical damage. Five females and two males of D. suzukii, 6 to 8 days-old were released inside the cages. The D. suzukii cohort used for the experiments was drawn from a long-lived colony raised in the DAFNAE laboratory and originated from adults collected from cherry, blueberry and grape in Verona district (Italy), as described in Tonina et al.61. Flies were nourished with a 10% sugar aqueous solution supplied in 2 ml plastic tubes stoppered with cotton. This unit was replicated four times for each cultivar. All flies were removed after 24 h of contact with berries and the presence of dead individuals was checked. The number of eggs laid on each berry was counted under stereo-microscope. Only the berries containing eggs were subsequently stored in plastic containers (100 ml) and kept in a climatic chamber at 22 °C, ~70% relative humidity, and a 16:8 hours (L:D) photoperiod. These berries were checked twice a week to assess adult emergence; the numbers of newly emerged adults were counted during the three following weeks.
Chemical analysis on grapes
Berries, from both the vineyard and the drying loft, were stored for no longer than 24 hours after collection at room temperature (21 ± 2 °C) before analysing. Physiological parameters were concurrently determined at each sampling on healthy berries for each tested cultivar by measuring the sugar content (°Brix) with a digital DBR35 refractometer, total acidity (meq/100 g) and pH using a titration Compact Titrator (Crison, Modena, Italy).
Texture measurements
A penetrometer texture analysis was chosen in order to mimic the oviposition activity of the pest and inferring the tissue and cellular opposing resistance to the insect ovipositor penetration and egg development in the fruit. For this reason, compression analyses were not considered because they mainly rely on the deduced indexes that we thought were not representative for our purpose. The texture analysis was performed with a Zwick texture analyser, Zwicki model (Zwick Roell Italia, Genova). All experiments were conducted at room temperature (21 ± 2 °C). For each replicate, twenty berries were analysed resulting in 20 individual measurements. Texture penetrometer tests were carried out as in Giongo et al.62, with a 12.56 mm2 area cylindrical probe, penetrating up to 99% fruit deformation at 0.3 mm/s and an accuracy of 0.001 N. Force vs. deformation curves were recorded and flesh rupture mean force penetration was calculated and corresponded to firmness expressed in Newton. Stress vs. strain curves were analysed and the gradient was obtained from the slope of the loading curve at the point of its highest gradient (supplementary Figure S5). Seven texture describing mechanical features were used to infer variability and genotypic profiles, as described in Table 5. All measurements were processed through the TextXpert II software, coupled to the machine.
Statistical analysis
All analyses and visualisations were implemented in R 3.4.163. To test the correlation between the percentage of infested berries and the number of eggs laid per berry, a Pearson’s correlation was used applying the function “cor.test” from the package “stats”64.
Through individual linear models the effects of grapevine cultivar over time on the rate of infestation, in both ripening and drying grapes, were tested. In each model, cultivar and data of collection (or drying length) were entered as categorical and linear factors respectively. For these analyses, the raw entomological datasets were used (see supplementary dataset). Percentage of infested berries was arcsine square-root transformed prior to analyses. In preliminary analyses, chemical features such as sugar content, pH and total acidity were tested to line up and monitor the progress of maturation. The assumptions of the models were evaluated by inspecting diagnostic plots of model residuals. The analyses were performed using the functions “lm” and “anova” from the package “nlme”65.
Correlation charts were conducted separately for ripening and drying grapes using the full entomological and chemical or texture mechanical data set (see supplementary dataset). Correlation chart reports the frequency distribution and kernel density estimation of each variable, the value of the Pearson’s correlation (r) and significance level (p-value) adjusted for multiple comparisons using Benjamini–Hochberg (BH) equation66. Correlation charts and relative analysis were performed using functions “chart.Correlation” and “corr.test” from the packages “PerformanceAnalytics”67 and “psych”68 respectively.
Principal Component Analysis (PCA) were conducted separately for ripening and drying grapes with the full texture mechanical data set (see supplementary dataset), using functions “prcomp” and “ggbiplot” from the packages “stats”64 and “ggbiplot”69 respectively.
Preliminary analyses were performed to determine variables with low loading on components.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Fornasiero, D., Pavan, F., Pozzebon, A., Picotti, P. & Duso, C. Relative infestation level and sensitivity of grapevine cultivars to the leafhopper Empoasca vitis (Hemiptera: Cicadellidae). Journal of Economic Entomology 109, 416–425 (2016).
Gabel, B. & Roehrich, R. Sensitivity of grapevine phenological stages to larvae of European grapevine moth, Lobesia botrana Den. et Schiff. (Lep., Tortricidae). Journal of Applied Entomology 119, 127–130 (1995).
Pavan, F. & Picotti, P. Influence of grapevine cultivars on the leafhopper Empoasca vitis and its egg parasitoids. BioControl 54, 55–63 (2009).
Sharon, R., Zahavi, T., Soroker, V. & Harari, A. R. The effect of grape vine cultivars on Lobesia botrana (Lepidoptera: Tortricidae) population levels. Journal of Pest Science 82, 187–193 (2009).
Basler, P. & Pfenninger, H. Disease-resistant cultivars as a solution for organic viticulture. In VIII International Conference on Grape Genetics and Breeding 603, 681-685 (2002).
Doster, M. A. & Schnathorst, W. C. Comparative susceptibility of various grapevine cultivars to the powdery mildew fungus Uncinula necator. American Journal of Enology and Viticulture 36, 101–104 (1985).
Fermaud, M. Cultivar susceptibility of grape berry clusters to larvae of Lobesia botrana (Lepidoptera: Tortricidae). Journal of Economic Entomology 91, 974–980 (1998).
Amati, A. et al. Influence of cultivar and cluster thinning on Valpolicella wine DOC phenolic composition. Rivista di Viticoltura e di Enologia 50, 37–46 (1997).
Du Plessis, C. S. Optimum maturity and quality parameters in grapes: a review. South African Journal of Enology and Viticulture 5, 34–42 (1984).
Cini, A., Ioriatti, C. & Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bulletin of Insectology 65, 149–160 (2012).
Kenis, M. et al. Non-crop plants used as hosts by Drosophila suzukii in Europe. Journal of Pest Science 89, 735–748 (2016).
Kanzawa, T. Studies on Drosophila suzukii Mats. (1939).
Lee, J. C. et al. The susceptibility of small fruits and cherries to the spotted‐wing drosophila, Drosophila suzukii. Pest Management Science 67, 1358–1367 (2011).
Seljak, G. Spotted wing drosophila-Drosophila suzukii (Matsumura). SAD, Revija za Sadjarstvo, Vinogradništvo in Vinarstvo 22 (2011).
Walsh, D. B. et al. Drosophila suzukii (Diptera: Drosophilidae): invasive pest of ripening soft fruit expanding its geographic range and damage potential. Journal of Integrated Pest Management 2, G1–G7, https://doi.org/10.1603/IPM10010 (2011).
Kehrli, P., Richoz, P., Linder, C. & Baroffio, C. The importance of Drosophila suzukii for grapevine production. IOBC-WPRS Bulletin 105, 211–218 (2014).
Maiguashca, F. et al. SWD ovipositing on grapes in laboratory; partial maggot survival inconclusive. Washington State University Extension, Spotted Wing Drosophila Grape Update, 28 August 2010, http://ipm.wsu.edu/small/pdf/NoChoiceSWDonGrapesAug28.pdf (accessed 23 August 2016) (2010).
Linder, C., Martin, C., Laboisse, S., Chatelain, P. G. & Kehrli, P. Susceptibility of various grape cultivars to Drosophila suzukii and other vinegar flies. IOBC-WPRS Bulletin 105, 219–224 (2014).
Atallah, J., Teixeira, L., Salazar, R., Zaragoza, G. & Kopp, A. The making of a pest: the evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proceedings of the Royal Society B: Biological Sciences 281, 20132840 (2014).
Rombaut, A. et al. Invasive Drosophila suzukii facilitates Drosophila melanogaster infestation and sour rot outbreaks in the vineyards. Royal Society Open Science 4, 170–117 (2017).
Ioriatti, C. et al. Drosophila suzukii (Diptera: Drosophilidae) contributes to the development of sour rot in grape. Journal of Economic Entomology 111, 283–292 (2017).
Entling, W. & Hoffmann, C. Single and combined effects of Drosophila suzukii and Drosophila melanogaster on sour rot development in viticulture. Journal of Applied Entomology https://doi.org/10.1111/jen.12721 (2019).
Griffo, R., Frontuto, A., Cesaroni, C. & Desantis, M. L’insetto Drosophila suzukii sempre più presente in Italia. L’Informatore Agrario 68, 56–60 (2012).
Marchesini, E., Mori, N. & Aldrighetti, F. Drosophila suzukii (Matsumura) a new pest of grape in Veneto Region (north-east Italy). IOBC-WPRS Bulletin 105, 229–232 (2014).
Burrack, H. J., Fernandez, G. E., Spivey, T. & Krausa, D. A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Management Science 69, 1173–1180 (2013).
Bellamy, D. E., Sisterson, M. S. & Walse, S. S. Quantifying host potentials: indexing postharvest fresh fruits for spotted wing drosophila, Drosophila suzukii. PLoS One 8, e61227 (2013).
Ioriatti, C. et al. Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. Journal of Economic Entomology 108, 1148–1155 (2015).
Lee, J. C. et al. Characterization and manipulation of fruit susceptibility to Drosophila suzukii. Journal of Pest Science 89, 771–780 (2016).
Pelton, E., Gratton, C. & Guédot, C. Susceptibility of cold hardy grapes to Drosophila suzukii (Diptera: Drosophilidae). Journal of Applied Entomology 141, 644–652 (2017).
Baser, N. et al. Susceptibility of table grape varieties grown in south‐eastern Italy to Drosophila suzukii. Journal of Applied Entomology 142, 465–472 (2018).
Entling, W., Anslinger, S., Jarausch, B., Michl, G. & Hoffmann, C. Berry skin resistance explains oviposition preferences of Drosophila suzukii at the level of grape cultivars and single berries. Journal of Pest Science 92, 477–484 (2019).
Wang, J. et al. Humidity hot air impingement blanching alters texture, cell-wall polysaccharides, water status and distribution of seedless grape. Carbohydrate polymers 194, 9–17 (2018).
Letaief, H., Rolle, L., Zeppa, G. & Gerbi, V. Assessment of grape skin hardness by a puncture test. Journal of the Science of Food and Agriculture 88, 1567–1575 (2008).
Letaief, H., Rolle, L. & Gerbi, V. Mechanical behavior of winegrapes under compression tests. American Journal of Enology and Viticulture 59, 323–329 (2008).
Rolle, L., Giacosa, S., Gerbi, V. & Novello, V. Comparative study of texture properties, color characteristics, and chemical composition of ten white table-grape varieties. American Journal of Enology and Viticulture 62, 49–56 (2011).
Rolle, L., Giacosa, S., Gerbi, V., Bertolino, M. & Novello, V. Varietal comparison of the chemical, physical, and mechanical properties of five colored table grapes. International Journal of Food Properties 16, 598–612 (2013).
Mori, N., Vincenzi, S., Marchesini, E., Curioni, A. & Duso, C. The effect of Drosophila suzukii attack during grape drying process in Veneto Region (north-eastern Italy). IOBC-WPRS Bulletin 105, 225–228 (2014).
Cai, P. et al. Potential host fruits for Drosophila suzukii: olfactory and oviposition preferences and suitability for development. Entomologia Experimentalis et Applicata 167, 880–890 (2019).
Santoiemma, G., Mori, N., Tonina, L. & Marini, L. Semi-natural habitats boost Drosophila suzukii populations and crop damage in sweet cherry. Agriculture, Ecosystems &. Environment 257, 152–158 (2018).
Tonina, L., Mori, N., Sancassani, M., Dall’Ara, P. & Marini, L. Spillover of Drosophila suzukii between noncrop and crop areas: implications for pest management. Agricultural and Forest Entomology 20, 575–581 (2018).
Audsley, N., Tonina, L. & Mori, N. Integrated management of tree fruit insect pests: Drosophila suzukii (Spotted Wing Drosophila). In Integrated Management of Diseases and Insect Pests of Tree Fruit. https://doi.org/10.1201/9780429266690 (2019).
Klick, J. et al. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. Journal of Applied Entomology 140, 37–46 (2016).
Santoiemma, G., Trivellato, F., Caloi, V., Mori, N. & Marini, L. Habitat preference of Drosophila suzukii across heterogeneous landscapes. Journal of Pest Science 92, 485–494 (2019).
Weißinger, L., Samuel, N., Breuer, M. & Müller, C. Effects of Variety and Grape Berry Condition of Vitis vinifera on Preference Behavior and Performance of Drosophila suzukii. Insects 10, 432 (2019).
Shawer, R., Tonina, L., Tirello, P., Duso, C. & Mori, N. Laboratory and field trials to identify effective chemical control strategies for integrated management of Drosophila suzukii in European cherry orchards. Crop protection 103, 73–80 (2018).
Little, C., Dixon, P., Chapman, T., & Hillier, N. Role of fruit characters and colour on host selection of boreal fruits and berries by Drosophila suzukii (Diptera: Drosophilidae). The Canadian Entomologist https://doi.org/10.4039/tce.2020.1 (2020).
Clemente, M., Fusco, G., Tonina, L. & Giomi, F. Temperature-induced phenotypic plasticity in the ovipositor of the invasive species Drosophila suzukii. Journal of thermal biology 75, 62–68 (2018).
Andreazza, F. et al. Susceptibility of grape vine genotypes by the infestation of Drosophila suzukii (Diptera: Drosophilidae). Pesquisa Agropecuária Brasileira 51, 599–606 (2016).
Jediyi, H., Naamani, K., Elkoch, A. A. & Lemjiber, N. Changes in grapes composition during ripening of five Vitis vinifera L. varieties as related to Tephritidae and Drosophilidae infestations. Physiology and Molecular Biology of Plants 25, 1407–1418 (2019).
Wang, J. et al. Pulsed vacuum drying of Thompson seedless grape: Effects of berry ripeness on physicochemical properties and drying characteristic. Food and Bioproducts Processing 106, 117–126 (2017).
Morata, A. et al. Emerging preservation technologies in grapes for winemaking. Trends in Food Science & Technology 67, 36–43 (2017).
Ma, C., Fu, Z., Xu, M., Trebar, M. & Zhang, X. Evaluation on home storage performance of table grape based on sensory quality and consumers’ satisfaction. Journal of Food Science and Technology 53, 1363–1370 (2016).
Giongo, L. et al. Raspberry texture mechanical profiling during fruit ripening and storage. Postharvest Biology and Technology 149, 177–186 (2019).
Becker, T. & Knoche, M. Deposition, strain, and microcracking of the cuticle in developing ‘Riesling’ grape berries. Vitis 51, 1–6 (2012).
Tasin, M. et al. Volatiles of grape inoculated with microorganisms: Modulation of grapevine moth oviposition and field attraction. Microbial ecology 76, 751–761 (2018).
Hamby, K. A. & Becher, P. G. Current knowledge of interactions between Drosophila suzukii and microbes, and their potential utility for pest management. Journal of Pest Science 89, 621–630 (2016).
Becher, P. G. et al. Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Functional Ecology 26, 822–828 (2012).
Entling, W. & Hoffmann, C. Drosophila suzukii and D. melanogaster and their impact on sour rot disease development in viticulture. Book of abstract IOBC/WPRS meeting “Integrated Protection in Viticulture” Portugal (2019).
Mori, N. et al. Integrated pest management approaches against Drosophila suzukii. Italus Hortus 26, 67–74 (2019).
Veneto Region - Phytosanitary service, Linee Tecniche di Difesa Integrata - Anno 2015. Regione del Veneto - Settore Fitosanitario DD n. 2015. https://www.regione.veneto.it/web/agricoltura-e-foreste/difesa-integrata [accessed 24 June 2015]
Tonina, L., Mori, N., Giomi, F. & Battisti, A. Development of Drosophila suzukii at low temperatures in mountain areas. Journal of Pest Science 89, 667–678 (2016).
Giongo, L., Poncetta, P., Loretti, P. & Costa, F. Texture profiling of blueberries (Vaccinium spp.) during fruit development, ripening and storage. Postharvest Biology and Technology 76, 34–39 (2013).
R Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org (2017).
R Core Team and contributors worldwide. The R Stats Package, https://stat.ethz.ch/R-manual/R-devel/library/stats/html/00Index.html (2019).
Pinheiro, J., Bates, D., DebRoy, S. & Sarkar, D. R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-142, https://CRAN.R-project.org/package=nlme (2019).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Statist Soc B 57, 289–300 (1995).
Peterson, B.G. et al. PerformanceAnalytics: Econometric Tools for Performance and Risk Analysis, https://cran.r-project.org/web/packages/PerformanceAnalytics/index.html (2020).
Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research, https://cran.r-project.org/web/packages/psych/index.html (2020).
Vu, Q., V. ggbiplot: Biplot for Principal Components using ggplot2, https://www.rdocumentation.org/packages/ggbiplot (2011).
Calò, A., Scienza, A. & Costacurta, A. Vitigni d’Italia. Calderini Edagricole (2001).
Acknowledgements
This work was funded by Servizio Fitosanitario Regione Veneto within the project “Monitoraggio ed impostazione di una razionale strategia di controllo contro Drosophila suzukii” for LT and MS for the entomological part. Moreover, the research leading to these results has received funding from the European Union’s Seventh Framework programme for research, technological development and demonstration under grant agreement number 613678 (DROPSA). In addition, texture analyses were supported by PAT (Provincia Autonoma di Trento) through AdP funding for LG and MA. We are grateful to Giuseppe Rama of “Cantina di Soave” for access to the cultivars collection vineyard and the drying loft. We also thank Enrico Marchesini for his field technical support; Alberto Grassi and Angela Gottardello for their help in the experiment design and berry damage lab assessment.
Author information
Authors and Affiliations
Contributions
L.T., M.S., N.M. and L.G. conceived and designed the research. L.T., M.S. and M.A. conducted the experiments. L.T., F.G. and L.G. analysed the data. L.T., F.G., N.M. and L.G. wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tonina, L., Giomi, F., Sancassani, M. et al. Texture features explain the susceptibility of grapevine cultivars to Drosophila suzukii (Diptera: Drosophilidae) infestation in ripening and drying grapes. Sci Rep 10, 10245 (2020). https://doi.org/10.1038/s41598-020-66567-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-66567-9
This article is cited by
-
A menu of measures to manage trade-related plant pest risks, and a review of methods for demonstrating measure efficacy
Biological Invasions (2023)
-
Drosophila suzukii energetic pathways are differently modulated by nutritional geometry in males and females
Scientific Reports (2022)
-
Drosophila suzukii preferentially lays eggs on spherical surfaces with a smaller radius
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
