
Abstract
Despite concerted international effort to track and interpret shifts in the abundance and distribution of Adélie penguins, large populations continue to be identified. Here we report on a major hotspot of Adélie penguin abundance identified in the Danger Islands off the northern tip of the Antarctic Peninsula (AP). We present the first complete census of Pygoscelis spp. penguins in the Danger Islands, estimated from a multi-modal survey consisting of direct ground counts and computer-automated counts of unmanned aerial vehicle (UAV) imagery. Our survey reveals that the Danger Islands host 751,527 pairs of Adélie penguins, more than the rest of AP region combined, and include the third and fourth largest Adélie penguin colonies in the world. Our results validate the use of Landsat medium-resolution satellite imagery for the detection of new or unknown penguin colonies and highlight the utility of combining satellite imagery with ground and UAV surveys. The Danger Islands appear to have avoided recent declines documented on the Western AP and, because they are large and likely to remain an important hotspot for avian abundance under projected climate change, deserve special consideration in the negotiation and design of Marine Protected Areas in the region.
Similar content being viewed by others


Introduction
Monitoring populations is essential to species conservation, and can be used to identify threats or changes in conservation status. Indeed, central to the Convention on Biological Diversity and related conservation measures is the assumption that we can quantify species diversity, abundance, and geographic distribution1,2. Numerous studies now suggest that Adélie penguin (Pygoscelis adeliae) populations are undergoing dramatic shifts in abundance, with marked declines along most of the Western Antarctic Peninsula (WAP) and associated sub-Antarctic Islands3,4,5,6,7,8 and sharp increases in the Ross Sea and Eastern Antarctica8,9,10,11,12,13,14. While the causal drivers of these changes remain unknown and may in fact vary across the continent, several studies have linked Adélie penguin population trends to changes in sea ice extent and concentration as well as changes in air temperature and precipitation patterns and their possible effects on prey availability14,15,16,17. Understanding the population dynamics of sentinel species, such as the Adélie penguin, will help illuminate the effects of climate change on less easily-studied components of the ecosystem18,19. Much of the concern regarding climate-driven changes has been focused on the WAP and South Shetland/South Orkney Islands, where Adélie penguin populations have declined sharply (~70%) over the last several decades20,21. Along the WAP, the northern end of Marguerite Bay (67°30′S) represents a clear boundary that divides areas of Adélie penguin population decline in the north from areas where abundances are either stable or increasing to the south22. Far less is known about Adélie penguin populations along the northern and eastern portions of the AP, a region perhaps more closely tied to the Weddell Sea in terms of climate and sea ice production than to the dynamics of the WAP23,24.
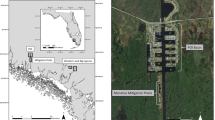
The Danger Island archipelago is comprised of 9 islands stretching over approximately 35 km at the northernmost tip of the AP in the north-western Weddell Sea (Fig. 1). Despite their relative proximity to the WAP, which sees much of the tourist and fishing ship traffic in the Antarctic25, pack ice is common around the Danger Islands even in austral summer26. In fact, due to the currents of the Weddell Sea, which drive sea ice northward, access to the islands is precluded in most years. Heroína Island, at the northeast end of the archipelago, is the most frequently visited of the Danger Islands and yet hosts a median visitation rate of only one ship landing per year27. It is also the only island to date with a population estimate (285,000–305,000) derived from a ground survey of the island28,29. While a previous geological expedition30 noted the presence of Adélie penguins on all of the Dangers Islands (with the exception of Darwin Island, which was not visited) and several others were photographed (by M.R. and S.F.) from a passing vessel in 2008/09, the presence of Adélie penguins on several of these islands went largely unrecognized until a recent Landsat satellite survey of the Antarctic identified several large penguin colonies supporting what appeared to be nearly 200,000 Adélie penguin nests31. The (re)discovery of these populations, combined with evidence that the Danger Islands as a group supported a regionally-significant population, motivated an expedition to the area. In this paper we report on the first comprehensive seabird survey of the Danger Islands and describe a heretofore unrecognized Adélie penguin hotspot. We document a novel multi-modal survey comprised of ground surveys and imagery from both satellites and unmanned aerial vehicle (UAV) photographic surveys, the results of which were combined with historic aerial photographs to assess long-term change in the region. We also discuss the implications of this seabird hotpot for the design of Marine Protected Areas in the Antarctic Peninsula region.

(a) Map showing the location of the Antarctic Peninsula and (b), the location of the Danger Islands group on the Antarctic Peninsula, both created using ESRI ArcMap 10.0 (http://desktop.arcgis.com/en/arcmap/). (c) Quickbird image of the Danger Islands taken 22 January 2011 (©2018, DigitalGlobe).
Results
Our survey found 751,527 (95th CI = [710,103–792,443]) nesting pairs of Adélie penguins in the Danger Islands (Table 1). When combined with known information on abundance elsewhere in the region21, we estimate the Danger Islands contain 55% of all Adélie penguins in subarea 48.1 as defined by the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR). This subarea includes all of the western AP, the waters north of the AP to 60°S, and a portion of the north-western Weddell Sea west of 50°W. Without the Danger Islands, 48.1 contains 606,526 (95th CI = [322,477–990,402]) Adélie penguin pairs21.
Visual comparison of available aerial, satellite, and unmanned aerial vehicle (UAV) images suggests that the area occupied by Adélie penguin colonies on the Danger Islands has remained stable or has modestly increased over the last 60 years, though our inference regarding dynamics is unavoidably limited by the lack of imagery between 1957 and 1990 (Fig. 2). A previous ground-based estimate28 of 285,000–305,000 Adélie penguin nests on Heroína Island in 1996/97 is remarkably consistent with our updated estimate of 292,363 nests. The consensus of all the data considered in this analysis strongly suggests that the Danger Islands have remained roughly stable since the earliest records of the 1950s, in stark contrast to declines seen along the WAP.

Guano areas (yellow) identified on Brash Island (at left) and Heroína Island (at right) from 1957 to present day. (a) and (h) manually classified from aerial imagery from 1957; (b) and (i) manually classified from aerial imagery from 1957 and reduced to 30 m cells for comparison with Landsat; (c) and (j): Landsat-4 in 1990 classified as described in Methods; (d) and (k) Landsat-7 in 2000 classified as described in the Methods; (e) and (l) Landsat-8 in 2015 classified as described in the Methods; (f) Worldview-2 image taken 19 February 2016 classified as described in the Methods (©2018, DigitalGlobe); (m) Worldview-2 image taken 2 December 2015 classified as described in the Methods (©2018, DigitalGlobe); (g) and (n) nests that were retained by the spatial filter marked as yellow dots overlaid on UAV imagery from ground survey described in this manuscript. Panels a, b, f, g, h, i, m, and n displayed using ESRI ArcMap 10.0 (http://desktop.arcgis.com/en/arcmap/); Panels c, d, e, h, k, and l displayed using ENVI 5.4 (https://www.harris.com/solution/envi).
In addition to Adélie penguins, we found several populations (>100 nests) of gentoo penguins (P. papua), particularly at Brash Island, and one small population (27 nests) of chinstrap penguins (P. antarctica) at Heroína Island. Additional information on flying birds and marine mammal observations collected during this survey are included in Supplementary Information Table S1.
Discussion
This survey provides the first estimates of penguin abundance for this portion of the AP region and the first direct ground survey of the Danger Islands beyond Heroína Island, which was last surveyed in 1996/97. Our estimate is more than three times the abundance estimated by an earlier survey8, largely because several colonies, not known to exist at the time, were missed entirely. We find the Adélie penguin colonies on Heroína Island and Beagle Island are the third and fourth largest Adélie penguin colonies in the world21, respectively, and represent the easternmost Antarctic colonies (54°W) of all three pygoscelid penguins until 45°E. Our ground- and UAV-derived survey provides important validation of satellite imagery as a tool for the discovery of new penguin colonies, and demonstrates how satellite imagery and field expeditions can be used in concert to track penguin biogeography and long-term trends. Our discovery of a major hotspot of abundance in the Danger Islands is important for our understanding of the global distribution of the Adélie penguin, and should be considered in the development of future conservation measures such as Antarctic Specially Protected Areas (ASPAs) or Marine Protected Areas (MPAs).
At a regional scale this survey increases the total estimated abundance of Adélie penguins in CCAMLR subarea 48.1 by 68%. This dramatic increase in the number of known Adélie penguin breeding pairs radically changes our estimates of krill predation in the Northern Weddell Sea, in a portion of 48.1 that, notably, has not experienced the levels of krill fishing seen elsewhere along the Antarctic Peninsula25,32. The Adélie penguins found in the Danger Islands are among those penguins breeding north of the “Adélie gap,” a stretch of the coast along the WAP roughly 400 km long from the Adélie colonies on the southwestern shore of Anvers Island to the south, to Nelson Island in the South Shetland Islands and nearly the tip of the AP to the north33,34. This gap is devoid of breeding Adélie penguins and geographically differentiates those Adélie penguins to the south from those in the north in both summer and winter foraging habitat34. The new abundance estimate for the northern portion of subarea 48.1 highlights the spatial structure of predator abundance, and reveals an area of high abundance distinct from the better surveyed coastline of the WAP. Accordingly, an updated understanding of predator distributions, particularly the location of major abundance hotspots, may have implications for the management of prey resources35,36.
While our inference on past trends is unavoidably limited by the lack of prior ground surveys, our analysis of the available imagery suggests that Adélie penguin colonies in the Danger Islands have not suffered the net declines seen on the WAP, where some colonies have declined by an order of magnitude or even disappeared completely14,37,38. Our findings are consistent with recent modelling work39 showing that the warming of the WAP has followed a west-to-east pattern, such that the Danger Islands have been largely spared the environmental changes experienced by the South Shetland Islands and the northern portion of the WAP. In particular, the Weddell Sea has not experienced the significant loss of sea ice seen in the Bellingshausen Sea, and instead shows slight gains over the past several decades40,41, providing more consistent foraging habitat for pagophilic species such as the Adélie penguin. We recognize, however, that while the evidence for stability from 1990-present is well supported by the available imagery, the evidence cannot rule out sequential and roughly compensatory periods of increase and decrease in the earlier period (1957–1990)42.
Given the large number of Adélie penguins breeding in the Danger Islands, and the likelihood that the northern Weddell Sea will remain suitable for Adélie penguins longer than the rest of the Antarctic Peninsula region, we suggest the Danger Islands should be strongly considered for further protection, either through an extension of the proposed Weddell Sea MPA that falls just to its south or by way of an MPA in the Western Antarctic Peninsula43,44. Like the Ross Sea, the northern Weddell Sea represents an Adélie penguin hotspot of significant conservation value as a potential refugium under climate change. By establishing the distribution and abundance of penguins in this region, we hope to highlight its importance to regional and global populations, and encourage more regular monitoring of the region.
Methods
We define the Danger Islands as including (from north to south): Brash Island, Heroína Island, Comb Island (also known as Peine Island), Beagle Island, Platter Island (Plato Island), Darwin Island, and Earle Island (Fig. 1). Dixey Rock and Scud Rock are also located in this area and were also surveyed. These islands range from generally low and flat (Platter Island), to sheer cliff faces (Darwin and Comb Islands), with most containing a mix of steep scree slopes, flat areas, and cliffs. The islands are composed of intrusive igneous rocks, predominantly feldspar-rich gabbro, that were formed during the late Cretaceous and are of similar age to the plutonic rock formations at the tip of the Antarctic Peninsula45. Data from this region are sparse, but during the last glacial maximum these islands may have been glaciated until around 6000 years before present (bp)46. While the Holocene occupation history of penguins on the Danger Islands is currently undescribed, radiocarbon-dated remains from other northern Antarctic Peninsula breeding sites indicates a relatively recent (~600 bp) advent of breeding populations47.
Field survey
Surveys were conducted from the M/V Hans Hansson from December 9–18, 2015. We used a variety of survey methods on each island depending on conditions and time ashore, including one or more of the following methods: (1) manually counting individual nests, (2) counting individual nests in panoramic photos taken from the ground or the vessel, and (3) counting individual penguins from photographs captured by UAV. The combination of these methods allowed for efficient data collection with opportunities for cross-validation of survey methods. The precision of census counts varied by island (Table 1), and island-specific error estimates were propagated to the archipelago-wide confidence intervals for total abundance.
The timing of our expedition was ideal in terms of penguin phenology48, and the surveyed colonies were dominated by individual penguins incubating well-established nests. Our estimates of abundance, therefore, represent a count of all ‘actively’ incubated nests on each island. Active nests were those occupied by a penguin, noting that the presence of eggs or chicks in the nest cannot be determined from aerial photography. To facilitate counting, islands were divided based on the natural boundaries between “sub-colonies,” or naturally-occurring, discrete groups of penguin nests. For those sub-colonies too large to count accurately in their entirety, divisions were made based on natural markers within the sub-colony or, in their absence, using brightly coloured rope laid between nests; such subsections were counted individually. To ensure accuracy, each division was counted three times and these three counts were required to agree within 5% of their mean. If counts did not agree, divisions were further subdivided until three subsequent counts did agree within 5% of the mean. This 5% accuracy threshold corresponds to the ‘N1’ level of precision described by Croxall and Kirkwood49 and used regularly to report penguin abundance in the Antarctic (e.g., refs28,33,50).
Full, site-wide counts for Adélie penguins were conducted at Platter and Earle Islands, the latter of which was also surveyed by UAV (Table 1). Manual counts of well-defined Adélie penguin sub-colonies were conducted at Heroína, Brash, and Beagle Islands as a validation of counts based on UAV photographs. Where present, we conducted site-wide counts of gentoo penguins and chinstrap penguins as well (Table 1). Adélie penguin populations at Comb and Darwin Islands were counted from images taken on the ground or from vessels offshore of the islands using Adobe Photoshop’s count tool. On all islands, the presence of other bird or mammal species was noted opportunistically (Supplementary Information Table S1).
All research was conducted with under the approval of Stony Brook University’s Institutional Animal Care and Use Committee (237420), Woods Hole Oceanographic Institution’s Institutional Animal Care and Use Committee (18958), and following ethical review by the University of Oxford. This expedition was permitted under Antarctic Conservation Act Permit ACA 2016-011. An Initial Environmental Evaluation was approved by the US Environmental Protection Agency on 1 December 2015, and Advance Notification provided to the US Department of State.
UAV-based survey
UAV surveys were performed with a DJI Phantom 3 quadcopter using its stock 1.2 Megapixel camera. The UAV was flown either manually or automatically using the mission planning software Map Pilot App to generate image coverage of each island with at least 70% overlap between images. Following the suggestions laid out by ref.51, a minimum height above ground of 25 m was set for all flights to avoid disturbance to wildlife, and a maximum height above ground of 45 m was selected to maintain image quality for penguin identification. The geotagged imagery collected with the UAV was post-processed using the commercial photogrammetry software Photoscan (Agisoft LLC, St. Petersburg, Russia) which generated full, georeferenced orthomosaics, a top-down view of the island – in which each pixel corresponds to a fixed physical dimension – of the surveyed islands and their penguin colonies.
Brash, Earle, Beagle, and Heroína Islands were surveyed using composite panoramic images captured by the UAV (example in Fig. 3). The timing of our survey was ideal for capturing incubating penguins on the nest and the imagery was, in the overwhelming majority of cases, unambiguous with respect to penguins that were incubating versus walking through the colony or from the ocean. To automatically identify and count the number of occupied nests in the UAV orthomosaics we used a Deep Neural Network (DetectNet) implemented in the open source software NVIDIA DIGITS (NVIDIA Corporation, Santa Clara, CA). DetectNet is based on the GoogLeNet image classification framework52 and is specifically designed to locate multiple objects of the same type within an image, making it well-suited to the task of detecting penguins in aerial imagery. The DetectNet network was trained to detect penguins using 512 × 512 sub-images selected from the orthophotos of the four islands being analysed and manually annotated with penguin locations. The images were split into two groups, one for training the network and one for validation, with 160 images and 1237 penguins in the training group and 93 images and 673 penguins in the validation group. The manually-labelled training data constituted 0.18% of the imaged area and 0.34% of the imaged penguins providing a massive decrease in manual labour required. Once trained, full island detection was performed by splitting the orthophotos into 512 × 512 sub-images which were run through the trained detector in DIGITS.

UAV orthomosaic image of Brash Island (above), with examples of zoomed-in penguin rookeries (below), displayed using ESRI ArcMap 10.0 (http://desktop.arcgis.com/en/arcmap/).
False positives generated by this automated nest detection algorithm were comprised of both individual non-nesting penguins and artefacts, such as rocks and shadows, that appear visually similar to nesting penguins. While nesting penguins are highly spatially structured, with strong attraction between individuals53, the false positives are largely spatially unstructured. We therefore applied an additional spatial filter to the detections, retaining nests based on the distribution of nearest neighbours. Points passing through this spatial filter are classified as unambiguous penguin nests and are retained; points rejected at this stage are comprised of false positives and a much smaller number of isolated nests incorrectly rejected by the filtering process. We validated our nest detections by manually counting a selection of each of the four islands surveyed by UAV (Brash, Earle, Beagle, Heroína), and created a simple linear regression model to estimate the number of nests based on the number detected (Supplementary Methods S1). These site-specific linear models allowed us to correct for any site-specific differences in the performance of the detection algorithm. Based on this analysis, we have classified our automated counts as \(\pm 10 \% \) accuracy (i.e. an ‘N2’ count), though we note that the average difference between the automated nest counting of UAV imagery and an in situ ground count of the same portion of the colony was only 0.6% and so our estimates may be even more precise than suggested by an ‘N2’ designation.
Historical aerial imagery
To understand the potential population trajectory of penguins in the Danger Islands region, the spatial extent of current penguin colonies can be qualitatively compared to historical aerial photographs (Fig. 2). We selected cloud-free photographs from the Falkland Islands Dependencies Aerial Survey Expedition (FIDASE) for Heroína54 and Brash55 islands. These islands were chosen as the FIDASE archive contained quality photographs shot nearly on nadir and both had been fully photographed by the UAV. The islands were photographed in black and white on Jan. 31, 1957 at an altitude of 4115 m to a scale of 1:27000, and digitally scanned by the U.S. Geological Survey. Both island frames were georeferenced to WorldView-2 satellite images and divided into polygons using segmentation algorithms tuned to provide a segment size reasonable for further analysis using Quantum GIS56 (QGIS). In both cases, segments were manually classified by a skilled observer as “guano” to designate recent guano deposition, “guano-like” to designate areas thought to be guano but with less certainly, “old guano” to designate areas of guano accumulation not necessarily associated with active nesting, and “non-guano,” a classification encompassing rock, water, snow, and all other substrates not covered by guano (Fig. 2).
Landsat satellite imagery analysis
Adélie penguin colonies identified in Fig. 2 were retrieved from Landsat imagery based on the algorithm described by refs31,57. The retrieval algorithm was originally developed for a single sensor and was modified in this case to operate on cross-calibrated data from Landsat-4 (imagery date 1990 in Fig. 2), −7 (2000) and −8 (2015). Cross-calibration among sensors was performed by calculating the mean difference of similar bands from Landsat-4 and-8 imagery compared to Landsat-7, and then adjusting the band values based on the mean differences in each spectral band. The algorithm was then applied to the cross-calibrated imagery to classify Adélie penguin colony areas.
High-resolution commercial imagery
Areas of guano staining were manually identified in high resolution satellite imagery (Fig. 2 panels f and m). A selection of unambiguous pixels within the guano stains were used to select other pixels (using Adobe Photoshop) similar in colour, and areas were added and removed manually based on manual interpretation of the imagery combined with auxiliary information from the UAV imagery mosaics.
Data availability
All Landsat data are archived by the U.S. Geological Survey. The data can be acquired at no cost via EarthExplorer (https://earthexplorer.usgs.gov/) and several other on-line tools hosted by the USGS. High resolution commercial satellite imagery (e.g., Worldview, Quickbird) is subject to a licensing agreement with Digital Globe, Inc. and inquiries should be directed to the Polar Geospatial Center (www.pgc.umn.edu/). UAV image mosaics, ArcGIS shapefile layers for penguin nests, and additional photographic data collected during this expedition may be obtained from the authors on request.
References
Agnew, D. J. The CCAMLR ecosystem monitoring programme. Antarct. Sci. 9, 235–242 (1997).
Chown, S. L. et al. Antarctica and the strategic plan for biodiversity. PLoS Biol 15, e2001656 (2017).
Trivelpiece, W. Z. & Fraser, W. R. The breeding biology and distribution of Adélie penguins: adaptations to environmental variability. Antarct. Res. Ser. 70, 273–285 (1996).
Fraser, W. R. & Patterson, D. L. Human disturbance and long-term changes in Adélie penguin populations: a natural experiment at Palmer Station, Antarctic Peninsula in Antarctic communities: Species, structure and survival (eds. Battaglia, B., Valencia, J. & Walton, D. W. H.) 445-452 (Cambridge University Press, 1997).
Sander, M., Balbão, T. C., Polito, M. J., Costa, E. S. & Bertoldi Carneiro, A. P. Recent decrease in chinstrap penguin (Pygoscelis antárctica) populations at two of Admiralty Bay’s islets on King George Island, South Shetland Islands, Antarctica. Polar Biol. 30, 659–661 (2007).
Forcada, J. & Trathan, P. N. Penguin responses to climate change in the Southern Ocean. Glob. Change Biol. 15, 1618–1630 (2009).
Lynch, H. J., Fagan, W. F. & Naveen, R. Population trends and reproductive success at a frequently visited penguin colony on the western Antarctic Peninsula. Polar Biol. 33, 493–503 (2010).
Lynch, H. J. & LaRue, M. A. First global survey of Adélie penguin populations. The Auk 131, 457–466 (2014).
Croxall, J. P., Trathan, P. N. & Murphy, E. J. Environmental change and Antarctic seabird populations. Science 297, 1510–1514 (2002).
Ainley, D. G. et al. Letter to theEditor: Adélie penguins and Environmental Change. Science 300, 429–430 (2003).
Jenouvrier, S., Barbraud, C. & Weimerskirch, H. Sea ice affects the population dynamics of Adélie penguins in Terra Adélie. Polar Biol. 29, 413–423 (2006).
Lyver, P. O. et al. Trends in the breeding population of Adélie Penguins in the Ross Sea, 1981–2012: A coincidence of climate and resource extraction effects. PLoS ONE 9, e91188, https://doi.org/10.1371/journal.pone.0091188 (2014).
Southwell, C. et al. Spatially-extensive standardized surveys reveal widespread, multi-decadal increase in East Antarctic Adélie penguin populations. PLoS ONE 10, e0139877, https://doi.org/10.1371/journal.pone.0139877 (2015).
Che-Castaldo, C. et al. Pan-Antarctic analysis aggregating spatial estimates of Adélie penguin abundance reveals robust dynamics despite stochastic noise. Nat. Comm. 8, 832, https://doi.org/10.1038/s41467-017-00890-0 (2017).
Ainley, D. G. et al. Decadal-scale changes in the climate and biota of the Pacific sector of the Southern Ocean, 1950s to 1990s. Antarctic Sci. 17, 171–182 (2005).
Forcada, J., Trathan, P. N., Reid, K., Murphy, E. J. & Croxall, J. P. Contrasting population changes in sympatric penguin species in association with climate warming. Glob. Change Biol. 12, 411–423 (2006).
Massom, R. A. et al. Extreme anomalous atmospheric circulation in the West Antarctic Peninsula region in austral spring and summer 2001/02, and its profound impact on sea ice and biota. J. Clim. 19, 3544–3571 (2006).
Trivelpiece, W. Z., Trivelpiece S. G., Geupel, G. R., Kjelmyr, J. & Volkman, N. J. Adelie and chinstrap penguins: their potential as monitors of the Southern Ocean marine ecosystem in Antarctic Ecosystems (eds. Kerry, K. R. & Hempel, G.) 191–202 (Springer, 1990).
Ainley, D. G. The Adélie Penguin: Bellwether of Climate Change (Columbia University Press, 2002).
Lynch, H. J., Naveen, R. & Casanovas, P. V. Antarctic Site Inventory breeding bird survey data 1994/95-2012/13. Ecology 94, 2653 (2013).
Humphries, G. R. W. et al. Mapping Application for Penguin Populations and Projected Dynamics (MAPPPD): Data and tools for dynamic management and decision support. Polar Rec. 53, 160–166 (2017).
Casanovas, P., Naveen, R., Forrest, S., Poncet, J. & Lynch, H. J. A comprehensive coastal seabird survey maps out the front lines of ecological change on the Western Antarctic Peninsula. Polar Biol. 38, 927–940 (2015).
Ducklow, H. W. et al. Marine pelagic ecosystems: the west Antarctic Peninsula. Philos. Trans. R. Soc. London, Ser. B 362, 67–94 (2007).
Murphy, E. J. et al. Spatial and temporal operation of the Scotia Sea ecosystem: a review of large-scale links in a krill centred food web. Philos. Trans. R. Soc. London, Ser. B 362, 113–148 (2007).
CCAMLR. Krill fishery report 2016 (Commission for the Conservation of Antarctic Marine Living Resources, 2016).
Comiso, J. C. & Gordon, A. J. Interannual variability in summer sea ice minimum, coastal polynyas and bottom water formation in the Weddell Sea. Antarctic Sea Ice: Physical Processes, Interactions and Variability. Antarctic Res. Ser. 74, 293–315 (1998).
IAATO Tourism statistics. International Association of Antarctica Tour Operators. https://iaato.org/tourism-statistics (2017).
Naveen, R., Forrest, S. C., Blight, L. K., Trivelpiece, W. Z. & Trivelpiece, S. G. Censuses of penguin, blue-eyed shag, and southern giant petrel populations in the Antarctic Peninsula region, 1994-2000. Polar Rec. 36, 323–334 (2000).
Lynch, H. J., Naveen, R. & Fagan, W. F. Censuses of penguin, blue-eyed shag, Phalarocorax atriceps and southern giant petrel Macronectes giganteus populations on the Antarctic Peninsula, 2001–2007. Mar. Ornithol. 36, 83–97 (2008).
Elliot, D. H., Watts, D. R., Alley, R. B. & Gracanin, T. M. Bird and seal observations at Joinville Island and offshore islands. Antarct. J. U. S. 13, 154–155 (1978).
Lynch, H. J. & Schwaller, M. R. Mapping the abundance and distribution of Adélie penguins using Landsat-7: first steps towards an integrated multi-sensor pipeline for tracking populations at the continental scale. PLoS ONE 9, e113301, https://doi.org/10.1371/journal.pone.0113301 (2014).
Nicol, S., Foster, J. & Kawaguchi, S. The fishery for Antarctic krill – recent developments. Fish Fish. 13, 30–40 (2012).
Woehler, E. J. The distribution and abundance of Antarctic and subantarctic penguins (Scientific Committee on Antarctic Research, 1993).
Polito, M. J., Lynch, H. J., Naveen, R. & Emslie, S. D. Stable isotopes reveal regional heterogeneity in the pre-breeding distribution and diets of sympatrically breeding Pygoscelis spp. penguins. Mar. Ecol. Prog. Ser. 421, 265–277 (2011).
Hinke, J. T. et al. Identifying risk: concurrent overlap of the Antarctic krill fishery with krill-dependent predators in the Scotia Sea. PLoS ONE 12, e0170132, https://doi.org/10.1371/journal.pone.00170132 (2017).
Reid, K. Sims, M., White, R. W. & Gillon K. W. Spatial distribution of predator/prey interactions in the Scotia Sea: implications for measuring predator/fisheries overlap. Deep Sea Res. Pt. II 51, 1383-1396 (2004).
Trivelpiece, W. Z. et al. Variability in krill biomass links harvesting and climate warming to penguin population changes in Antarctica. Proc. Natl. Acad. Sci. 108, 7625–7628 (2011).
Lynch, H. J., Naveen, R., Trathan, P. N. & Fagan, W. F. Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 93, 1367–1377 (2012).
Cimino, M. A., Lynch, H. J., Saba, V. S. & Oliver, M. J. Asymmetric response of Adélie penguins to Antarctic climate change. Sci. Rep. 6, 28785, https://doi.org/10.1038/srep28785 (2016).
Parkinson, C. L. Trends in the length of the Southern Ocean sea-ice season, 1979–99. Ann. Glaciol. 34, 435–440 (2002).
Parkinson, C. L. & Cavalieri, D. J. Antarctic sea ice variability and trends, 1979–2010. The Cryosphere 6, 871–880 (2012).
Laws, R. M. The ecology of the Southern Ocean. Amer. Sci. 73, 26–40 (1985).
Teschke, K., et al. Proposal for the establishment of a marine CCAMLR MPA in the Weddell Sea (Antarctica) – First conceptual outline. Report WG-EMM-13/22. (Commission for the Conservation of Antarctic Marine Living Resources, 2013).
Teschke, K. et al. Scientific background document in support of a CCAMLR MPA in the Weddell Sea (Antarctica) – Version2016 – Part A: General context of the establishment of MPAs and background information on the Weddell Sea MPA planning area. Report WG-EMM-16/01 (Commission for the Conservation of Antarctic Marine Living Resources, 2016).
Hamer, R. D. & Hyden, G. The geochemistry and age of the Danger Islands pluton, Antarctic Peninsula. Br. Antarct. Surv. Bull. 64, 1–19 (1984).
Ingólfsson, Ó. Glacial and climate history of the Antarctic Peninsula since the Last Glacial Maximum. Arct. Antarct. Alp. Res. 35, 175–186 (2003).
Emslie, S. D., McKenzie, A., Marti, L. J., & Santos, M. Recent occupation by Adelie penguins (Pygoscelis adeliae) at Hope Bay and Seymour Island and the ‘northern enigma’ in the Antarctic Peninsula. Polar Biol. https://doi.org/10.1007/s00300-017-2170-8 (2017).
Black, C. A. comprehensive review of the phenology of Pygoscelis penguins. Polar Biol. 39, 405–432 (2016).
Croxall, J. P. & Kirkwood, E. D. The distribution of penguins on the Antarctic Peninsula and islands of the Scotia Sea (British Antarctic Survey, 1979).
Naveen, R., Lynch, H. J., Forrest, S., Mueller, T. & Polito, M. First direct, site-wide penguin survey at Deception Island, Antarctica, suggests significant declines in breeding chinstrap penguins. Polar Biol. 35, 1879–1888 (2012).
Goebel, M. E. et al. A small unmanned aerial system for estimating abundance and size of Antarctic predators. Polar Biol. 35, 619–630 (2015).
Szegedy, C. et al. Going deeper with convolutions in Proceedings of the IEEE conference on computer vision and pattern recognition. 1–9 (Boston, 2015).
McDowall, P. & Lynch, H. J. Ultra-Fine Scale Spatially-Integrated Mapping of Habitat and Occupancy Using Structure-From-Motion. PLoS ONE 12, e0166773, https://doi.org/10.1371/journal.pone.0166773 (2017).
FIDASE - Falkland Islands and Dependencies Aerial Survey Expedition. ARX26FID0102127. 1:27,000. Roll 000102. Frame 127. Cambridge, U.K. (1957a).
FIDASE - Falkland Islands and Dependencies Aerial Survey Expedition. ARX26FID0102115. 1:27,000. Roll 000102. Frame 115. Cambridge, U.K. (1957b).
Quantum GIS Development Team. Quantum GIS geographic information system. Open Source Geospatial Foundation Project. http://qgis.osgeo.org (2016).
Schwaller, M. R., Southwell, C. J. & Emmerson, L. M. Continental-scale mapping of Adélie penguin colonies from Landsat imagery. Remote Sens. Environ. 139, 353–364 (2013).
Acknowledgements
We gratefully acknowledge the financial support of the Dalio Foundation, Inc. through the Dalio Explore Fund, which provided all the financing for the Danger Island Expedition. We are grateful to the crew of the yacht M/V Hans Hansson – Dion Poncet, Juliette Hannequinn, Alec and Gizelle Hazell for getting us to the Danger Islands and ensuring the safety and well-being of the research team during their work. We would like to thank additional support for analysis from the National Science Foundation (NSF PLR&GSS 1255058 - H.J.L. and P.M.; NSF PLR 1443585 – M.J.P.) and the National Aeronautical and Space Administration (NNX14AC32G; H.J.L. and M.S.). Geospatial support for the analysis of high resolution satellite imagery provided by the Polar Geospatial Center under NSF PLR awards 1043681 & 1559691.
Author information
Authors and Affiliations
Contributions
A.B. and H.J.L. wrote the main manuscript text and prepared the figures. S.J., P.M., C.Y., T.S.-M., T.H., M.J.P. and M.S. contributed to the manuscript. H.J.L., T.H., S.J., M.J.P., and H.S. conceived of and designed the project. S.J., M.J.P. and H.S. obtained funding for the expedition. A.B., P.M., C.Y., T.S.-M., G.C., R.H., S.F., M.R., T.H., and M.J.P. conducted field surveys. T.S.-M. and P.M. carried out the UAV imagery analysis. H.J.L. and M.S. provided satellite imagery analysis. A.B. conducted the historical aerial analysis. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Borowicz, A., McDowall, P., Youngflesh, C. et al. Multi-modal survey of Adélie penguin mega-colonies reveals the Danger Islands as a seabird hotspot. Sci Rep 8, 3926 (2018). https://doi.org/10.1038/s41598-018-22313-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-22313-w
This article is cited by
-
ShetlandsUAVmetry: unmanned aerial vehicle-based photogrammetric dataset for Antarctic environmental research
Scientific Data (2024)
-
Adélie penguins north and east of the ‘Adélie gap’ continue to thrive in the face of dramatic declines elsewhere in the Antarctic Peninsula region
Scientific Reports (2023)
-
Artificial intelligence for automated detection of large mammals creates path to upscale drone surveys
Scientific Reports (2023)
-
Quantifying Antarctic krill connectivity across the West Antarctic Peninsula and its role in large-scale Pygoscelis penguin population dynamics
Scientific Reports (2023)
-
Penguindex: a Living Planet Index for Pygoscelis species penguins identifies key eras of population change
Polar Biology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
