
Abstract
Membrane fouling still remains a major obstacle for wider applications of membrane bioreactor (MBR), which is mainly caused by soluble microbial products (SMP). Identification of key bacteria responsible for SMP production is essential for mitigation of membrane fouling. Here, we investigated the effect of microbial interaction on membrane fouling. We measured the membrane fouling potentials of 13 bacterial strains isolated from a pilot-scale MBR treating domestic wastewater when they were cultivated as single-culture and co-culture. We found that fouling-causing bacteria (FCB) displayed much higher fouling potential when co-cultured even with non-FCB and mixed population (activated sludge). In particular, the fouling potential of strain S26, one of FCB, increased 26.8 times when cultivated with strain S22 (fouling-enhancing bacteria, FEB). The secretion of N-octanoyl-L-homoserine lactone (C8-HSL) was increased by co-cultivating S22 and S26 as compared with cultivating as single culture, which stimulated the production of fouling-causing SMP by S26 and consequently resulted in severe membrane fouling. This result suggests that AHL-mediated quorum-sensing (QS) regulatory system was involved in secretion of fouling-causing SMP.
Similar content being viewed by others

Introduction
Membrane bioreactor (MBR) is considered to be a core wastewater reclamation technology to fulfill our diverse water demand1, 2. However, membrane fouling is still one of the main obstacles for its wider applications2, 3. Soluble microbial product (SMP) and extracellular polymeric substances (EPS) were considered to be main causes of membrane fouling in MBRs4. Therefore, better understanding of the detailed mechanisms of SMP and EPS production is essential for membrane fouling mitigation and control strategies5.
Extensive studies have been conducted to isolate and characterize key bacteria responsible for membrane fouling6,7,8,9. In our previous study, forty-one bacterial strains were isolated from a pilot-scale MBR treating municipal wastewater, tested for their fouling potential using a cross-flow filtration unit, and related to their cellular properties8. Thirteen out of 41 isolated strains displayed significant fouling potentials when they were cultivated as single-cultures, and thus were considered as fouling-causing bacteria (FCB). However, since bacteria were commonly present as mixed species in actual MBRs, the influence of microbial interaction of isolated bacterial strains on membrane fouling should be examined.
Several studies have revealed microbial interactions stimulated the formation of thicker biocake10, 11. The bacteria initially colonized on membrane surface and their SMP and EPS secretion facilitated subsequent bacterial attachment and growth, and eventually caused severe membrane fouling10, 11. However, it is not clear how microbial interaction influences the production of SMP and EPS and consequently membrane fouling.
It is well known that a variety of bacteria are capable of producing signal molecules (i.e. N-acyl homoserine lactones (AHL)), communicating each other and regulating gene expression in response to population density, which is known as AHL-mediated quorum-sensing (QS) system12. Several AHLs-producing bacteria were identified in MBRs13. Furthermore, several studies have revealed the links between the presence of various AHL signals with biocake formation and membrane fouling in MBRs14,15,16. Although the amounts of SMP and EPS in both biocake and mixed liquor were related to the increase in AHL concentrations15, 16, the exact role and mechanism of AHL-based QS in membrane fouling are not still clearly understood due to complex microbial interactions.
The aim of this study is, therefore, to investigate whether microbial interaction influences on the fouling potential. To achieve this goal, thirteen bacterial strains isolated in our previous study were tested for their fouling potential when cultivated as single-culture and co-culture. As a result, three strains exhibited significant fouling potential as single-cultures, thus they were defined as fouling-causing bacteria (FCB). Furthermore, co-culturing strain S26, one of the FCB, with strain S22 dramatically increased the fouling potential, which was caused by AHL-mediated SMP production by S26. The SMP produced in the co-culture of FCB and fouling-enhancing bacteria (FEB, S22 in this study) was further characterized.
Materials and Methods
Bacterial strains
Representative strains were selected from 17 operational taxonomic units (OTUs) composed of 41 bacterial strains isolated in our previous study8 (Table 1). However, since 4 strains could not be cultured on M9 medium, only 13 strains were used for the following studies. The degree of fouling potential of the isolated strains was categorized in three groups (high, I; moderate, II; low: III)8 (Table 1). All the strains were cultivated with M9 medium containing 48 mM Na2HPO4, 22 mM KH2PO4, 8.5 mM NaCl, 19 mM NH4Cl, 2 mM MgSO4, 100 μM CaCl2, 20 mM glucose, and trace element.
Measurement of membrane fouling potential
Membrane fouling potential of isolated strains was determined by dead-end filtration. Colonies of each individual strain grown on R2A agar plates was suspended in M9 medium, and OD600 of the bacterial suspension was adjusted to 1.0 with fresh M9 medium. For single-culture studies, 0.5 mL of the bacterial culture (OD600 = 1.0) were inoculated into 50 mL of M9 medium and incubated for 48 hr at 30 °C. For co-culture studies, two strain cultures (0.25 mL each, OD600 = 1.0) were inoculated into 50 mL of M9 medium and incubated for 48 hr at 30 °C. For mixed population (sludge) study, each culture (0.05 mL, OD600 = 1.0) of FCB and mixed liquor (0.45 mL, OD600 = 1.0) of MBR treating municipal wastewater was inoculated into 50 mL of M9 medium and incubated for 48 hr at 30 °C17. After incubation, OD600 values of all the bacterial cultures were adjusted to approximately 0.3 with fresh M9 medium to eliminate cell concentration.
The supernatant after centrifugation (at 4 °C, 6,000 × g for 15 min) of the bacterial culture was subjected to dead-end filtration test to evaluate the fouling potential. Five mL of the supernatant was transferred to a filtration unit (UHP-25K; Advantec Toyo; Tokyo, Japan) with a flat membrane filter (Pore size is 0.2 μm and membrane diameter is 25 mm, hydrophilic PTFE; Advantec Toyo; Tokyo, Japan), and filtered with continuous stirring under 50 kPa of ambient air. Thereafter, MilliQ water (approximately 10 mL) was added in the filtration unit and filtered again under the same pressure but without continuous stirring18. The permeate flow rate of MilliQ water was measured, and the membrane resistance of the fouled membrane was calculated as follows;
where, P is the pressure (Pa), A is the filtration area of membrane (m2), μ is the dynamic viscosity of MilliQ water (Pa · s), and Q is the permeate flow rate of MilliQ water (m3/s).
Effect of supernatant and AHL on fouling potential
To investigate the microbial interaction between fouling-causing bacteria (FCB) and fouling-enhancing bacteria (FEB), S26 was cultured with the supernatant of either S22 or S31, and its fouling potential was measured. For the cultivation of S26, M9 medium was made with the supernatant of bacterial culture (OD600 = 0.3) sterilized with syringe filter (0.2 μm, Mixed Cellulose Ester; Advantec Toyo; Tokyo, Japan) to check the effect of microbial interaction on fouling potential via bacterial secretion. S22 and S31 were also cultured with M9 medium made with the supernatant of S26 to confirm the effect of the supernatant of FCB.
In order to evaluate the effect of AHL on enhancement of fouling potential, S22 and S26 was cultivated with M9 medium containing 4.4 μM of N-octanoyl-L-homoserine lactone (C8-HSL). The bacterial cultivation and the fouling potential measurement were carried out as mentioned above.
Thin-layer chromatograph (TLC) assay for AHL
TLC assay was conducted to identify the AHL produced by isolated strains as described elsewhere in previous study with small modification19, 20. Bacterial cultures of S22 and S26 as single culture, and S22 and S26 as co-culture culture were prepared as described for measurement of membrane fouling potential. After 48 hr incubation at 30 °C, the bacterial cultures was centrifuged (4 °C, 6,000 × g, 15 min) and filtered with the sterilized syringe filter (0.45 μm, Mixed Cellulose Ester; Advantec Toyo; Tokyo, Japan) to remove bacterial cells. The supernatant (100 mL) was concentrated about 20 times by freeze-dry and AHL was extracted four times with 10 mL of ethyl acetate. The ethyl acetate was evaporated and the residual was dissolved with 10 μL of ethyl acetate. This solution and 5 μL of AHL standards were spotted on the C18 reversed-phase plate (TLC silica gel 60 RP-18 F254s; Merck Co.; Darmstadt, Germany) and the chromatogram was developed with methanol/water (60:40, v/v). The production of AHL was determined with Chromobacterium violaceum VIR24 (VIR24)21. After dried in air, the plate was overlaid with thin film of the mixture of LB broth containing 0.6% (w/v) and an overnight culture of VIR24 at the ratio of 1:1. After the incubation overnight at 30 °C, purple spot appeared in response to the presence of AHL on the plate. Based on the color development, Rf value of AHL standard and AHL produced by the strains were estimated as described elsewhere16. Each spot was analyzed by using ImageJ to quantify the concentration of AHL using commercial C8-HSL (Cayman Chemical Co.; MI, USA) as the standard22.
SMP extraction and characterization
In this study, SMP was defined as total organic carbon (TOC) in the supernatant after centrifugation of bacterial culture at 4 °C, 6,000 × g for 15 min. The supernatant was dialyzed to remove residual glucose as described elsewhere23. OD600 value of all the culture was adjusted to approximately 0.3 with fresh M9 medium to eliminate cell concentration effect before the dialysis. Carbohydrate and protein concentrations in dialyzed SMP were quantified with phenol-sulfonic acid method using glucose as the standard and Lowry method using BSA as the standard, respectively. In order to estimate the quantities of SMP trapped on membrane, carbohydrate and protein in SMP before and after the dead-end filtration was measured24.
SMP was also characterized by using fourier transform infrared (FTIR). The spectrum of the freeze-dried SMP was measured by a FTIR spectrometry (FT/IR-660 Plus; JASCO Co.; Tokyo, Japan). The operating range was from 4000 to 1000 cm−1 with a resolution of 2 cm−1, and transmittance spectra were generated25. Principle compartment analysis (PCA) was carried out with the normalized spectra26.
All the statistical analyses including PCA were carried out with R 3.0.2 (R Development Core Team; Vienna, Austria). P values less than 0.05 were considered statistically significant in all analyses.
Results
Fouling potential of isolated strains as single-culture
Fouling potential of 13 isolated strains cultivated as single-cultures was determined by dead-end filtration (Fig. 1). Strain S05, S26, and S32 showed significantly high fouling potential as compared with other strains. Therefore, these strains were considered as fouling-causing bacteria (FCB) in this study.

Fouling potential of 13 isolated strains (see Table 1) cultivated as single-cultures and virgin membrane. Since S05, S26, and S32 exhibited high fouling potential, they were regarded as fouling-causing bacteria (FCB) in this study. The error bars indicate the standard deviations of two independent experiments.
Fouling potential of isolated strains as co-culture
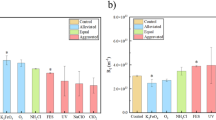
Fouling potential of 13 isolated strains was determined by dead-end filtration when they were cultivated as co-culture (Fig. 2A). There were 78 possible co-culture combinations of 13 isolated strains (the fouling potential and the OD600 value of all combinations were summarized in Tables S1 and S2, respectively). The fouling potential was elevated when all strains except for a few strains (S01, S12, and S14) was co-cultured with FCB (S05, S26, and S32). On the other hand, co-culturing with non-FCB did not significantly promote the fouling potential (Fig. 2A). These results suggest that membrane fouling was mainly caused by FCB, and some strains (S01, S12, and S14) might have capability to mitigate membrane fouling of FCB.

(A) Fouling potential of co-culture of 13 isolated strains (in total 78 combinations). (B) Effect of FCB (S05, S26, and S32) addition to activated sludge on fouling potential. The error bars indicate the standard deviations of two independent experiments. The bar following sign indicates statistical difference between two groups with respective p value: *P < 0.05. (C) Fouling potential of S26 as single-culture and co-culture with S22 or S31.
Furthermore, FCB were mixed with activated sludge of MBR treating municipal wastewater and cultured for 2 days at 30 °C, and then the fouling potential of culture medium was measured (Fig. 2B). The addition of all FCB significantly enhanced the fouling potential of activated sludge (p < 0.05), suggesting that FCB dominantly responsible for membrane fouling even in complex mixed populations.
Significant increase in fouling potential by co-culturing S22 and S26
When S26 was co-cultured with S22 or S31, the fouling potential increased to 1.1 × 1013 and 3.3 × 1012 (m−1), respectively, which were 26.8 or 7.8 times greater than the one of S26 single-culture (4.3 × 1011 (m−1)) (Table S1, Fig. 2C). The growth of S26 was similar regardless of single and co-cultures (Fig. S1). Furthermore, the fouling potential of S26 was significantly increased when S26 was cultured in M9 medium made with the filter-sterilized supernatant of S22 culture medium (9.6 times, p < 0.05) (Fig. 3). On the other hand, the fouling potential of S22 and S31 did not change when S22 and S31 were cultured in the M9 medium made with the filter-sterilized supernatant of S26 culture medium (Fig. 3). In addition, the fouling potential of the supernatant S26 did not change when mixed with the supernatant of S22 or S31, indicating that abiotic interaction such as aggregation did not affect the membrane fouling (Fig. 3). These results suggest that production and secretion of fouling-causing matter, probably soluble microbial product (SMP), by S26 was enhanced by addition of the filter-sterilized supernatant of S22 or S31 culture medium. Thus, S22 and S31 were considered as fouling-enhancing bacteria (FEB) in this manuscript.

Effect of addition of the supernatants on fouling potential. The filter-sterilized supernatant of bacterial culture was added to M9 medium for cultivation of other strains. For example, S26 + (S22) indicates that S26 was cultured in the M9 medium with the filter-sterilized supernatant of S22 culture. The mixture of supernatants was also tested for fouling potential to assess abiotic effects by co-cultivating. The error bars indicate the standard deviations of two replicates except for the mixture of supernatants. The bar following sign indicates statistical difference between two groups with respective p value: *P < 0.05.
The strain S22 was affiliated with the Genus Thermomonas and shared 98.5% of 16 S rRNA gene sequence with Thermomonas fusca DSM 1542427 (Table 1). The S26 was affiliated with the Genus Mesorhizobium and shared 98.4% of 16 S rRNA gene sequence with Mesorhizobium ciceri biovar biserrulae WSM127128 (Table 1). The S31 was closely related to Bacillus subtilis (100%), which is a gram negative bacteria. Mesorhizobium is known to produce AHL29. Although the ability of Thermomonas to produce AHL is not reported in previous studies, the genus Stenotrophomonas, which is also affiliated with the Family Xanthomonadaceae like Thermomonas, is known to produce AHL30. In addition, Mesorhizobium is known to possess LuxR/LuxI-type quorum-sensing regulatory system31, 32. Based on these evidences, it is speculated that the increased fouling potential of S26 is related to AHL-mediated processes.
TLC assay was performed to determine the production of AHL by S22 and S26. As a result, it was confirmed that both S22 and S26 produced most likely N-octanoyl-L-homoserine lactone (C8-HSL), but S26 produced more significantly than S22 (Fig. 4A and Fig. S2). In contrast, the productions of any AHL by the other FCB (i.e., S05 and S32) and S31 were not confirmed (data not shown). The C8-HSL production by S26 was increased by 78% when co-cultured with S22 (Fig. 4B). Furthermore, the fouling potential of S26 was significantly enhanced when S26 was co-cultured with S22 and cultured in M9 medium containing 4.4 μM C8-HSL for 2 day at 30 °C (p < 0.01) (Fig. 4C). In contrast, addition of C8-HSL did not increase the fouling potential of S22 (Fig. 4C). These results indicated that the increased production of C8-HSL during co-culturing S26 with S22 may have caused the increased fouling potential of S26.

(A) Thin-layer chromatograph assay of N-acyl-homoserine-lactone (AHL) produced by S22 and S26 (Full-size images were shown in Fig. S2). (B) N-octanoyl-L-homoserine lactone (C8-HSL) concentration produced by single-culture of S22, S26, and co-culture of S22 and S26. (C) Effect of C8-HSL addition on the fouling potential of S22 and S26 culture. The error bars indicate the standard deviations of two independent experiments. The bar following sign indicates statistical difference between two groups with respective p value: **P < 0.01.
Characterization of SMP
The carbohydrate content in SMP produced by S26 was higher than that of S22 (Fig. 5A), which might reflect the difference of fouling potential (Fig. 4C). Although total amount of SMP (carbohydrate and protein) did not change when S26 was cultured with S22 and C8-HSL (Fig. 5A), the membrane fouling potential was significantly increased (Fig. 4C). However, amount of SMP trapped on the membrane surface after filtration was significant different. The more SMP was detected on the membrane surface when strain S26 was cultured with S22 and C8-HSL (Fig. 5B), which consisted with the increase in the fouling potential (Fig. 4C). In contrast, no accumulation of SMP was found for the S22 culture medium. These results might indicate that the composition or characteristics, not total quantity, of SMP was an important factor determining membrane fouling and changed through C8-HSL produced by co-culturing with S22.

(A) Carbohydrate and protein concentrations in soluble microbial product (SMP) produced by S22, S26, S22 + S26 (co-culture) and S26 cultured with C8-HSL (4.4 μM). (B) Carbohydrate and protein contents in SMP trapped on membrane surface in the dead-end filtration experiment for these cultures. The error bars indicate the standard deviations of two independent experiments. The bar following sign indicates statistical difference between two groups with respective p value: *P < 0.05. (C) Principal compartment analysis based on fourier transform infrared (FTIR) spectrum of SMP. “Mixture” indicates the mixture of the SMP produced by strain S22 and S26 with a ratio 1:1. The SMP produced by co-culture was close to the SMP produced by S26, indicating that the co-culture SMP was mostly produced by strain S26.
The composition of SMP was characterized with PCA based on the spectrum gained by a FTIR spectrometry (Fig. 5C; the FTIR spectra were shown in Fig. S3). The FTIR spectrum of SMP in the co-culture of S26 and S22 was different from that in the single-culture of S22 and S26. For example, peaks around 3300 cm−1 and 1080 cm−1 in the co-culture or S26 + C8-HSL were bigger than in both single-cultures. There were more peaks in the range of 2500–1500 cm−1 in the co-culture than both single-cultures. The peaks around 3300 and 1080 cm−1 were designated as the presence of carbohydrate-like, while the peaks around 1650 cm−1 were designated as protein-like substances17, 25, 33. The SMPs produced by a single-culture S22 and S26 were plotted apart from each other, indicating different compositions. The mixture of the supernatants of single-culture S22 and S26 (1:1) was located in the middle of both plots (Fig. 5C). However, the plot representing co-culture of S22 and S26 was located near the plot representing S26 rather than S22, suggesting the co-culture SMP was resemble the SMP produced by S26 (Fig. 5C). Moreover, the SMP produced by S26 with exogenous C8-HSL was close to the plot representing co-culture of S22 and S26, suggesting that the composition of SMP produced by the co-culture was more similar to that by S26 under the presence of exogenous C8-HSL.
Discussion
In our previous study, the fouling potential of 41 phylogenetically distant strains that were previously isolated from the fouled membranes of a pilot-scale MBR treating real domestic wastewater were evaluated as single-culture8. However, the effect of co-culturing these isolates on membrane fouling was unknown presently; therefore it was investigated in the present study. It was found that FCB induced severe membrane fouling even in co-cultures with non-FCB and complex microbial community (activated sludge) (Fig. 2). In particular, the fouling potential of S26, one of the FCB, was increased 26.8 times when cultivated with S22 that stimulated the production of fouling-causing SMP by S26. On the other hand, any co-culture of non-FCB did not show higher fouling potential than those of single FCB cultures (Table S1). It is, therefore, speculated that FCB could be responsible for membrane fouling in the pilot-scale MBRs treating domestic wastewater, suggesting that membrane fouling potential of single-cultures of isolated strains provided useful, but limited, information.
The microbial interaction that stimulated fouling-causing SMP production by S26 and consequently caused severe membrane fouling was further investigated in detail. The production of SMP was considered to be highly dependent on microbial populations in MBR34. However, our findings demonstrated the secretion of C8-HSL molecule was significantly increased by co-culturing S26 and S22, which induced S26 to produce fouling-causing SMP (change the SMP composition), leading to severe membrane fouling (Fig. 5). Therefore, the concentration of C8-HSL was one of key factors to determine the fouling potential of co-culture of S26 and S22. When S26 was co-cultured with S31, the fouling potential was 7.8 times greater than the one of S26 single-culture. However, S31 was closely related to a gram negative bacterium, Bacillus subtilis (100%), which did not produce AHL, suggesting other regulatory mechanism could be involved in the increased fouling potential of S26.
Although many studies have examined the role of AHL-mediated QS in pure culture systems so far, available reports for co-culture or defined population systems are very limited. Given extensive studies so far, it is now conceivable that AHL-mediated QS system is important for biofilm formation. It is also known that EPS synthesis is subject to AHL molecules in both qualitative and quantitative manner35,36,37. It was reported that the AHL-mediated QS was involved in production and secretion of hydrophobic extracellular proteins, which promote microbial aggregation of activated sludge38. The carbohydrate and protein contents in EPS were also regulated by AHL38, 39. In MBRs, it was reported that the amounts of SMP and EPS in both biocake and mixed liquor were closely related to the increase in AHL concentration16. However, the exact mechanism of AHL in membrane fouling was not clearly revealed in these studies. To the best of our knowledge, the present study showed for the first time that AHL concentration was elevated by co-culturing FCB and FEB, which stimulated the production of fouling-causing SMP (induced the composition change) by FCB and consequently resulted in enhancement of membrane fouling.
Mesorhizobium and Thermomonas have been frequently detected in MBRs40,41,42. They were also found in both mixed liquor (approximately 0.004% and 1.6% of total reads analyzed by next-generation sequencing, respectively) and gel layer on fouled membrane (0.01% and 2.9% of the total reads, respectively) in the MBR where the bacterial strains were isolated8. Mesorhizobium is capable of producing AHL31. M. tianshanense possess MtrI and MtrR, which are homolog of LuxR and LuxI family proteins and act as a QS regulatory system32. In this genus, QS system has been linked to the regulation of nodulation efficiency, growth rate, and exopolysaccharide production, and nitrogen fixation43, 44. In contrast, there is limited information on the AHL-mediated QS of Thermomonas although it was reported that the growth was likely promoted by AHL45.
In addition to AHL-mediated QS systems, existence of quorum-quenching (QQ) systems that block the QS systems in MBRs has been reported15, 45, 46. Therefore, complex microbial interactions among QS bacteria, QQ bacteria, FCB, and FEB must be studied in the future for better understanding of membrane fouling in MBRs. In addition, possible combinations of QQ and other approaches such as chlorination47, enrichment of fouling-reducing bacteria (i.e. Bacillus and Chloroflexi)48, 49, and anodic respiration24 should be investigated for efficient membrane fouling mitigation.
Since microfiltration (MF) membrane with pore size of 0.2 μm was used in this study, a portion of SMP and AHLs might pass through the membrane instead of rejected by and remained on the membrane. Therefore, the membrane fouling potentials measured in this study would not represent correctly the fouling potentials of the isolated bacterial strains.
In conclusion, the effect of co-culturing bacterial strains isolated from a pilot-scale MBR on membrane fouling were investigated in the present study. It was found that FCB, especially S26 (closely related to Mesorhizobium ciceri (98.4%)), induced severe membrane fouling when co-cultured with non-FCB and activated sludge, suggesting that FCB were mainly responsible for membrane fouling in the pilot-scale MBRs treating domestic wastewater. In particular, the fouling potential of S26 was increased 26.8 times when cultivated with S22 (closely related to Thermomonas fusca (98.5%)). The mechanism enhancing membrane fouling in this co-culture was further investigated in detail. As a result, co-culturing S22 and S26 induced the increase in C8-HSL production and thereby stimulated the production of fouling-causing SMP (induced the composition change) by S26, which consequently resulted in enhancement of membrane fouling. Taken together, AHL-mediated QS system was involved in SMP production and membrane fouling, which could not be revealed by single-culture studies.
References
Judd, S. The status of membrane bioreactor technology. Trends Biotechnol. 26, 109–116 (2008).
Meng, F. et al. Recent advances in membrane bioreactors (MBRs): membrane fouling and membrane material. Water Res. 43, 1489–1512 (2009).
Zuthi, M. F. R., Guo, W., Ngo, H., Nghiem, L. & Hai, F. Enhanced biological phosphorus removal and its modeling for the activated sludge and membrane bioreactor processes. Bioresour. Technol. 139, 363–374 (2013).
Tran, N. H., Ngo, H. H., Urase, T. & Gin, K. Y. H. A critical review on characterization strategies of organic matter for wastewater and water treatment processes. Bioresour. Technol. 193, 523–533 (2015).
Malaeb, L., Le-Clech, P., Vrouwenvelder, J. S., Ayoub, G. M. & Saikaly, P. E. Do biological-based strategies hold promise to biofouling control in MBRs? Water Res. 47, 5447–5463 (2013).
Choi, H., Zhang, K., Dionysiou, D. D., Oerther, D. B. & Sorial, G. A. Effect of activated sludge properties and membrane operation conditions on fouling characteristics in membrane bioreactors. Chemosphere 63, 1699–1708 (2006).
Pang, C. M., Hong, P., Guo, H. & Liu, W. T. Biofilm formation characteristics of bacterial isolates retrieved from a reverse osmosis membrane. Environ. Sci. Technol. 39, 7541–7550 (2005).
Ishizaki, S., Fukushima, T., Ishii, S. & Okabe, S. Membrane fouling potentials and cellular properties of bacteria isolated from fouled membranes in a MBR treating municipal wastewater. Water Res. 100, 448–457 (2016).
Juang, Y. C., Adav, S. S. & Lee, D. J. Strains of internal biofilm in aerobic granular membrane bioreactors. Appl. Microbiol. Biotechnol. 86, 1987–1993 (2010).
Gao, D. W., Wen, Z. D., Li, B. & Liang, H. Membrane fouling related to microbial community and extracellular polymeric substances at different temperatures. Bioresour. Technol. 143, 172–177 (2013).
Vanysacker, L., Declerck, P., Bilad, M. & Vankelecom, I. Biofouling on microfiltration membranes in MBRs: Role of membrane type and microbial community. J. Membrane Sci. 453, 394–401 (2014).
Fuqua, W. C., Winans, S. C. & Greenberg, E. P. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J. bacteriol. 176, 269 (1994).
Lade, H., Paul, D. & Kweon, J. H. Isolation and molecular characterization of biofouling bacteria and profiling of quorum sensing signal molecules from membrane bioreactor activated sludge. Int. J. Biol. Sci. 15, 2255–2273 (2014).
Huang, J. et al. Acyl-homoserine lactone-based quorum sensing and quorum quenching hold promise to determine the performance of biological wastewater treatments: An overview. Chemosphere 157, 137–151 (2016).
Yu, H., Xu, G., Qu, F., Li, G. & Liang, H. Effect of solid retention time on membrane fouling in membrane bioreactor: from the perspective of quorum sensing and quorum quenching. Appl. Microbiol. Biotechnol. 100, 7887–7897 (2016).
Yeon, K. M. et al. Quorum sensing: a new biofouling control paradigm in a membrane bioreactor for advanced wastewater treatment. Environ. Sci. Technol. 43, 380–385 (2008).
Kimura, K., Yamato, N., Yamamura, H. & Watanabe, Y. Membrane fouling in pilot-scale membrane bioreactors (MBRs) treating municipal wastewater. Environ. Sci. Technol. 39, 6293–6299 (2005).
Kimura, K. et al. Further examination of polysaccharides causing membrane fouling in membrane bioreactors (MBRs): Application of lectin affinity chromatography and MALDI-TOF/MS. Water Res. 46, 5725–5734 (2012).
Shaw, P. D. et al. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. 94, 6036–6041 (1997).
McClean, K. H. et al. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143, 3703–3711 (1997).
Someya, N. et al. Distribution of N-acylhomoserine lactone-producing fluorescent pseudomonads in the phyllosphere and rhizosphere of potato (Solanum tuberosum L.). Microbes Environment 24, 305–314 (2009).
Kanfi, Y. et al. Regulation of SIRT1 protein levels by nutrient availability. FEBS lett. 582, 2417–2423 (2008).
Miura, T. et al. Histo-blood group antigen-like substances of human enteric bacteria as specific adsorbents for human noroviruses. J. Virol. 87, 9441–9451 (2013).
Ishizaki, S., Terada, K., Miyake, H. & Okabe, S. Impact of anodic respiration on biopolymer production and consequent membrane fouling. Environ. Sci. Technol. 50, 9515–9523 (2016).
Hou, X., Liu, S. & Zhang, Z. Role of extracellular polymeric substance in determining the high aggregation ability of anammox sludge. Water res. 75, 51–62 (2015).
Lammers, K., Arbuckle-Keil, G. & Dighton, J. FT-IR study of the changes in carbohydrate chemistry of three New Jersey pine barrens leaf litters during simulated control burning. Soil Biol. Biochem. 41, 340–347 (2009).
Mergaert, J., Cnockaert, M. C. & Swings, J. Thermomonas fusca sp. nov. and Thermomonas brevis sp. nov., two mesophilic species isolated from a denitrification reactor with poly (ε-caprolactone) plastic granules as fixed bed, and emended description of the genus Thermomonas. Int. J. Syst. Evol. Microbiol. 53, 1961–1966 (2003).
Nandasena, K. G., O’Hara, G. W., Tiwari, R. P., Willlems, A. & Howieson, J. G. Mesorhizobium ciceri biovar biserrulae, a novel biovar nodulating the pasture legume Biserrula pelecinus L. Int. J. Syst. Evol. Microbiol. 57, 1041–1045 (2007).
Lade, H., Paul, D. & Kweon, J. H. Quorum quenching mediated approaches for control of membrane biofouling. Int. J. Biol. Sci. 10, 550–565 (2014).
Valle, A., Bailey, M. J., Whiteley, A. S. & Manefield, M. N-acyl-L-homoserine lactones (AHLs) affect microbial community composition and function in activated sludge. Environ. Microbiol. 6, 424–433 (2004).
Zheng, H. et al. A LuxR/LuxI-type quorum-sensing system in a plant bacterium, Mesorhizobium tianshanense, controls symbiotic nodulation. J. Bacteriol. 188, 1943–1949 (2006).
Ramsay, J. P. et al. A LuxRI-family regulatory system controls excision and transfer of the Mesorhizobium loti strain R7A symbiosis island by activating expression of two conserved hypothetical genes. Mol. Microbiol. 73, 1141–1155 (2009).
Yamamura, H., Okimoto, K., Kimura, K. & Watanabe, Y. Hydrophilic fraction of natural organic matter causing irreversible fouling of microfiltration and ultrafiltration membranes. Water res. 54, 123–136 (2014).
Guo, W., Ngo, H. H. & Li, J. A mini-review on membrane fouling. Bioresour. Technol. 122, 27–34 (2012).
Marketon, M. M., Glenn, S. A., Eberhard, A. & González, J. E. Quorum sensing controls exopolysaccharide production in Sinorhizobium meliloti. J. Bacteriol. 185, 325–331 (2003).
Rinaudi, L. V. & González, J. E. The low-molecular-weight fraction of exopolysaccharide II from Sinorhizobium meliloti is a crucial determinant of biofilm formation. J. Bacteriol. 191, 7216–7224 (2009).
Sakuragi, Y. & Kolter, R. Quorum-sensing regulation of the biofilm matrix genes (pel) of Pseudomonas aeruginosa. J. Bacteriol. 189, 5383–5386 (2007).
Lv, J. et al. The effect of quorum sensing and extracellular proteins on the microbial attachment of aerobic granular activated sludge. Bioresour. Technol. 152, 53–58 (2014).
Tan, C. H. et al. The role of quorum sensing signalling in EPS production and the assembly of a sludge community into aerobic granules. ISME J. 8, 1186–1197 (2014).
Shen, L., Yao, Y. & Meng, F. Reactor performance and microbial ecology of a nitritation membrane bioreactor. J. Membrane Sci. 462, 139–146 (2014).
Silva, A. F. et al. Impact of sludge retention time on the fine composition of the microbial community and extracellular polymeric substances in a membrane bioreactor. Appl. Microbiol. Biotechnol. 100, 8507–8521 (2016).
Tian, H. L. et al. Bacterial community shift along with the changes in operational conditions in a membrane-aerated biofilm reactor. Appl. Microbiol. Biotechnol. 99, 3279–3290 (2015).
Yang, M. et al. Functional analysis of three AHL autoinducer synthase genes in Mesorhizobium loti reveals the important role of quorum sensing in symbiotic nodulation. Can. J. Microbiol. 55, 210–214 (2009).
Cao, H. et al. Complex quorum-sensing regulatory systems regulate bacterial growth and symbiotic nodulation in Mesorhizobium tianshanense. Arch. Microbiol. 191, 283–289 (2009).
Hu, J., Li, H., Li, H., Liuacd, Y. & Song, S. Role of N-acyl-homoserine lactone (AHL) based quorum sensing on biofilm formation on packing media in wastewater treatment process. RSC Adv. 6, 11128–11139 (2016).
Oh, H. S. et al. Control of membrane biofouling in MBR for wastewater treatment by quorum quenching bacteria encapsulated in microporous membrane. Environ. Sci. Technol. 46, 4877–4884 (2012).
Weerasekara, N. A., Choo, K. H. & Lee, C. H. Biofouling control: Bacterial quorum quenching versus chlorination in membrane bioreactors. Water Res. 103, 293–301 (2016).
Miura, Y., Watanabe, Y. & Okabe, S. Significance of Chloroflexi in performance of submerged membrane bioreactors (MBR) treating municipal wastewater. Environ. Sci. Technol. 41, 7787–7794 (2007).
Zhang, Y. et al. Antibiofilm activity of Bacillus pumilus SW9 against initial biofouling on microfiltration membranes. Appl. Microbiol. Biotechnol. 98, 1309–1320 (2014).
Acknowledgements
This research was partly supported by Sumitomo Foundation, Japan, which was granted to Satoshi Okabe and by Grant-in Aid for JSPS Fellows (2600217604) of the Japan Society for the Promotion of Science (JSPS). The authors would like to thank to Hiroshi Miyake, Lei Zhang, Tomoyo Kageyama, and Tatsuma Takagi for technical assistance.
Author information
Authors and Affiliations
Contributions
S.I. and S.O. designed research; S.I. and R.S. performed research; S.I. and S.O. analyzed data; S.I. and S.O. wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ishizaki, S., Sugiyama, R. & Okabe, S. Membrane fouling induced by AHL-mediated soluble microbial product (SMP) formation by fouling-causing bacteria co-cultured with fouling-enhancing bacteria. Sci Rep 7, 8482 (2017). https://doi.org/10.1038/s41598-017-09023-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-09023-5
This article is cited by
-
Advanced wastewater treatment and membrane fouling control by electro-encapsulated self-forming dynamic membrane bioreactor
npj Clean Water (2022)
-
The biological role of N-acyl-homoserine lactone-based quorum sensing (QS) in EPS production and microbial community assembly during anaerobic granulation process
Scientific Reports (2018)
-
Fouling Development in A/O-MBR under Low Organic Loading Condition and Identification of Key Bacteria for Biofilm Formations
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
