
Abstract
SR-BI is the main receptor for high density lipoproteins (HDL) and mediates the bidirectional transport of lipids, such as cholesterol and vitamin E, between these particles and cells. During early development, SR-BI is expressed in extraembryonic tissue, specifically in trophoblast giant cells in the parietal yolk sac. We previously showed that approximately 50% of SR-BI−/− embryos fail to close the anterior neural tube and develop exencephaly, a perinatal lethal condition. Here, we evaluated the role of SR-BI in embryonic vitamin E uptake during murine neural tube closure. Our results showed that SR-BI−/− embryos had a very low vitamin E content in comparison to SR-BI+/+ embryos. Whereas SR-BI−/− embryos with closed neural tubes (nSR-BI−/−) had high levels of reactive oxygen species (ROS), intermediate ROS levels between SR-BI+/+ and nSR-BI−/− embryos were detected in SR-BI−/− with NTD (NTD SR-BI−/−). Reduced expression of Pax3, Alx1 and Alx3 genes was found in NTD SR-BI−/− embryos. Maternal α-tocopherol dietary supplementation prevented NTD almost completely (from 54% to 2%, p < 0.001) in SR-BI−/− embryos and normalized ROS and gene expression levels. In sum, our results suggest the involvement of SR-BI in the maternal provision of embryonic vitamin E to the mouse embryo during neural tube closure.
Similar content being viewed by others
Introduction
Scavenger Receptor Class B type I (SR-BI) is the main receptor for high density lipoproteins (HDL), and numerous studies have described its role in mediating the bidirectional transport of lipids between these lipoproteins and cells1. In the liver, SR-BI is involved in the uptake of cholesterol from HDL and its excretion in bile, the final step in reverse cholesterol transport. SR-BI also participates in the uptake of cholesterol in steroidogenic tissues, such as the adrenal glands and ovaries, to be used as a substrate for steroid hormone synthesis2. Important information on the roles of SR-BI other than in cholesterol homeostasis and cholesterol provision for steroidogenesis, such as platelet aggregation, erythrocyte maturation and oocyte meiosis, has been generated from the SR-BI knock out (SR-BI−/−) mouse since it was generated almost two decades ago3.
In generating SR-BI−/− mice via heterozygous intercrosses, researchers noted that the proportion of weaned homozygous null mice was half that expected by the Mendelian ratio3. This evidence, together with the fact that SR-BI is present in murine trophoblasts involved in maternal-foetal nutrient exchange at different stages of gestation4, led researchers to postulate that this HDL receptor might be involved in embryonic development. We recently showed that nearly 50% of SR-BI−/− embryos fail to close the anterior neural tube and develop cranial NTD and exencephaly5, leading to perinatal death, which explains the deviation from the Mendelian ratio previously reported in weaned SR-BI null mice3. Among the spectrum of defective neurulation conditions conferred by abnormal closure at different portions of the neural tube, only cranial NTD is observed in SR-BI−/− embryos.
During murine early development, SR-BI is not detected in the embryo itself but rather in trophoblast giant cells (TGC) from the parietal yolk sac4, 5. TGC play a critical role in embryonic uptake of various nutrients from the maternal blood supply before the establishment of a mature placenta6. Despite the prominent role of SR-BI in cholesterol uptake1 and the key role of cholesterol during embryonic development7, we have shown that the embryonic cholesterol content is similar in SR-BI+/+ and SR-BI−/− embryos5.
In addition to cholesterol, HDL particles also transport other lipids, such as triglycerides, lipophilic vitamins and hydrophobic signalling molecules. Studies using adult SR-BI−/− mice demonstrated lower levels of vitamin E in several SR-BI-expressing tissues, suggesting that beyond its role as a cholesterol transporter, this receptor also mediates the cellular uptake of vitamin E from HDL8. Vitamin E is a generic name for a group of isomers of two related molecules: tocopherols and tocotrienols. This vitamin was first described almost 100 years ago to be an essential factor for the success of pregnancy in rats9. Vitamin E isomers have antioxidant activity and intercalate between lipids in biological membranes, where they stop reactive oxygen species (ROS)-based reactions that generate lipoperoxides10. In addition, some isomers have other less well-defined biological activities, such as modulation of intracellular signalling pathways, gene expression and cell proliferation11. Vitamin E deficiency in rodents has been associated with congenital malformations, including neural tube defects (NTD)12. Consistent with this finding, this vitamin is effective in preventing NTD in mouse models of maternal diabetes-induced malformations, which are associated with an increased oxidative status13.
Considering this evidence, we hypothesized that NTD in SR-BI−/− embryos was due to oxidative stress resulting from impaired vitamin E uptake from maternal circulation. To test this hypothesis, we studied the incidence of NTD, vitamin E content, ROS levels and gene expression in embryos and in TGC retrieved from dams fed with control or α-tocopherol-enriched diets.
Results
Effect of maternal α-tocopherol supplementation on NTD in SR-BI−/− embryos
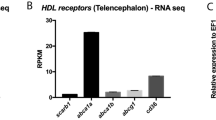
As a first approach to determine if defective maternal-embryonic vitamin E metabolism contributes to neural malformation in SR-BI−/− embryos, we sought to analyse whether α-tocopherol supplementation in pregnant SR-BI+/− dams reduced the incidence of NTD. The rationale behind this hypothesis was that if SR-BI is involved in vitamin E transport across the maternal foetal interface, then increasing the concentration of this vitamin in the maternal plasma may compensate for the lack of SR-BI via SR-BI-independent transport pathways. This possibility was strongly supported by our analysis of transcriptomic data in parietal TGC generated by Hannibal et al.14 which showed that, besides SR-BI, TGC express other proteins that are involved in lipoprotein metabolism, such as the HDL receptor Gpihbp1, receptors for others lipoproteins (e.g., Ldlr, Apobr, and Lrp1), and the lipid transporters Abca1 and Abcg1, which mediate lipid efflux from cells to Apoa1 and HDL, respectively (Table 1).
Because vitamin E supplementation has been shown to reduce plasma cholesterol levels in rodent models of hypercholesterolemia15,16,17 and because cholesterol has been shown to be crucial for embryonic development7, we analysed the impact of α-tocopherol supplementation on the plasma cholesterol levels of pregnant dams. Consumption of the vitamin-enriched diet did not affect the total circulating cholesterol levels nor its distribution into lipoproteins, as both parameters were similar in dams fed control or supplemented diets (Supplementary Fig. 1a and b). Alpha-tocopherol dietary supplementation from conception until gestational day 9.5 (E9.5) resulted in a significant rise in this lipid in maternal plasma (Fig. 1a). Interestingly, the vitamin E distribution in lipoproteins was affected by α-tocopherol supplementation. Compared to plasma from control mice, in which almost no α-tocopherol was detected in larger lipoproteins such as LDL, VLDL or chylomicrons, plasma from dams fed the vitamin E-enriched diet exhibited similar levels of α-tocopherol in HDL and non-HDL particles (Fig. 1b). In sum, the expression of other lipoprotein receptors than SR-BI in TGC (Table 1) and the detection of α-tocopherol in non-HDL lipoproteins in SR-BI+/− dams fed the vitamin E-supplemented diet (Fig. 1b) suggest the co-existence of different α-tocopherol transport mechanisms from maternal blood into TGC, involving both HDL and non-HDL lipoproteins.

Plasmatic total (a) and lipoprotein (b) α-tocopherol levels in SR-BI+/− pregnant dams fed with control or vitamin E supplemented diets. Concentrations of α-tocopherol in total plasma (a) or in different lipoproteins (b). VLDL/ChM: fraction containing very low density lipoproteins and chylomicrons. LDL: low density lipoproteins. HDL: high density lipoproteins. The horizontal lines indicate median (a) and mean (b). *p < 0.05, assessed by Mann-Whitney (a) or two-way ANOVA with Holms-Sidak post-test (b).
To analyse the impact of α-tocopherol supplementation on the incidence of NTD, embryos at E9.5 were retrieved from chow- or vitamin E-fed heterozygous dams and phenotypically classified as normal or as having NTD. Consistent with previous results from our group, a significant proportion of SR-BI−/− embryos (54%) and a few SR-BI−/+ embryos (6%) from chow-fed SR-BI−/+ dams exhibited cranial NTD (Table 2). By contrast, only 1 out of 59 SR-BI−/− embryos and 2 out of 129 SR-BI−/+ embryos (less than 2% in both cases) showed cranial NTD in vitamin E-supplemented SR-BI−/+ pregnancies (Table 2). Interestingly, α-tocopherol supplementation after implantation, starting from E4.5, also reduced the proportion of SR-BI−/− and SR-BI−/+ embryos with NTD to levels comparable with supplementation from E0.5 to E9.5 (Table 3). Injecting pregnant dams with folic acid, a water-soluble vitamin that is well known to prevent NTD, reduced the percentage of SR-BI−/− embryos with NTD from 56% to 19%, showing that NTD is folate-sensitive in SR-BI−/− embryos (Table 4). The mean number of viable embryos that showed cardiac activity and the mean number of resorptions retrieved per dam were similar in the control, vitamin E- and folate-supplemented groups, indicating that embryonic viability and female fertility were not affected by these maternal interventions (Supplementary Table 2). As in our previous work, only cranial NTD, and not defective closure in the dorsal or posterior neural tube, was observed in SR-BI−/− and SR-BI−/+ embryos, irrespective of treatment. In summary, α-tocopherol dietary supplementation, starting either after mating or at implantation, proved to be a highly efficient intervention to prevent NTD in SR-BI-deficient embryos.
Vitamin E content in embryos and parietal yolk sacs lacking SR-BI
We next sought to determine whether the lack of SR-BI affected the vitamin E content in SR-BI−/− mouse embryos and TGC, as previously reported for certain adult tissues8. Supporting the notion that SR-BI is involved in vitamin E maternal-embryonic provision, α-tocopherol quantification in SR-BI−/− embryos showed a 50-fold reduction compared to SR-BI+/+ embryos (Fig. 2a). Two unexpected results were obtained from these experiments: i) the very low vitamin E content was similar in SR-BI−/− embryos with NTD (SR-BI−/− NTD) or with normal neural tube closure (nSR-BI−/−) and ii) maternal supplementation with α-tocopherol did not increase the embryonic vitamin E levels, despite its striking effect on NTD prevention (Fig. 2a).

Vitamin E content in embryos (a) and parietal yolk sac (PYS) (b) obtained from SR-BI+/− dams fed with control or vitamin E supplemented diets. Levels of α-tocopherol were measured in pairs of wild-type embryos (SR-BI+/+), normal knock-out embryos (nSR-BI−/−) and knock-out embryos with NTD (SR-BI−/− NTD) (a) and in single PYS (b). **p ≤ 0.01, ***p ≤ 0.001, determined by one-way ANOVA and Tukey’s post-test.
SR-BI is specifically expressed in TGC, but these large cells are tightly adhered to small parietal endoderm cells by a thick extracellular matrix constituting the parietal yolk sac (PYS). In order to avoid potential modifications in the metabolic or transcriptional status of TGCs due to isolation procedures, we determined the content of α-tocopherol in the PYS as a whole. The α-tocopherol content was similar in PYS from SR-BI+/+ and SR-BI−/− embryos with or without NTD obtained from chow-fed dams (Fig. 2b). Although maternal supplementation with α-tocopherol did not increase the vitamin E content in SR-BI+/+ PYS, the vitamin E content was almost 5-fold higher in SR-BI−/− PYS.
Redox status in SR-BI−/− embryos and TGC
Because accumulation of oxidative species has been shown to impair neural tube closure in mice13, we determined the reactive oxygen species (ROS) levels by fluorimetry in vitamin E-deficient SR-BI−/− embryos (see methods and Supplementary Fig. 2). In chow-fed dams, nSR-BI−/− embryos exhibited a 5-fold higher fluorescence intensity compared to SR-BI+/+ embryos (Fig. 3a). Surprisingly, NTD SR-BI−/− embryos showed intermediate fluorescence levels compared to SR-BI+/+ and nSR-BI−/−. Maternal α-tocopherol supplementation resulted in significantly lower ROS levels in SR-BI−/− embryos, similar to the fluorescence levels in SR-BI+/+ embryos. Trophoblast giant cells have been shown to produce significant ROS due to high activity of the enzyme NADPH oxidase18. However, SR-BI deficiency did not affect ROS levels in parietal yolk sacs from embryos obtained from mothers fed control or vitamin E-enriched diets (Fig. 3b). Together, these results show that excess ROS in SR-BI−/− embryos can be prevented by maternal α-tocopherol supplementation.

ROS index in embryos (a) and parietal yolk sac (PYS) (b) obtained from SR-BI+/− dams fed with control or vitamin E supplemented diets. Levels of reactive oxygen species were measured using DCF-DA in pairs of wild-type embryos (SR-BI+/+), normal knock-out embryos (nSR-BI−/−) and knock-out embryos with NTD (SR-BI−/− NTD) (a) and single PYS (b). *p ≤ 0.05, **p ≤ 0.01, determined by one-way ANOVA and Tukey’s post-test.
High ROS production and oxidative stress are associated with an antioxidant response that is characterized by up-regulation of the expression of several enzymes that metabolize ROS into harmless compounds. Using real time PCR, we compared the expression of genes encoding antioxidant enzymes in SR-BI+/+ and SR-BI−/− embryos (with or without NTD) as well as in their respective PYS, from dams fed chow or vitamin E-enriched diets. Our results showed similar expression of Gsr, Cat, Sod2, Txn2 and Glrx among embryos of different genotypes, phenotypes or maternal diets (Fig. 4). We also evaluated the expression of Ppargc1a, a target and master regulator of the antioxidant response19, and found that its expression was also similar among the embryos from different groups (Fig. 4). In PYS, minor changes with unclear biological significance were found in Cat, Txn2 and Glrx gene expression (Supplementary Fig. 3), independent of genotype, phenotype, or dietary treatment. We did not compare the expression of Ppargc1a in PYS because the mRNA levels were below the detection limit in this tissue.

Antioxidant gene expression response in embryos obtained from SR-BI+/− dams fed with control or vitamin E supplemented diets. Expression levels of genes activated during antioxidant response were measured in pools of 3 wild-type embryos (SR-BI+/+), normal knock-out embryos (nSR-BI−/−) and knock-out embryos with NTD (SR-BI−/− NTD). N = 3 pools per group.
Expression of genes involved in neural tube closure in SR-BI−/− embryos
As gene expression orchestrates the molecular and cellular processes that take place during neural tube closure, many mutations in mouse genes coding for transcription factors give rise to NTD20. Thus, we tested whether abnormal gene expression contributed to NTD in SR-BI−/− embryos and whether this expression was normalized after maternal α-tocopherol supplementation. We compared the mRNA levels for genes known to be relevant for neural tube closure in SR-BI+/+ and SR-BI−/− embryos of both phenotypes (normal or NTD) obtained from control chow- or vitamin E-fed dams. We first analysed the expression of genes coding for proteins involved in the Hedgehog (Hh) signalling pathway, one of the main regulators of neural tube closure and neuronal specification21, 22. We observed similar expression for Hh gene targets in SR-BI−/− embryos compared to SR-BI+/+ embryos (Supplementary Fig. 4). We also checked the mRNA levels for Pax3, a key paired-box transcription factor whose inactivation leads to NTD with total penetrance in the Splotch mouse23, 24. Pax3 expression is significantly reduced in murine models of maternal diabetes, in association with an embryonic accumulation of ROS and a partially penetrant NTD phenotype13. Our results showed altered Pax3 expression, specifically in NTD SR-BI−/− embryos from chow-fed dams compared to nSR-BI−/− (Fig. 5a). We also examined gene expression of two members of the aristaless-like (Alx) homeobox protein family that are involved in neural tube closure. One of these genes is Alx3, whose inactivation induces a partially penetrant NTD phenotype25. Interestingly, reduction in Alx3 expression is observed in mouse embryos deficient for Lrp2, a multiligand receptor mediating HDL endocytosis26. Our results showed that expression of Alx3 was significantly reduced in NTD SR-BI−/− embryos compared to SR-BI+/+ embryos and to nSR-BI−/− embryos (Fig. 5b). Maternal treatment with vitamin E normalized Alx3 expression in SR-BI−/− embryos. Another member of the aristaless-like family of proteins that is involved in neural tube closure is Alx1 27, which has an expression domain and function that are partly redundant with Alx3 28. Alx1 expression in NTD SR-BI−/− embryos was 8-fold lower than that in nSR-BI−/− embryos (Fig. 5b). Regardless of genotype, embryos from vitamin E-supplemented dams had higher Alx1 expression than embryos from chow-fed dams.

Expression of neural tube closure-related transcription factors in embryos obtained from SR-BI+/− dams fed with control or vitamin E supplemented diets. Expression levels of Pax3 (a) and Alx transcription factors (b) were determined in pools of 3 E9.5 wild-type embryos (SR-BI+/+), normal knock-out embryos (nSR-BI−/−) and knock-out embryos with NTD (SR-BI−/− NTD). N = 3 pools per group.
Discussion
Although several lipids have been shown to be crucial for early development, the molecular mechanisms explaining the transport of these molecules between the mother and embryo or foetus are still not completely understood. Early embryonic nutrition is accomplished by the transport of nutrients from maternal endometrial glands to the embryo through TGC and visceral endoderm cells of the yolk sac. Given the expression of SR-BI in TGC and the high incidence of NTD in SR-BI−/− mouse embryos4, 5, we assessed the role of SR-BI in embryonic vitamin E uptake and its implications for neural tube closure. Our main findings are that SR-BI−/− embryos exhibit defective embryonic vitamin E levels and that maternal α-tocopherol supplementation can almost completely prevent NTD in SR-BI−/− embryos.
In rodents, vitamin E is essential for the success of pregnancy, and maternal deficiency of this vitamin has been associated with NTD in offspring, among other malformations9, 29. In one study describing embryos lacking the main intracellular α-tocopherol binding protein (Ttpa), researchers suggested that vitamin E is required for placental formation and not essential for embryo development itself30. Our results support the idea that uptake of this nutrient is required for neural tube closure during early development, even before the establishment of a fully functional placenta.
Impaired lipoprotein metabolism in the maternal-embryonic interface has been previously linked to neural tube closure abnormalities. For example, genetic inactivation of microsomal triglyceride transfer protein (Mttp) impairs lipoprotein packaging in the endoplasmic reticulum of the visceral endoderm and is associated with embryonic lethality and cranial NTD31. Apob-containing lipoproteins have been shown to be secreted from the visceral endoderm into the embryonic environment32. In various mouse models with inactivating mutations in the Apob gene, homozygosity leads to embryonic lethality, characterized by the presence of cranial NTD in a subset of the mutants33,34,35. Interestingly, embryos lacking Apob have slightly lower cholesterol than wild-type embryos and extremely low levels of vitamin E36, providing further support to the idea that reduced transport of this vitamin may impair neural tube closure. However, in contrast to SR-BI−/− embryos, maternal vitamin E supplementation of dams that are heterozygous for the Apob mutation does not prevent NTD in homozygous embryos37. It is possible that the visceral endoderm entirely relies on Apob and Mttp to secrete vitamin E to Apob-containing lipoproteins, rendering maternal supplementation ineffective. By contrast, TGC may rely on other vitamin E transport mechanism besides SR-BI, as suggested by the expression of diverse proteins that are involved in lipoprotein uptake by receptor-mediated endocytosis in extraembryonic tissues. These pathways might be efficient enough to compensate for the lack of SR-BI in SR-BI−/− TGC, providing them with vitamin E after maternal overloading of this nutrient. The redistribution of this vitamin into different classes of lipoproteins in α-tocopherol-supplemented dams further supports this possibility.
An unexpected result was that, despite the high levels of vitamin E in SR-BI−/− TGC and prevention of NTD in SR-BI−/− embryos, embryonic vitamin E levels were not restored by maternal supplementation with α-tocopherol. Among the possibilities that could explain this finding is that vitamin E transported by SR-BI-independent mechanisms may be oxidized or metabolized rapidly before or during transport to the embryo, becoming undetectable by the methods used. Future experiments involving the detection of vitamin E metabolites in PYS and embryos could aid in testing this hypothesis. Alternatively, there may be an indirect effect on the embryo caused by vitamin E loading of TGC. Although this possibility is not supported by our results, which show similar levels of ROS and mRNA for genes from the antioxidant response in PYS of different genotypes or phenotypes, the use of whole PYS instead of TGCs might have reduced the sensitivity of the assay. In this work, TGCs were not separated from parietal endoderm cells to avoid the potential effects of the isolation procedure on the cellular composition.
Among various genetic and environmental causes of NTD identified in both human and animal experimental studies, nutrient inadequacies are important factors increasing the susceptibility to defective neural tube closure. Over the last few decades, maternal periconceptional supplementation with folic acid has proven to be a safe and efficient intervention to reduce the incidence of human and experimental NTD. However, both folate responsive and folate-resistant NTD have been detected in human and animal embryos, leading researchers to propose the use of combined therapies including folate and other nutrients, such as inositol38 or multivitamin supplements39, to reduce NTD. In this work, our results show that cephalic defective neurulation in SR-BI-deficient embryos can be reduced by supplementing dams with folate or vitamin E. Although we cannot compare the effectiveness of each intervention due to the use of unique doses and different administration routes, our findings of vitamin E deficiency and NTD prevention in SR-BI null embryos support the idea that other nutrients besides folate need to be considered for the prevention of NTD, given the complex and heterogeneous aetiology of this condition.
One of the principles underlying NTD and other congenital malformations in rodents is excessive embryonic oxidative stress, which is observed in vivo in rodent models of maternal diabetes13 and ethanol consumption40, and in mice deficient for thioredoxin 2 (Txn2), a protein scavenging ROS in mitochondria41. Vitamin E has proven to be effective in preventing ROS-induced NTD in murine models both in vivo 13, 42 and in vitro 43. In this work, normalization of ROS levels in SR-BI−/− embryos after maternal α-tocopherol supplementation suggests an antioxidant effect of this vitamin E13. However, we cannot rule out, at present, the existence of additional non-antioxidant effects of α-tocopherol contributing to the results described.
It is worth noting that although all SR-BI−/− embryos had low levels of vitamin E, only approximately half of the embryos exhibited NTD. The incomplete penetrance of NTD in isogenic embryos that developed in a homogeneous uterine environment has been previously observed in mouse models20. Phenotypic discordance for disease susceptibility has also been shown in monozygotic human twins44, 45, including twins discordant for anencephaly46. Studies in C. elegans have suggested that inter-individual stochastic variations in gene expression and activation of compensatory mechanisms could account for different phenotypic consequences of mutations in those organisms47. Similarly, we detected that SR-BI−/− embryos that underwent neural tube closure exhibited higher mRNA levels for a subset of genes involved in neural tube closure compared to SR-BI−/− with NTD, including Pax3 and 2 genes of the aristaless-like family (Alx1 and Alx3). The fact that SR-BI−/− embryos obtained from vitamin E-supplemented dams exhibited similar or even higher expression of those genes than embryos from chow-fed dams suggests that normalization of the expression of those genes might contribute to the prevention of NTD in our model. It cannot be discounted, however, that deficiencies in mRNA levels for those genes may be a consequence and not a cause of failed neurulation.
Despite the involvement of SR-BI in human cholesterol homeostasis and cardiovascular function, no null mutations and very few loss-of-function mutations in the human gene coding for SR-BI have been reported48, 49. Considering that SR-BI mutations are very rare in humans and that SR-BI is expressed in first trimester human trophoblasts, it could be speculated that null or severely deficient SR-BI gene expression might hinder human embryonic development. In humans, vitamin E supplementation during mid and late pregnancy has proven to be inefficient in preventing stillbirth, neonatal death, preterm birth, pre-eclampsia or poor foetal growth50. Two retrospective studies have suggested a relationship between maternal vitamin E consumption, estimated through food questionnaires, and offspring NTD51, 52. In one of the studies, a reduction of the incidence of anencephaly, and not of other NTD, was only observed in the third quartile of vitamin E consumption51. In the second study, a higher intake of vitamin E and other micronutrients was associated with decreased risk of spina bifida52. Although the incidence of NTD in various human populations has been successfully reduced by folic acid consumption, in particular in countries where primary preventive strategies have been implemented53, this malformation has not completely been abolished. In this regard, ensuring adequate vitamin E levels by appropriate nutritional counselling for pregnant women could aid in reducing the remaining risk of human NTD. This could be especially beneficial for pregnant women with disorders associated with a higher oxidative status, such as obesity and type 1 and 2 diabetes, all of which exhibit a higher risk for NTD54,55,56. Short-term vitamin E supplementation, although not free from potential associated risks, could also be envisioned as a potential intervention for women willing to conceive after one or more previously folate-resistant NTD-affected pregnancies.
Methods
RNA-Seq analysis pipeline
We analysed the dataset generated by Hannibal et al.14, which was produced by RNA sequencing of polyA+ RNA from isolated mouse E9.5 parietal TGC. The Single End Reads RNA raw sequence files (.fastq files) were extracted from GEO Accession Number GSE50585. We used the entries GSM1223565, GSM1223566, GSM1223567 and GSM1223568 to generate our data, which underwent quality control analysis using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Single End RNAseq reads were groomed using FASTQ groomer V1.0457, and groomed reads were mapped to the mouse genome (mm10) using TopHat2 version 0.758. Aligned reads were counted using HTseq-count Version 1.0.059 to generate a digital expression matrix. To ensure that all of the sequences were processed consistently, all of the above steps were performed as part of a Galaxy workflow, which can be found at https://usegalaxy.org/u/laiumiunix/w/rnaseq.
A list of all of the lipoprotein receptors was obtained from the Gene Ontology Consortium using the “Lipoprotein particle receptor activity” and “Regulation of plasma lipoprotein particle levels” search terms. The positive controls were marker genes of parietal TGC, whereas the negative controls were markers of TGC from a different lineage that does not give rise to parietal TGC60. We considered a gene to be expressed by TGC when the mean read count of that gene was superior to the mean +3 SD of Tpbpa, the negative control with the highest count. This criterion set the threshold at 71 mean reads.
Animals
We used mice in a mixed C57Bl6/J ×129 background carrying a targeted mutation in the SR-BI locus, generously provided by Monty Krieger (B6;129S2-Scarb1tm1Kri/J)3. Animals were maintained in the animal facility of the Department of Gastroenterology (School of Medicine, Pontificia Universidad Católica de Chile) at 25 °C and 12 h light:dark cycling and fed standard chow (Prolab RMH3000, Labdiet; 75 IU vitamin E/kg) and water ad libitum. Protocols were conducted in agreement with the National Research Council (NRC) publication Guide for Care and Use of Laboratory Animals (copyright 1996, National Academy of Science). Studies were approved by the Ethics Committee for Animal Welfare from the School of Medicine of the Pontificia Universidad Católica de Chile.
To generate intercrosses, 2- to 4-month-old heterozygous (SR-BI+/−) females were caged with 2- to 6-month-old SR-BI+/− males at a 1:1 or 2:1 ratio. Female mice were checked daily for the presence of a vaginal plug during the first hour of the light cycle. The day a plug was detected was recorded as embryonic day 0.5 (E0.5) and the female was separated from the male and maintained with one or two females in the same cage. All embryos were collected on day E9.5, when neural tube closure is complete in wild-type embryos. Pregnant dams were anaesthetized with a mixture of ketamine:xylazine (0.18 mg:0.012 mg per gram of body weight) and the peritoneal cavity was exposed. A blood sample was taken from the abdominal vena cava and uteri were excised. Implantation sites were retrieved and the embryos and PYS were collected. Embryos were assessed for neural tube closure and classified into normal or NTD. The visceral yolk sac was snap frozen in liquid nitrogen, preserved at −80 °C and used for individual genotyping. Embryo and PYS samples were also immediately frozen in liquid nitrogen and stored individually at −80 °C until use. As each dam produced variable numbers of embryos with different genotypes and phenotypes, pools of embryos retrieved from different dams were used.
For all experiments analysing the impact of vitamin E dietary supplementation, pregnant dams were fed α-tocopherol-enriched chow (Prolab 5P00 diet enriched with 2,000 I.U. α-tocopherol/kg, PMI Nutrition International, Richmond, IN) only after mating, from E0.5 to E9.5. Only in the experiment shown in Table 3, some of the dams were fed with the vitamin E-enriched diet from E4.5 to E9.5. The diet was kept at −20 °C until use and stored for no longer than 18 months, and feed in the cages was changed at least every two days. The α-tocopherol-enriched diet contained 1.1 ppm folic acid, which was very similar to the folic acid concentration in the chow diet.
Folic acid (Sigma, MO) in PBS was injected i.p. daily (10 mg/kg body weight)61 to pregnant dams from E0.5 to E9.5. As a control, pregnant dams were injected i.p. with PBS during the same period.
Lipoprotein separation
Whole plasma (200 µl) was chromatographically fractionated by Fast Protein Liquid Chromatography (FPLC) using a Superose-6 column (GE Life Sciences, PA). The mobile phase was an aqueous solution at pH 7.4 containing 150 mM NaCl and 1 mM EDTA, at a constant flow of 9 ml/hour. The first 15 ml was discarded, and then 40 fractions of 300 µl each were recovered. To determine the α-tocopherol content of the various lipoprotein classes, the fractions corresponding to each lipoprotein class (as indicated in Supplementary Fig. 1B) were pooled.
Cholesterol determination
Cholesterol in whole plasma (10 µl) and FPLC fractions (300 µl) was measured with the enzymatic method reported by Allain et al.62. Samples were incubated for 30 minutes at 37 °C with reaction buffer (0.5 M Tris pH 7.6, 50 mM phenol, 50 mM 4-chlorophenol, 1% Triton X-100, 0.37% sodium cholate, 0.04% 4-aminoantipyrine, 0.35 U/ml cholesterol esterase, 0.1 U/ml cholesterol oxidase, 1.1 U/ml peroxidase). The absorbance was measured at 490 nm in a plate reader.
Vitamin E determination
Pairs of embryos or single PYS were lysed in RIPA buffer containing 1% N-acetylcysteine and centrifuged at 12,000 g at 4 °C for 10 minutes. Lysates, as well as whole plasma (200 µl) and pooled FPLC fractions (1.5 to 3 ml), were extracted for 1 hour in 500 µl methanol (0.01% butylated hydroxytoluene) and 5 ml hexane. Methanol and hexane mixtures were centrifuged for 5 minutes at 1,000 g at room temperature, and the supernatants were recovered and dried under nitrogen. Dried extracts were resuspended in methanol:ethanol 1:1 (200 µl of mixture) and filtered through a 0.22-µm PTFE filter. The samples were subjected to HPLC through a Symmetry LC-8 reverse phase column (Waters, MA). The mobile phase used was 20 mM sodium perchlorate in methanol:water 97.5:2.5 at a constant flux of 1 ml/minute controlled by a L-6000 HPLC Pump (Hitachi, IL). Elution signal was detected with a LC-4C amperometric detector (Bioanalytical Systems, Inc., IN) at 600 mV vs. Ag/AgCl. The samples were run in triplicate and the signal was interpolated in a standard curve for α-tocopherol. Among the various vitamin E isomers, the only detectable isomer in both PYS and embryos was α-tocopherol.
Reactive oxygen species analysis
The presence of reactive oxygen species was detected using the method from Chen63 with modifications. Pairs of embryos and single parietal yolk sacs were kept on ice and lysed by sonication for 2 seconds in 100 µl reaction buffer (130 mM KCl, 5 mM MgCl2, 20 mM KH2PO4, 20 mM Tris, 30 mM glucose). The lysates were incubated with dihydrodichlorofluorescein diacetate (DCF-DA; Sigma, MO) at a final concentration of 50 µM, and fluorescence was detected continuously for 1 hour at 37 °C with an excitation wavelength of 485 nm and a detection wavelength of 535 nm in a plate reader. For each sample, fluorescence was adjusted based on the protein content. Representative results of this assay for embryos and PYS are shown in Supplementary Fig. 2. Negative controls, including lysate-free samples and DCF-DA free samples, showed no fluorescence. As a positive control, lysates were incubated with H2O2 (100 µM final concentration) alone or together with N-acetylcysteine (NAC; 0.1% final concentration) just prior to the addition of DCF-DA. Incubation of lysates with H2O2 resulted in a significant increase in fluorescence that was reduced by incubation of lysates with H2O2 and NAC.
Sex determination
Individual sexing of embryos was performed by allele discrimination using PCR, as described previously64. The primer sequences F: 5′-CCGCTGCCAAATTCTTTGG-3′ and R: 5′-TGAAGCTTTTGGCTTTGAG-3′ were used to amplify the bands corresponding to the smcx and y alleles in the X and Y chromosomes, respectively.
Real time PCR
Total RNA was extracted from pools of 3 female embryos and individual parietal yolk sacs using the PureLink RNA Micro Kit (Invitrogen, CA), following the manufacturer’s instructions. The embryos used to make the pools came from 9 control litters and 6 vitamin E-supplemented litters. RNA integrity was evaluated using the Bioanalyser 2100 (Agilent, CA) with the Eukaryote Total RNA Nano assay (Agilent, CA). All samples had an RNA Integrity Number of 10. Purified RNA (500 ng) was used for retrotranscription with the iScript RT Supermix (Biorad, CA). The resulting cDNA was amplified by real time PCR with a StepOnePlus thermocycler (Applied Biosystems, CA) using the PowerUp SYBR Green master mix (Thermo, MA) and 100 nM of each primer. The primers, annealing temperatures and amplification efficiencies are listed in Supplementary Table 1. All primers were designed using NCBI’s Primer-BLAST65. The amplification conditions were as follows: 5 minutes at 95 °C and 40 cycles of 15 seconds at 95 °C, 15 seconds of annealing and 30 seconds at 72 °C. After every reaction, a melting curve was performed to ensure the amplification of a single product. The amplification efficiency of each pair of primers was determined by serial dilution of a mixture of the cDNAs. Then, the relative expression was calculated for each sample using the equation by Pfaffl66 (equation 1 in the reference) and the TATA-box binding protein (Tbp) as reference gene.
Statistics
Sample sizes were calculated to achieve an 80% power of detecting a 2-fold change with α = 0.05. For supplementation with α-tocopherol after implantation, we sought an 80% power to detect a 95% reduction in the presence of NTD in SR-BI−/− embryos with α = 0.05.
The assignment of pregnant dams to each treatment group was pseudo-randomized. Each day, the first female with a vaginal plug was assigned to the control group, the second one to one of the treatment groups, and so on. If only one female had a plug one day, the next day the order was reversed. The phenotypic assessments were performed blinded to the genotype of the embryo, but not to the treatment group. Biochemical and real time PCR experiments were performed blind to the genotype and the treatment group of the sample.
Results are shown as scatter plots with a horizontal line indicating the mean (or median where indicated) for arithmetic data, mean ± SEM for lipoprotein profiles and geometric mean + error for exponential data obtained from real time PCR experiments. The error represents the uncertainty in estimating the relative expression and is computed using Taylor’s series relative to the control group67. Therefore, error is reported only for the non-control groups.
The statistical significance of the difference between proportions was evaluated with the Fisher’s exact test. Differences between arithmetical means were tested for significance using one-way ANOVA with a Tukey’s post-hoc test or two-way ANOVA with the Holms-Sidak post-test. If variances were different between groups, then a non-parametric test was used (Mann-Whitney for two group comparison and Kruskal-Wallis with Dunn’s post-test for multigroup comparison). The significance of the difference in gene expression was tested using the Pair-wise Fixed Reallocation Randomization test with the Relative Expression Software Tool67 Multiple Comparison Solution14.
All tests were two-sided, and results were considered significant at p ≤ 0.05. The statistically significant differences between groups are symbolized by asterisks (*p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001).
References
Acton, S. et al. Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science 271, 518–520 (1996).
Rigotti, A., Miettinen, H. E. & Krieger, M. The role of the high-density lipoprotein receptor SR-BI in the lipid metabolism of endocrine and other tissues. Endocrine reviews 24, 357–387 (2003).
Rigotti, A. et al. A targeted mutation in the murine gene encoding the high density lipoprotein (HDL) receptor scavenger receptor class B type I reveals its key role in HDL metabolism. Proceedings of the National Academy of Sciences of the United States of America 94, 12610–12615 (1997).
Hatzopoulos, A. K., Rigotti, A., Rosenberg, R. D. & Krieger, M. Temporal and spatial pattern of expression of the HDL receptor SR-BI during murine embryogenesis. Journal of lipid research 39, 495–508 (1998).
Santander, N. G. et al. Developmental abnormalities in mouse embryos lacking the HDL receptor SR-BI. Human molecular genetics 22, 1086–1096, doi:10.1093/hmg/dds510 (2013).
Lindegaard, M. L. et al. Characterization of placental cholesterol transport: ABCA1 is a potential target for in utero therapy of Smith-Lemli-Opitz syndrome. Human molecular genetics 17, 3806–3813 (2008).
Woollett, L. A. Review: Transport of maternal cholesterol to the fetal circulation. Placenta 32(Suppl 2), S218–221 (2011).
Mardones, P. et al. Alpha-tocopherol metabolism is abnormal in scavenger receptor class B type I (SR-BI)-deficient mice. The Journal of nutrition 132, 443–449 (2002).
Evans, H. M. & Bishop, K. S. On the Existence of a Hitherto Unrecognized Dietary Factor Essential for Reproduction. Science 56, 650–651, doi:10.1126/science.56.1458.650 (1922).
Packer, L., Weber, S. U. & Rimbach, G. Molecular aspects of alpha-tocotrienol antioxidant action and cell signalling. The Journal of nutrition 131, 369S–373S (2001).
Rimbach, G. et al. Regulation of cell signalling by vitamin E. The Proceedings of the Nutrition Society 61, 415–425 (2002).
Cheng, D. W., Bairnson, T. A., Rao, A. N. & Subbammal, S. Effect of variations of rations on the incidence of teratogeny in vitamin E-deficient rats. The Journal of nutrition 71, 54–60 (1960).
Chang, T. I. et al. Oxidant regulation of gene expression and neural tube development: Insights gained from diabetic pregnancy on molecular causes of neural tube defects. Diabetologia 46, 538–545, doi:10.1007/s00125-003-1063-2 (2003).
Hannibal, R. L. et al. Copy number variation is a fundamental aspect of the placental genome. PLoS Genet 10, e1004290 (2014).
Watkins, T. et al. gamma-Tocotrienol as a hypocholesterolemic and antioxidant agent in rats fed atherogenic diets. Lipids 28, 1113–1118 (1993).
Busso, D. et al. Early onset intrauterine growth restriction in a mouse model of gestational hypercholesterolemia and atherosclerosis. BioMed research international 2014, 280497 (2014).
Nicolosi, R. J., Lawton, C. W. & Wilson, T. A. Vitamin E reduces plasma low density lipoprotein cholesterol, LDL oxidation, and early aortic atherosclerosis compared with black tea in hypercholesterolemic hamsters. Nutrition Research 19, 1201–1214 (1999).
Bevilacqua, E., Gomes, S. Z., Lorenzon, A. R., Hoshida, M. S. & Amarante-Paffaro, A. M. NADPH oxidase as an important source of reactive oxygen species at the mouse maternal-fetal interface: putative biological roles. Reproductive biomedicine online 25, 31–43 (2012).
St-Pierre, J. et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell 127, 397–408 (2006).
Harris, M. J. & Juriloff, D. M. An update to the list of mouse mutants with neural tube closure defects and advances toward a complete genetic perspective of neural tube closure. Birth defects research. Part A, Clinical and molecular teratology 88, 653–669 (2010).
Copp, A. J. & Greene, N. D. Neural tube defects–disorders of neurulation and related embryonic processes. Wiley interdisciplinary reviews. Developmental biology 2, 213–227 (2013).
Briscoe, J. & Small, S. Morphogen rules: design principles of gradient-mediated embryo patterning. Development 142, 3996–4009 (2015).
Auerbach, R. Analysis of the developmental effects of a lethal mutation in the house mouse. Journal of Experimental Zoology 127, 305–329 (1954).
Goulding, M. et al. Analysis of the Pax-3 gene in the mouse mutant splotch. Genomics 17, 355–363 (1993).
Lakhwani, S., Garcia-Sanz, P. & Vallejo, M. Alx3-deficient mice exhibit folic acid-resistant craniofacial midline and neural tube closure defects. Developmental biology 344, 869–880 (2010).
Kur, E., Mecklenburg, N., Cabrera, R. M., Willnow, T. E. & Hammes, A. LRP2 mediates folate uptake in the developing neural tube. Journal of cell science 127, 2261–2268 (2014).
Zhao, Q., Behringer, R. R. & de Crombrugghe, B. Prenatal folic acid treatment suppresses acrania and meroanencephaly in mice mutant for the Cart1 homeobox gene. Nature genetics 13, 275–283 (1996).
ten Berge, D. et al. Mouse Alx3: an aristaless-like homeobox gene expressed during embryogenesis in ectomesenchyme and lateral plate mesoderm. Developmental biology 199, 11–25 (1998).
Cheng, D. W., Chang, L. F. & Bairnson, T. A. Gross observations on developing abnormal embryos induced by maternal vitamin E deficiency. The Anatomical record 129, 167–185 (1957).
Jishage, K.-i. et al. Vitamin E is essential for mouse placentation but not for embryonic development itself. Biology of reproduction 73, 983–987 (2005).
Raabe, M. et al. Knockout of the abetalipoproteinemia gene in mice: reduced lipoprotein secretion in heterozygotes and embryonic lethality in homozygotes. Proceedings of the National Academy of Sciences of the United States of America 95, 8686–8691 (1998).
Terasawa, Y. et al. Apolipoprotein B-related gene expression and ultrastructural characteristics of lipoprotein secretion in mouse yolk sac during embryonic development. Journal of lipid research 40, 1967–1977 (1999).
Homanics, G. E. et al. Targeted modification of the apolipoprotein B gene results in hypobetalipoproteinemia and developmental abnormalities in mice. Proceedings of the National Academy of Sciences of the United States of America 90, 2389–2393 (1993).
Huang, L. S. et al. apo B gene knockout in mice results in embryonic lethality in homozygotes and neural tube defects, male infertility, and reduced HDL cholesterol ester and apo A-I transport rates in heterozygotes. The Journal of clinical investigation 96, 2152–2161 (1995).
Farese, R. V. Jr., Ruland, S. L., Flynn, L. M., Stokowski, R. P. & Young, S. G. Knockout of the mouse apolipoprotein B gene results in embryonic lethality in homozygotes and protection against diet-induced hypercholesterolemia in heterozygotes. Proceedings of the National Academy of Sciences of the United States of America 92, 1774–1778 (1995).
Farese, R. V. et al. A novel function for apolipoprotein B: lipoprotein synthesis in the yolk sac is critical for maternal-fetal lipid transport in mice. Journal of lipid research 37, 347–360 (1996).
Homanics, G. E. et al. Exencephaly and hydrocephaly in mice with targeted modification of the apolipoprotein B (Apob) gene. Teratology 51, 1–10 (1995).
Greene, N. D., Leung, K. Y. & Copp, A. J. Inositol, neural tube closure and the prevention of neural tube defects. Birth defects research 109, 68–80, doi:10.1002/bdra.23533 (2017).
Wolf, H. T., Hegaard, H. K., Huusom, L. D. & Pinborg, A. B. Multivitamin use and adverse birth outcomes in high-income countries: a systematic review and meta-analysis. American journal of obstetrics and gynecology, doi:10.1016/j.ajog.2017.03.029 (2017).
Miller-Pinsler, L. & Wells, P. G. Embryonic catalase protects against ethanol embryopathies in acatalasemic mice and transgenic human catalase-expressing mice in embryo culture. Toxicology and applied pharmacology 287, 232–239, doi:10.1016/j.taap.2015.06.007 (2015).
Nonn, L., Williams, R. R., Erickson, R. P. & Powis, G. The absence of mitochondrial thioredoxin 2 causes massive apoptosis, exencephaly, and early embryonic lethality in homozygous mice. Molecular and cellular biology 23, 916–922 (2003).
Reece, E. A., Wu, Y. K., Zhao, Z. & Dhanasekaran, D. Dietary vitamin and lipid therapy rescues aberrant signaling and apoptosis and prevents hyperglycemia-induced diabetic embryopathy in rats. American journal of obstetrics and gynecology 194, 580–585 (2006).
Francis, A. J., Anderson, D., Jenkinson, P. C. & Parke, D. V. The protective effects of L-ascorbic acid and DL-alpha-tocopherol on cultured rat embryos treated with xanthine/xanthine oxidase. Mutation research 214, 137–145 (1989).
Czyz, W., Morahan, J. M., Ebers, G. C. & Ramagopalan, S. V. Genetic, environmental and stochastic factors in monozygotic twin discordance with a focus on epigenetic differences. BMC medicine 10, 93, doi:10.1186/1741-7015-10-93 (2012).
Baranzini, S. E. et al. Genome, epigenome and RNA sequences of monozygotic twins discordant for multiple sclerosis. Nature 464, 1351–1356, doi:10.1038/nature08990 (2010).
Lim, K. I., Dy, C., Pugash, D. & Williams, K. P. Monoamniotic twins discordant for anencephaly managed conservatively with good outcomes: two case reports and a review of the literature. Ultrasound Obstet Gynecol 26, 188–193, doi:10.1002/uog.1947 (2005).
Burga, A., Casanueva, M. O. & Lehner, B. Predicting mutation outcome from early stochastic variation in genetic interaction partners. Nature 480, 250–253 (2011).
Vergeer, M. et al. Genetic variant of the Scavenger Receptor BI in humans. The New England journal of medicine 364, 136–145 (2011).
Zanoni, P. et al. Rare variant in scavenger receptor BI raises HDL cholesterol and increases risk of coronary heart disease. Science 351, 1166–1171 (2016).
Rumbold, A., Ota, E., Hori, H., Miyazaki, C. & Crowther, C. A. Vitamin E supplementation in pregnancy. The Cochrane database of systematic reviews, Cd004069, doi:10.1002/14651858.CD004069.pub3 (2015).
Chandler, A. L. et al. Neural tube defects and maternal intake of micronutrients related to one-carbon metabolism or antioxidant activity. Birth defects research. Part A, Clinical and molecular teratology 94, 864–874, doi:10.1002/bdra.23068 (2012).
Gilboa, S. M. et al. Maternal intake of vitamin E and birth defects, national birth defects prevention study, 1997 to 2005. Birth defects research. Part A, Clinical and molecular teratology 100, 647–657 (2014).
De-Regil, L. M., Fernandez-Gaxiola, A. C., Dowswell, T. & Pena-Rosas, J. P. Effects and safety of periconceptional folate supplementation for preventing birth defects. The Cochrane database of systematic reviews, CD007950 (2010).
Huang, H. Y., Chen, H. L. & Feng, L. P. Maternal obesity and the risk of neural tube defects in offspring: A meta-analysis. Obesity research & clinical practice. doi:10.1016/j.orcp.2016.04.005 (2016). doi:.
Reece, E. A. Diabetes-induced birth defects: what do we know? What can we do? Current diabetes reports 12, 24–32 (2012).
Kim, H. S. et al. Maternal and Neonatal Outcomes in Korean Women with Type 1 and Type 2 Diabetes. Diabetes & metabolism journal 39, 316–320, doi:10.4093/dmj.2015.39.4.316 (2015).
Blankenberg, D. et al. Manipulation of FASTQ data with Galaxy. Bioinformatics 26, 1783–1785 (2010).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome biology 14, R36 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Hu, D. & Cross, J. C. Development and function of trophoblast giant cells in the rodent placenta. The International journal of developmental biology 54, 341–354 (2010).
Burren, K. A. et al. Gene-environment interactions in the causation of neural tube defects: folate deficiency increases susceptibility conferred by loss of Pax3 function. Human molecular genetics 17, 3675–3685 (2008).
Allain, C. C., Poon, L. S., Chan, C. S. G., Richmond, W. & Fu, P. C. Enzymatic Determination of Total Serum Cholesterol. Clinical Chemistry 20, 470–475 (1974).
Chen, S. Y. Analysis of Nrf2-mediated transcriptional induction of antioxidant response in early embryos. Methods in molecular biology 889, 277–290 (2012).
Jiménez, A. et al. Experimental demonstration that pre- and post-conceptional mechanisms influence sex ratio in mouse embryos. Molecular reproduction and development 66, 162–165 (2003).
Ye, J. et al. Primer-BLAST: a tool to design target-specific primers for polymerase chain reaction. BMC bioinformatics 13, 134 (2012).
Pfaffl, M. W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic acids research 29, e45 (2001).
Pfaffl, M. W., Horgan, G. W. & Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic acids research 30, e36 (2002).
Acknowledgements
This study was financed by the Chilean National Council for Scientific and Technological Research (CONICYT) programs Fondo Nacional del Desarrollo Científico y Tecnológico (FONDECYT) no. 1141236 (to D.B.), no. 1150399 (to A.R.), no. 1140697 (to V.P.), Ph.D. Fellowship no. 21130444 (to N.S.) and the School of Medicine grant PMD-04/16 (to N.S.). The authors acknowledge Ludwig Amigo for his aid in lipoprotein isolation studies by FPLC and Dr. Pablo Olmos for his generous advice on statistical analyses.
Author information
Authors and Affiliations
Contributions
N.S. performed most of the experiments reported in this paper and significantly participated in the preparation of the manuscript and figures. M.J.P. performed folate injections for prevention of NTD. A.Q. maintained the SR-BI+/− mouse colony, genotyped the animals and provided heterozygous SR-BI+/− males and females for matings. D.P. and G.E. set up and performed vitamin E determinations by HPLC. C.L. extracted raw count data from the RNA-Seq dataset. L.U. and V.P. analysed the expression of the Hh pathway genes in the embryos. A.R. provided the mouse model and participated in the design of experiments and discussion of results. D.B. participated in the generation of hypotheses, design of experiments, discussion of the results and preparation of the final version of the manuscript and figures. All of the contributors reviewed the manuscript and provided their feedback before the final submission.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Accession codes: Hannibal et al. generated the RNA dataset used in this study and it is available for download in GEO accession number GSE50585.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Santander, N., Lizama, C., Parga, M.J. et al. Deficient Vitamin E Uptake During Development Impairs Neural Tube Closure in Mice Lacking Lipoprotein Receptor SR-BI. Sci Rep 7, 5182 (2017). https://doi.org/10.1038/s41598-017-05422-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-05422-w
This article is cited by
-
Phytoconstituents for Boosting the Stem Cells Used in Regenerative Medicine
Current Pharmacology Reports (2023)
-
Vitamin E is necessary for zebrafish nervous system development
Scientific Reports (2020)
-
Roles of vitamins in stem cells
Cellular and Molecular Life Sciences (2020)
-
Attenuation of atherogenic apo B-48-dependent hyperlipidemia and high density lipoprotein remodeling induced by vitamin C and E combination and their beneficial effect on lethal ischemic heart disease in mice
Biological Research (2018)
-
Transcriptional profiling of embryos lacking the lipoprotein receptor SR-B1 reveals a regulatory circuit governing a neurodevelopmental or metabolic decision during neural tube closure
BMC Genomics (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
