
Abstract
Understanding the Tibetan Plateau’s palaeogeography and palaeoenvironment is critical for reconstructing Asia’s climatic history; however, aspects of the plateau’s uplift history remain unclear. Here, we report a fossil biota that sheds new light on these issues. It comprises a fossil climbing perch (Anabantidae) and a diverse subtropical fossil flora from the Chattian (late Oligocene) of central Tibet. The fish, Eoanabas thibetana gen. et sp. nov., is inferred to be closely related to extant climbing perches from tropical lowlands in south Asia and sub-Saharan Africa. It has osteological correlates of a labyrinth organ, which in extant climbing perches gives them the ability to breathe air to survive warm, oxygen-poor stagnant waters or overland excursion under moist condition. This indicates that Eoanabas likewise lived in a warm and humid environment as suggested by the co-existing plant assemblage including palms and golden rain trees among others. As a palaeoaltimeter, this fossil biota suggests an elevation of ca. 1,000 m. These inferences conflict with conclusions of a high and dry Tibet claimed by some recent and influential palaeoaltimetry studies. Our discovery prompts critical re-evaluation of prevailing uplift models of the plateau and their temporal relationships with the Cenozoic climatic changes.
Similar content being viewed by others


Introduction
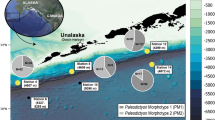
The rise of the Tibetan Plateau is generally held responsible for the climatic cooling and monsoon intensification during the Cenozoic1,2,3. However, the history of the plateau has been fiercely debated, with conflicting uplift models proposed4. Some recent palaeoaltimetry studies, including those isotope-based ones5,6,7, suggest the establishment of near-present high elevations and harsh environment in the Tibetan interior since the Eocene or late Oligocene (~50–26 Ma)5,6,7,8,9 or some high and east-west trending mountain ranges in southern and northern plateau since Palaeocene10. Other tectonic models propose much younger uniform11 or northward stepwise uplift events since late Eocene12. Compared to the prevailing abiotic (tectonic, geophysical and geochemical, etc.) data, palaeobiological evidence bearing on timelines for uplift is scant, especially for the vast interior plateau5,6,7,8,9. Among the biotic proxies, plant megafossils, fishes and other vertebrates are environmentally sensitive and thus promising candidates for constraining the uplift of the plateau13,14,15,16,17,18,19,20,21,22. In addition to a fragmentary forelimb bone of an early Miocene fossil rhino20, only two late Oligocene to early Miocene fossil cyprinid (carps) species have been reported to date from the vast central area of the plateau. One is a primitive snow carp, Plesioschizothorax macrocephalus Wu et Chen16, 19, which has living relatives endemic to the plateau and its periphery17, 18. The other, Tchunglinius tchangii Wang et Wu, is a barbine carp, related to a lineage otherwise currently distributed in South Asia and Africa21. Both of these fishes were probably inhabitants of comparatively low altitudes16,17,18,19, 21. The plant megafossils from the interior of the plateau are even less common; there is but one barberry species formally reported from the Miocene Hox Xil basin in the northern plateau that possibly suggests an elevation ~2–3 km lower than modern-day levels14. Our recent discovery of a dozen well-preserved skeletons of a fossil climbing perch (Perciformes: Anabantidae) (Figs 1 and 2, Supplementary Figs 3–5) together with fifty exquisitely-preserved plant megafossils (Fig. 3) from the upper Oligocene (ca. 26~23.5 Ma) of the Nima and Lunpola basins in central Tibet (Supplementary Figs 1 and 2, and see Supplementary Information) is an important addition to the fossil record of the Tibetan Plateau. These fossils represent a biota that comprises ideal environmental indicators of warm and humid environment at low elevations.

A new fossil climbing perch, Eoanabas thibetana gen. et sp. nov. from the upper Oligocene of central Tibet. It resembles its extant tropical relatives in having a labyrinth organ for air breathing and postocular contact organ in male fishes for stimulating the female during a mating clasp. (a) Photograph and (b) line drawing of holotype (IVPP V22782a), image horizontally rotated. (c) Photograph and (d) line drawing of the head of IVPP V18412a, red area in (d) representing muscular attachment facet. Abbreviations: alm, attachment facet of levator operculi muscle; Cbr1, ceratobranchial of first gill arch; op.st, V-shaped struts on inner side of opercles.

Fossil climbing perch, Eoanabas thibetana gen. et sp. nov. from the upper Oligocene of central Tibet. (a) Line drawing of the head of IVPP V18414a. (b) Photograph of IVPP V18414a. (c) Photograph of IVPP V18581a. (d) Scanning Electron Microscope (SEM) images of relics of labyrinth organ in (c), arrows pointing the pores on the lamellae. (e) Computerized tomography of labyrinth organ (lateral view) of Anabas testudineus (OP 435). (f) Cleared and stained head showing the labyrinth organ and associated structures of Anabas testudineus (collection no. OP 432). (g) Cleared and stained specimen of Anabas testudineus (collection no. OP 433). (h) Osteological restoration of Eoanabas, purported male; not to scale. Images in (c), (d) are horizontally rotated. Abbreviations: br, branchiostegal rays; hp1, hypural 1; m., muscle; php, parhypural.

Plant megafossils associated with Eoanabas, showing the taxonomic and morphological diversity of the plant assemblage. (a) Palmate leaf of palm, IVPP B 2505. (b) Capsular valve of Koelreuteria sp., a golden rain tree, IVPP B 2506. (c) Leaflet of Pistacia sp., IVPP B 2508. (d) Winged fruit of Cedrelospermum sp., IVPP B 2510. (e) Undetermined toothed leaf, IVPP B 2519. (f) Undetermined entire-margined leaf, IVPP B 2518. (g) Undetermined toothed leaf, IVPP B 2527. (h) Infructescence of probable Araceae, IVPP B 2535. (i) Leaf of Araliaceae, IVPP B 2515. (j) Palmately compound leaf of Handeliodendron sp., with six leaflets, IVPP B 2513. (k) Leaf fragment of Limnobiophyllum sp., IVPP B 2514. (l) Leaf of Exbucklandia sp., IVPP B 2516. (m) Leaf of Magnoliales, IVPP B 2526. (n) Undetermined entire-margined leaf, IVPP B 2525. (o) Populus sp., IVPP B 2523. (p) Undetermined entire-margined leaf, IVPP B 2517. (q) Leaf fragment of Typha sp., IVPP B 2529.
The climbing perches (anabantid fishes) are closely related to kissing gouramies and fighting fishes23,24,25 in the perciform suborder Anabantoidei (labyrinth fishes)25. They are common in freshwaters in tropical Asia and sub-Saharan Africa23, 24. Their labyrinth organs are so large that they occupy a large portion of the gill chamber. In fact, their tissues are not sufficient to meet the respiratory needs, and they are therefore obligatory air-breathers24. The capability of accessary air breathing allows some of anabantids to move onto land under moist conditions, or even reputedly climb trees, thus earning the common name “climbing perch”23.
Results
Systematic Palaeontology of Fossil Climbing Perch
Teleostei Müller, 1845
Anabantiformes sensu Wiley and Johnson, 2010
Anabantoidei sensu Lauder and Liem, 1983
Anabantidae Bonaparte, 1839
Eoanabas thibetana gen. et sp. nov.
Etymology. The generic name combines ‘Eo-’ (Greek, early/primeval) with ‘Anabas’, the type genus of Anabantidae from tropical Asia. The specific name refers to Tibet, China.
Holotype. IVPP V 22782, a complete skeleton, part and counterpart (Fig. 1a,b).
Paratypes. Sixteen specimens are designated as paratypes (Supplementary Information).
Locality and Horizon. Jiangnongtangga (type locality) and Songwori in south Nima Basin and Dayu in Lunpola Basin in central Tibet (Supplementary Figs 1 and 2). Middle-upper part of Dingqing Formation, late Oligocene (Chattian) (ca. 26~23.5 Ma)6, 20, 26.
Diagnosis. A labyrinth fish displaying anabantid characteristics including a posterior notch of the opercle bounded by spines, a V-shaped strut on inner side of opercle and six to nine anal-fin spines. It shares with Asian anabantids the following derived characters: broad infraorbitals 3–5 completely covering the cheek, a sensory canal pore just behind sphenotic/pterotic junction and pelvic plate lying flat; and it shares with African anabantids some derived characters, e.g., sensory canal opening in between the infraorbitals, supraorbital commissure of the sensory canal absent and male postocular contact organ present.
Description
The body is oblong with long-based dorsal and anal fins comprising anterior spinous and posterior soft-ray sections, a thoracic pelvic fin anteceded by a spine, and a truncated caudal fin (Figs 1a,b and 2b,c,h, Supplementary Figs 3a,b,i,j, 4a–e, j–m and 5a,b,d–f).
The skull roof is smooth. The frontals carry paired coronal pores on the supraorbital sensory canals (Fig. 1a,b and Supplementary Fig. 3a,b) and, as in the species of African Microctenopoma 24, lack the supraorbital commissure. The circumorbital bones include a ventrally serrated lacrimal and broad infraorbitals 3–5 covering the cheek completely (Figs 1 and 2a, Supplementary Fig. 5a,b). Similarly large infraorbitals are only seen in Anabas (Fig. 2g); however, the sensory pores are open between these elements, a character known only in African anabantids24. Just posterior to the sphenotic/pterotic junction, the pterotic bears a sensory opening (Fig. 1 and Supplementary Fig. 5a,b), which is present in Anabas but not in African forms24. More posteriorly, it has a depressed attachment facet for the levator operculi muscle (Fig. 1c,d and Supplementary Fig. 5a,b) as in living anabantids24. Immediately ventral to this facet is the suprabranchial chamber (Fig. 1c,d) housing the labyrinth organ referred to below. The parasphenoid shaft is straight without oral or orbital processes (Supplementary Figs 3a,b,g and 4j–m), and the transverse process is likely present (Supplementary Fig. 3h).
The opercle has a round posterior notch bounded by two large spines or spine patches (Figs 1c,d and 2a,b, Supplementary Figs 3a,b and 5a,b) and a V-shaped strut on the inner side emerging from the articular socket with the hyomandibular (Figs 1a,b and 2c, Supplementary Figs 3a,b, 4k and 5a,b).
The maxilla is relatively short and stout with a clubbed posterior end (Figs 1 and 2a,b). Stout and recurved teeth occur on the anterior oral edges of the premaxilla and dentary (Figs 1a,b and 2a, Supplementary Figs 3f and 4k).
The ventral postcleithrum directly contacts the flat lying pelvic plate (Fig. 1a,b and Supplementary Fig. 3g), which resembles that of Anabas, but differs from the dorsally angled one in African forms24.
The long-based dorsal and anal fins show relatively low numbers of anal spines (7–9) and soft rays (7–9) in both fins (Figs 1a,b and 2b,c, Supplementary Figs 3a,b,i,j, 4a–e, j–m and 5a,b,d–f, and Supplementary Table 1). The truncated caudal fin has a principal fin-ray formula of I-7/7-I (Figs 1a,b and 2b,c, Supplementary Figs 3a,b,i,j, 4j–m and 5d,e and Supplementary Table 1). The parhypural firmly attaches to the compound vertebra (Fig. 1a,b and Supplementary Fig. 3a,b,i,j), whereas in living anabantids, it is detached and widely separated from the vertebra24 (Fig. 2g).
In one specimen of Eoanabas, layers of bony lamellae just above the branchial chamber (Fig. 2c and Supplementary Fig. 3g,h) show a cribrate structure (Fig. 2d) almost identical to that of the expanded plates of the labyrinth organ (modified first epibranchial)24 in living anabantids (Fig. 2e,f), suggesting the presence of a labyrinth organ also in the fossil fish (Fig. 2h).
Sexual dimorphism is exemplified by the postocular contact organ located immediately posteroventral to the orbit, a patch of scales with large ctenii (Fig. 2a,b and Supplementary Figs 3a,b,d and 5c) that are much larger than those of flank scales (Supplementary Fig. 3a,c,e). This organ is rare among perciform fishes24 and is seen in all African anabantids except for the species of Microctenopoma and Sandelia capensis 24, 27.
The Plant Megafossils Assemblage Associated with Eoanabas
Eoanabas was found in the lacustrine depositional environments with several aquatic herbs (Fig. 3 and Supplementary Information) such as duckweed (Limnobiophyllum, Lemnoideae, Araceae) (Fig. 3k) and bulrush (Typha) (Fig. 3q), associated with a diverse lowland vegetation (Fig. 3 and Supplementary Information) including palms (Fig. 3a), golden rain trees (Koelreuteria) (Fig. 3b), Pistacia (Fig. 3c), Ulmaceae (Fig. 3d), Araliaceae (Fig. 3i), Handeliodendron (Fig. 3j), Exbucklandia (Fig. 3l) and aspens (Populus) (Fig. 3o). Among the 15 types of woody dicot leaves in this plant assemblage, only five types have a toothed leaf margin, whereas the remaining 10 types all have an entire margin (Supplementary Table 2). Regarding the leaf size28, microphyll leaves are dominant in the assemblage, with at least 10 morphotypes.
Discussion
A phylogenetic analysis based on morphological data resolves Eoanabas as the earliest diverging lineage within the family Anabantidae, with its African relatives being monophyletic and sister to the Asian single genus Anabas (Supplementary Fig. 6). Eoanabas is also the oldest anabantid known so far, extending the geological record (previously represented only by three opercular bones from Pleistocene Java)29, 30 of this family for ca. 20 Ma and expanding the geographic distribution of this family to the middle Cenozoic central Tibet.
Living anabantids are confined mostly to the regions in Asia and Africa dominated by the tropical monsoon (Supplementary Fig. 7), with the greatest diversity in the rain forests of western and central Africa24, 31. They inhabit small bodies of water in lowlands in tropical Asia and sub-Saharan Africa23, 24, 31 (Supplementary Table 3), rarely higher than 1,000 m asl31. With few exceptions in the rivers of west Africa24, they prefer warm and periodically oxygen-poor stagnant waters, under the optimal temperature range of 18 to 30 °C (Supplementary Table 3). The key adaptations to such conditions are the labyrinth organ (Fig. 2f), a highly convoluted structure covered by richly vascularized epithelia24, and the resultant accessary air-breathing capability24, 32. Their air-breathing behaviour is facilitated by several muscles, including the levator operculi muscle32 that leaves a characteristic marked facet on the pterotics in living anabantids24. Interestingly, Eoanabas also has a labyrinth organ (Fig. 1d) and similar arrangement of the air-breathing muscle on the pterotics (Fig. 1c,d and Supplementray Fig. 5a,b). These strikingly similar characters appear physiologically specific and environmentally diagnostic31, and thus may indicate similar habitat conditions.
Eoanabas possesses a postocular contact organ in the males, which is unique to certain African anabantids24, 27. This structure indicates a typical anabantid spawning behavior. When the male embraces the female, it thrusts the spines of this organ into her abdomen to stimulate her to release eggs and ensure fertilization27. The similar reproductive behaviour strengthens the ecological similarities between Eoanabas and its living confamilial relatives.
The plant megafossil assemblage found with Eoanabas contains some taxa with living analogues (e.g., the palms and duckweed) commonly associated with the extant climbing perches (Supplementary Information), and hence reinforces the above palaeoenvironmental inference. Although the available material is insufficient to conduct a quantitative climatic reconstruction, the relatively high percentage (>60%) of the entire-margined types among the woody dicot leaves suggests a warm and humid climate (Supplementary Information). The dominance of microphyll leaves is consistent with this climatic inference, and appears to further suggest that the late Oligocene central Tibetan vegetation is more comparable to modern subtropical vegetation in Zhejiang, East China33. Based on the co-existing range of the living analogues of these fossil plants, this assemblage is assumed to grow most likely at an elevation of ca. 1,000 m asl (Supplementary Information).
Collectively, the ecological signal of Eoanabas and associated plant megafossils, as well as the co-existing fossil barbel fish Tchunglinius 21, suggests that warm and humid lowland habitats were present in the Tibetan interior ca. 26 Ma, probably at an elevation ~1,000 m asl, just like the environmental settings for modern climbing perches (Supplementary Fig. 8). Such a vibrant ecosystem represented by a spiny-rayed fish, barbel carp, and a diverse subtropical vegetation is in sharp contrast to that in the central Tibet today15, 17, 18. Yet the general lowland landscape might exhibit some topographic variation within the catchment area according to the contemporaneous pollen data of a mixed coniferous-broadleaved forest26.
Such palaeoenvironmental and palaeotopographic inferences will help us reevaluate our current understanding of the rise of the plateau and its environmental and climatic consequences. A warm and humid central Tibet requires a palaeogeographic setting allowing sufficient warm moisture to reach as far as the plateau interior at least up to the late Oligocene. As such, the current high-standing topographic features in southern Tibet might not exist then to block the southerly moisture from the ocean. This is in conflict with the notions that high Gangdese range has already been in place since the Palaeocene10 or the plateau had undergone northward stepwise uplift from late Eocene time onward12, but is compatible with the predictions of the rise of the Himalaya until the Neogene34 and newly proposed hypothesis for the oceanic Greater Indian Basin that would have maintained a deep tropical water mass between Asia and India throughout most of the Palaeogene35. Given the intrinsic environmental signals of the fossil climbing perch and the highly diversified sub-tropical plants, the establishment of a central Tibet with near-present elevation and climatic condition since early to late Eocene5, 7,8,9 or late Oligocene6 also seems highly problematic. Therefore, we caution against simple links between the rather early uplift of the plateau and the global climatic cooling during the early Cenozoic era9. The generally low elevation in plateau interior and probably more southern areas around the Palaeogene/Neogene transition is an important condition to test the existing tectonic models of plateau uplift. It is compatible with the inferences of either a period of un-roofing2 or the cease of the uplift process and a Tibet no higher than 1,000 m during ca. 30–20 Ma11, but contradicts the models for the constantly thickened crust in central Tibet since at least as early as the late Oligocene6 or even earlier8, 9. Our estimation of a fairly low, warm and wet central Tibet implies that this area has been elevated ~4,000 m to attain its current conditions since latest Palaeogene. In light of our fossil discovery, the geological history of the Tibetan Plateau is in need of critical re-evaluation.
Methods
Fossils were prepared mechanically with sharp needles. The Recent material for comparison was cleared and stained according to the method of Bai36. Photographs of fossils were taken with Nikon except Fig. 2d and Supplementary Fig. 3f,h that were images produced using Hitachi S-3700N Scanning Electron Microscope (SEM) (working condition: Fig. 2d, 3 kV, 98 µA, Working Distance = 25.4 mm; Supplementary Fig. 3f, 5 kV, 99 µA, WD = 16.7 mm; Supplementary Fig. 3h, 3 kV, 101 µA, WD = 25.5 mm); and Fig. 2e that is an X-ray micro-computerized tomography. For the latter, the scanning was carried out at the Key Laboratory of Vertebrate Evolution and Human Origins, Chinese Academy of Sciences (CAS) using the 225 kV micro-computerized tomography (developed by the Institute of High Energy Physics, CAS. The specimen (partial labyrinth organ of Anabas) was scanned with beam energy of 90 kV and a flux of 100 μA at a detector resolution of 6.27 μm per pixel using a 360° rotation with a step size of 0.5° and an unfiltered aluminium reflection target. A total of 720 transmission images were reconstructed in a 2,048*2,048 matrix of 1,563 slices using a two-dimensional reconstruction software developed by the Institute of High Energy Physics, CAS. The three dimensional reconstructions were created in VG Studio Max (v2.0), and images of the reconstructions were exported from VG Studio and finalized in Adobe Photoshop and Adobe Illustrator. Line drawings were done under a Leica MZ8 microscope with a camera lucida attachment and further prepared using Adobe Photoshop CS4.
To conduct a phylogenetic analysis to assess the systematic position of Eoanabas within the family Anabantidae, we developed a data matrix including 165 soft tissue, behavioural, and skeletal morphological characters (Wu et al. 2016.nex, Wu et al. 2016.tnt). These morphological characters are updated from ref. 24 and include 33 newly defined and other characters adopted from references elsewhere (for details see Appendix section below). Of the total 21 taxa, Badis and kissing gouramy Helostoma were designated as outgroups. The ingroup comprises fossil Eoanabas thibetana and 18 living anabantid species and species assemblages according to ref. 24. The data matrix was edited in WinClada 1.00.0837 and saved in the NEXUS format (Wu et al. 2016.nex) and TNT format (Wu et al. 2016.tnt). Additionally, to further test the phylogenetic position of the fossil taxon, we also tried the backbone-constrained search with the relationships based on Rüber et al.’s gene matrix25 of the extant taxa involved in this study used as backbone (datamatix: Wu et al. 2016 backboned.tnt). Parsimony analysis was undertaken using TNT, Tree analysis using New Technology, a parsimony analysis program supported by the Willi Hennig Society38. The analyses were conducted using a traditional search strategy, with default settings except for the following: 10,000 maximum trees in memory and 100 replications. Bremer support values were generated in TNT by applying the ‘New Traditional Search’ using TBR and collecting suboptimal topologies with 100 replicates. All characters were treated as unordered, and weighted equally.
References
Licht, A. et al. Asian monsoons in a late Eocene greenhouse world. Nature 513, 501–506, doi:10.1038/nature13704 (2014).
Li, J.-J. & Fang, X.-M. Research on the uplift of the Qinghai-Xizang Plateau and environmental changes. Chi. Sci. Bull. 43, 1569–1574 (1998).
Molnar, P., Boos, W. R. & Battisti, D. S. Orographic controls on climate and paleoclimate of Asia: Thermal and mechanical roles for the Tibetan Plateau. Annu. Rev. Earth Planet. Sci. 38, 77–102, doi:10.1146/annurev-earth-040809-152456 (2010).
Deng, T. & Ding, L. Paleoaltimetry reconstructions of the Tibetan Plateau: progress and contradictions. Natl. Sci. Rev. 2, 417–437, doi:10.1093/nsr/nwv062 (2015).
Rowley, D. B. & Currie, B. S. Palaeo-altimetry of the late Eocene to Miocene Lunpola basin, central Tibet. Nature 439, 677–681, doi:10.1038/nature04506 (2006).
DeCelles, P. et al. High and dry in central Tibet during the Late Oligocene. Earth Planet. Sci. Lett. 253, 389–401, doi:10.1016/j.epsl.2006.11.001 (2007).
Polissar, P. J., Freeman, K. H., Rowley, D. B., McInerney, F. A. & Currie, B. S. Paleoaltimetry of the Tibetan Plateau from D/H ratios of lipid biomarkers. Earth Planet. Sci. Lett. 287, 64–76, doi:10.1016/j.epsl.2009.07.037 (2009).
Wang, C.-S. et al. Constraints on the early uplift history of the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 105, 4987–4992, doi:10.1073/pnas.0703595105 (2008).
Dai, J.-G. et al. The vast pro to-Tibetan Plateau: New constraints from Paleogene Hoh Xil Basin. Gondwana Res. 22, 434–446, doi:10.1016/j.gr.2011.08.019 (2012).
Ding, L. et al. The Andean-type Gangdese Mountains: paleoelevation record from the Paleocene–Eocene Linzhou Basin. Earth Planet. Sci. Lett. 392, 250–264, doi:10.1016/j.epsl.2014.01.045 (2014).
Zhao, W.-L. & Morgan, W. J. Uplift of Tibetan Plateau. Tectonics 4, 359–369, doi:10.1029/TC004i004p00359 (1985).
Tapponnier, P. et al. Oblique stepwise rise and growth of the Tibet plateau. Science 294, 1671–1677, doi:10.1126/science.105978 (2001).
Spicer, R. A. et al. Constant elevation of southern Tibet over the past 15 million years. Nature 421, 622–624, doi:10.1038/nature01356 (2003).
Sun, B. et al. Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence. Sci. Rep. 5, 10379, doi:10.1038/srep10379 (2015).
Wang, X.-M. et al. Cenozoic vertebrate evolution and paleoenvironment in Tibetan Plateau: Progress and prospects. Gondwana Res. 27, 1335–1354, doi:10.1016/j.gr.2014.10.014 (2015).
Wu, Y.-F. & Chen, Y.-Y. Fossil cyprinid from the late Tertiary of north Xizang, China. Vert. PalAsiat. 18, 15–20 (1980).
Chang, M.-M., Miao, D.-S. & Wang, N. In Darwin’s heritage today: Proceedings of the Darwin 200 Beijing International Conference (eds Long, M.-Y., Gu, H.-Y. & Zhou, Z.-H.) 60–75 (Higher Education, 2010).
Cao, W.-X., Chen, Y.-Y., Wu, Y.-F. & Zhu, S.-Q. In The comprehensive scientific expedition to the Qinghai-Xizang Plateau, Studies on the Period, Amplitude and Type of Uplift of the Qinghai-Xizang Plateau. (ed. by Chinese Academy of Sciences) 118–130 (Science Press, 1981).
Chang, M.-M. et al. Extraordinarily thick-boned fish linked to the aridification of the Qaidam Basin (northern Tibetan Plateau). Proc. Natl. Acad. Sci. USA 105, 13246–13251, doi:10.1073/pnas.0805982105 (2008).
Deng, T. et al. A mammalian fossil from the Dingqing Formation in the Lunpola Basin, northern Tibet, and its relevance to age and paleo-altimetry. Chin. Sci. Bull. 57, 261–269, doi:10.1007/s11434-011-4773-8 (2012).
Wang, N. & Wu, F.-X. New Oligocene cyprinid in the central Tibetan Plateau documents the pre-uplift tropical lowlands. Ichthyol. Res. 62, 274–285, doi:10.1007/s10228-014-0438-3 (2015).
Chang, M.-M. & Miao, D.-S. Review of the Cenozoic fossil fishes from the Tibetan Plateau and their bearings on paleoenvironment (in Chinese with English abstract). Chin. Sci. Bull. 61, 981–995 (2016).
Tim, M. B. Freshwater Fish Distribution (Univ. Chicago Press, 2007).
Norris, S. M. The osteology and phylogenetics of the Anabantidae (Osteichthyes, Perciformes). Ph. D. Dissertation (Arizona State Univ., Tempe, 1994).
Rüber, L., Britz, R. & Zardoya, R. Molecular phylogenetics and evolutionary diversification of labyrinth fishes (Perciformes: Anabantoidei). Syst. Biol. 55, 374–397, doi:10.1080/10635150500541664 (2006).
Sun, J.-M. et al. Palynological evidence for the latest Oligocene−early Miocene paleoelevation estimate in the Lunpola Basin, central Tibet. Palaeogeogr. Palaeoclimatol. Palaeoecol. 399, 21–30, doi:10.1016/j.palaeo.2014.02.004 (2014).
Cambray, J. A. The spawning behaviour of the endangered Eastern Cape rocky, Sandelia bainsii (Anabantidae), in South Africa. Environ. Biol. Fish. 49, 293–306, doi:10.1023/A:1007316519904 (1997).
Wolfe, J. A. A method of obtaining climatic parameters from leaf assemblages. US Geol. Surv. Bull. 2040, 1–71 (1993).
Liem, K. F. The comparative osteology and phylogeny of the Anabantoidei (Teleostei, Pisces). Illinois Biol. Monogr. 30, 1–149 (1963).
Koumans, F. P. On some fossil fish remains from Java. Zool. Med. 5, 77–82 (1949).
Forselius, S. Studies of anabantid fishes. Zool. Bidrag. Uppsala 32, 93–597 (1957).
Liem, K. F. Functional design of the air ventilation apparatus and overland excursions by teleosts. Fieldiana (Zool.) 37, 1–29 (1987).
Wang, B. Phytocoenology (Higher Edu. Press, 1987).
Wang, Y., Deng, T. & Biasatti, D. Ancient diets indicate significant uplift of southern Tibet after ca. 7 Ma. Geology 34, 309–312, doi:10.1130/G22254.1 (2006).
Lippert, P. C., van Hinsbergen, D. J. J. & Dupont-Nivet, G. Early Cretaceous to present latitude of the central proto-Tibetan Plateau: a paleomagnetic synthesis with implications for Cenozoic tectonics, paleogeography, and climate of Asia. Geol. Soc. Am. Spec. Pap. 507, 1–21 (2014).
Bai, F.-X. The method to make cleared and stained fish skeleton. Technol. Info. 2, 128 (2012).
Nixon, K. C. WinClada. ver. 1.00.08. Published by the author, Ithaca, NY (2002).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786, doi:10.1111/cla.2008.24.issue-5 (2008).
Acknowledgements
We thank E.M. Friis and J. Qiu for helpful suggestions; B. Kang, S.O. Kullander, T. Su, Y. Lin and J. Xue for valuable discussions; Z. Wang for preparing and W. Gao, G. Xu, W. Zhang for photographing fossils; L. Dong, F. Shi, Y. Rong, Y. Chen, C. Jin, F. Wu and other colleagues for help in collecting fossils; X. Sa, B. Kang for supplying extant specimens; Y. Zhao and L. Liang for helping with making cleared and stained specimens; and two anonymous reviewers for their helpful suggestions. This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (grant XDB03020104), the National Basic Research Program of China (grant 2012CB821900), National Natural Science Foundation of China (grants 41430102, 41272028 and 41206173), the Excavation Funding and Emphatic Deployed Project of IVPP, CAS to F. Wu, and Youth Scholars Program of Beijing Normal University to N. Wang.
Author information
Authors and Affiliations
Contributions
F.W. and N.W. identified the fossil fish and designed the research; F.W. performed the research; F.W. analyzed the data and wrote the paper; D.M. and M.C. participated in the research and writing; G.S. identified the fossil plants and participated in the research and writing.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, F., Miao, D., Chang, Mm. et al. Fossil climbing perch and associated plant megafossils indicate a warm and wet central Tibet during the late Oligocene. Sci Rep 7, 878 (2017). https://doi.org/10.1038/s41598-017-00928-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-00928-9
This article is cited by
-
Paleogene integrative stratigraphy, biotas, and paleogeographical evolution of the Qinghai-Tibetan Plateau and its surrounding areas
Science China Earth Sciences (2024)
-
The Sino-Himalayan flora evolved from lowland biomes dominated by tropical floristic elements
BMC Biology (2023)
-
Molecular phylogeny of mega-diverse Carabus attests late Miocene evolution of alpine environments in the Himalayan–Tibetan Orogen
Scientific Reports (2023)
-
Cenozoic plants from Tibet: An extraordinary decade of discovery, understanding and implications
Science China Earth Sciences (2023)
-
Reorganization of Asian climate in relation to Tibetan Plateau uplift
Nature Reviews Earth & Environment (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
