
Abstract
Little is known regarding the underlying neurobiology of smoking cessation. Neuroimaging studies indicate a role for the insula in connecting the interoceptive awareness of tobacco craving with a larger brain network that motivates smoking. We investigated differences in insula-based functional connectivity between smokers who did not relapse during a quit attempt vs those who relapsed. Smokers (n=85) underwent a resting-state functional connectivity scan and were then randomized into two groups (either smoking usual brand cigarettes or smoking very low nicotine cigarettes plus nicotine replacement therapy) for 30 days before their target quit date. Following the quit date, all participants received nicotine replacement therapy and their smoking behavior was observed for 10 weeks. Participants were subsequently classified as nonrelapsed (n=44) or relapsed (i.e., seven consecutive days of smoking ⩾1 cigarette/day; n=41). The right and left insula, as well as insula subdivisions (posterior, ventroanterior, and dorsoanterior) were used as seed regions of interest in the connectivity analysis. Using the right and left whole-insula seed regions, the nonrelapsed group had greater functional connectivity than the relapsed group with the bilateral pre- and postcentral gyri. This effect was isolated to the right and left posterior insula seed regions. Our results suggest that relapse vulnerability is associated with weaker connectivity between the posterior insula and primary sensorimotor cortices. Perhaps greater connectivity in this network improves the ability to inhibit a motor response to cigarette cravings when those cravings conflict with a goal to remain abstinent. These results are consistent with recent studies demonstrating a positive relationship between insula-related functional connectivity and cessation likelihood among neurologically intact smokers.
Similar content being viewed by others
INTRODUCTION
Tobacco use continues to be the leading preventable cause of death in the United States (U.S. Department of Health and Human Services 2014). Despite extensive research on the neurobiological mechanisms underlying tobacco addiction and relapse during a quit attempt (e.g., Benowitz 2010; Lester 2011), relatively little is known about the mechanisms involved in successful tobacco cessation. However, the ability to resist drug craving is believed to rely on the proper function of neuronal circuits that regulate top-down control of behavior (Volkow and Baler 2014). In order to shed more light on the neurocircuitry of smoking cessation, the present study investigated differences in resting-state functional brain connectivity between smokers who did not relapse during a 10-week quit attempt compared with those who relapsed.
Recent studies on the neurobiology of addictive disorders, including tobacco addiction, have implicated the insula in the subjective, interoceptive awareness of drug craving (Garavan 2010; Naqvi et al, 2014). Insula activation is consistently reported in functional imaging studies of drug craving and cue reactivity, along with activation in other interconnected corticolimbic regions including the striatum, cingulate, and prefrontal cortex (Garavan 2010). Among smokers, insula activation has been shown to correlate with self-reported craving evoked by cigarette cues (Brody et al, 2002; Franklin et al, 2009), and tobacco use and dependence have been negatively associated with insula cortical thickness (Morales et al, 2014); however, insula lesions have been associated with abrupt disruption of tobacco addiction (Naqvi et al, 2007). Although the exact relationship between insula, tobacco use, and behavioral control is not yet known, the insula is believed to provide conscious awareness of the aversive subjective experience of craving as part of a larger brain network that contributes to the motivation to use tobacco or other drugs of abuse (Garavan 2010).
The neurobiological effects of chronic tobacco use and acute tobacco abstinence have been investigated using functional connectivity analyses, which determine the temporal dependency of neuronal activation patterns across different brain regions (van den Heuvel and Pol 2010). Several studies examining resting-state functional connectivity have shown that 11- to 12-h abstinent smokers exhibit greater connectivity between the insula and default mode network compared with satiated smokers and nonsmokers (Huang et al, 2014; Wang et al, 2014), and the intensity of abstinence-induced craving positively correlates with connectivity between the insula and brain regions involved in interoception and self-awareness, such as the precuneus, during smoking cue exposure (Moran-Santa Maria et al, 2014) and at rest (Huang et al, 2014). Alternatively, one study reported weaker connectivity during a smoking-cue reactivity task between an insula-containing network and brain regions involved in cognitive control among smokers who lapsed during a quit attempt, compared with those who maintained abstinence (Janes et al, 2010). Altogether, this research suggests that insula connectivity mediates the relationship between cigarette craving and the cognitive control of smoking behavior.
Many studies identify connectivity with the insula as a whole; however, the insula is not a homogenous cortical region. Functional connectivity and histological studies have shown that the insula can be subdivided into two or three distinct functional subdivisions (Kurth et al, 2010; Cauda et al, 2011; Deen et al, 2011; Chang et al, 2013; Klein et al, 2013). These subdivisions have unique connections and functions: the posterior subdivision connects with primary and secondary somatosensory regions, and is functionally related to interoception and sensorimotor information; the dorsoanterior subdivision connects with dorsal anterior cingulate cortex and prefrontal cortex, and is functionally related to cognitive control and attention; and the ventroanterior subdivision connects with the anterior cingulate cortex and the temporal cortex, and is functionally related to emotion and chemosensation (Cauda et al, 2011; Deen et al, 2011; Chang et al, 2013). These insular subdivisions may have unique roles in maintaining addiction. In particular, the dorsoanterior insula may help support smoking cessation because of the role it has in cognitive control (Craig 2009; Ullsperger et al, 2010; Chang et al, 2013; Klein et al, 2013).
Taken together, these findings provide strong support for the role of insula connectivity in smoking behavior. Our study extends this prior research by using a seed-based functional connectivity approach in order to specifically isolate insular subdivisions potentially involved in relapse risk in a large sample of treatment-seeking smokers. Our participants completed a resting-state functional neuroimaging scan before being randomized into one of two 30-day pre-quit treatment conditions. Following the pre-quit treatment, participants initiated a quit attempt supported by standard nicotine replacement therapy on a target quit-date and smoking behavior was recorded over a 10-week observation period. Based on our review of previous research, we hypothesized that smokers who relapsed would exhibit weaker functional connectivity between the dorsoanterior insula and cognitive control regions (e.g., anterior cingulate, prefrontal cortex, and sensorimotor cortices) compared with smokers who did not relapse.
MATERIALS AND METHODS
Participants
Smokers were recruited from the Durham/Chapel Hill/Raleigh region of North Carolina via newspaper and internet advertisements. Participants were included in the study if they were in general good health, right-handed, aged 18–55 years, smoked⩾10 cigarettes/day, and had an interest in quitting smoking. The exclusion criteria consisted of significant health problems, contraindications for magnetic resonance imaging (MRI), use of psychoactive medications, use of smokeless tobacco or nicotine replacement therapy, current drug or alcohol abuse (based on self-report), use of illicit drugs (based on a urine drug screen), a positive urine pregnancy test (among women), or an afternoon expired carbon monoxide (CO) level<10 p.p.m. using a handheld monitor (Vitalograph, Lenexa KS). On the scan day, participants with a positive breath alcohol level (Alert breathalyzer; Columbia Laboratory Supplies) were excluded from participation that day. Participants provided written informed consent, and this study was approved by the Institutional Review Board at Duke University.
Procedure
Participants completed an initial screening/training session in which they were administered a smoking history questionnaire and the Fagerström Test of Nicotine Dependence (Heatherton et al, 1991). They were also familiarized with a mock MRI scanner. The following week, participants underwent an initial MRI scan in their native cigarette-satiated state (participants were asked to smoke 30 min before the scan). Then, participants were randomized into two treatment groups for an experimental smoking cessation program that lasted 30 days. During these 30 days, participants were instructed to either continue smoking a typical number of their usual brand cigarettes (group 1), or to smoke an equivalent number of very low nicotine cigarettes (<0.05 mg/cigarette; Quest brand, Vector Tobacco) and wear a standard 21-mg nicotine replacement skin patch (Nicoderm CQ, GlaxoSmithKline) every day (group 2). Following their quit-date, all participants underwent standard nicotine replacement therapy (21 mg/day for 6 weeks, 14 mg/day for 2 weeks, and 7 mg/day for 2 weeks) using skin patches (Nicoderm CQ, GlaxoSmithKline). Advice on quitting and minimal support was provided as requested by individual participants.
Participants returned to the laboratory to report their smoking behavior on four occasions spaced across 10 weeks. Daily diaries of cigarette and nicotine patch use were collected at each visit as well as expired CO to confirm reported abstinence or relapse. Consistent with other fMRI/treatment outcome studies that have used continuous measures of abstinence (Grusser et al, 2004; Kosten et al, 2006), the primary smoking cessation outcome variable was relapse, defined as seven consecutive days of smoking cigarettes (at least one cigarette/day) following the quit day. Participants who met this criterion within the 10-week observation period were grouped as ‘relapsed’. Using an intent-to-treat approach, participants lost to contact during these 10 weeks were considered to have relapsed to smoking on the last date of contact. Participants who did not meet the 7-day relapse criterion within the 10-week period were grouped as ‘nonrelapsed’. In order to promote truthful reports, participation in the study (and hence payment for participation) was not contingent upon abstinence. All participants were encouraged to maintain abstinence and to attend all laboratory visits for the duration of the study.
These results are part of a larger study on the effects of an experimental smoking cessation treatment on smoking cessation outcomes and cue reactivity. Only the methods that pertain to the results presented here are described. Methods and outcomes that pertain to other parts of the study will be published elsewhere.
Image Acquisition
MRI scanning was conducted on a 3T General Electric Signa EXCITE HD scanner (Milwaukee, WI) equipped with 40 mT/m gradients. A high-resolution three-dimensional fast spoiled gradient recalled echo (3D-SPGR) anatomical sequence was collected (TR=7.468 ms, TE=2.976 ms, field of view=25.6 cm2, matrix=256 × 256, flip angle=25°, 166 slices, and slice thickness=1 mm).
A gradient-recalled inward spiral pulse imaging sequence (SENSE spiral) was used to collect a resting-state functional connectivity scan (TR=1500 ms, TI=0, TE=30 ms, flip angle=60°, acquisition matrix=64 × 64, field of view=25.6 cm2, and slice thickness=3.8 mm resulting in size 4 × 4 × 3.8 mm voxels, 30 axial slices, 196 volumes for a total duration of 4 min 54 s). During the resting-state scan, participants were instructed to keep their eyes closed. An infrared camera attached to the head coil was used to monitor alertness. These images were collected as part of a 90-min scanning session; following the resting-state scan, a perfusion MRI, diffusion tensor imaging, and two functional MRI scans were also acquired; results from these scans will be reported elsewhere.
Image Preprocessing and Data Analysis
Functional data were preprocessed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm). Images were slice time corrected via sinc interpolation and were realigned to correct for head motion over the course the resting-state scan. Realignment provided the spatial deviation for translational (x, y, z) and rotational (pitch, roll, yaw) movements for each time point. Anatomical images were segmented into native space gray matter and coregistered to the mean of functional images. Functional images were then normalized to the Montreal Neurologic Institute template. During normalization, images were high-pass filtered (filter width=128 s) and voxels were resampled to 2 × 2 × 2 mm. Finally, images were smoothed using a Gaussian kernel with a full-width at half-maximum of 8 mm.
Motion outliers were detected using the artifact detection toolbox (http://www.nitrc.org/projects/artifact_detect/). Time points in individuals’ scans were marked as outliers if the global signal exceeded three SD’s from the mean or if scan to scan motion exceeded 1 mm deviation (Redcay et al, 2013). Individuals were excluded if >20% of time points were identified as outliers; however, no participants met this exclusion criterion. Voxel-wise regression analyses were conducted on each seed region with the six motion parameters from realignment and each outlier time point as separate covariates of no interest.
The functional connectivity toolbox Conn (Whitfield-Gabrieli and Nieto-Castanon 2012) was used to assess resting-state functional connectivity. Conn implements an anatomical CompCor method (Behzadi et al, 2007) to account for physiological noise from individuals’ segmented white matter and cerebral spinal fluid voxels, and temporal derivatives are included to allow for temporal lags or other dynamics between regions.
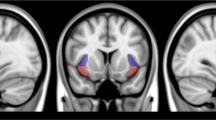
Functional connectivity measures were computed between a seed region of interest (ROI) and every other voxel in the brain. WFU pickatlas (Maldjian et al, 2003) was used to create right and left whole-insula ROIs, drawn from the Automated Anatomical Labeling atlas. These ROIs were loaded into MRIcron (www.mccauslandcenter.sc.edu/mricro/) and divided into posterior (p-INS), dorsoanterior (da-INS), and ventroanterior (va-INS) segments (after Deen et al, 2011) by deleting a 4-voxel-width margin between each segment in order to minimize the shared signal; see Figure 1. Significant main effects of the whole left and right insula seed regions were followed up with the left and right segmented seed regions.

The left and right whole-insula regions of interest were subdivided into posterior (shown in blue), dorsoanterior (shown in red), and ventroanterior (shown in yellow) segments. Significant main effects of the whole left and right insula seed regions were followed up with the left and right segmented seed regions. A full color version of this figure is available at the Neuropsychopharmacology journal online.
Whole-brain regression analyses were conducted for each of the eight ROIs on each individual’s preprocessed data. This resulted in a Fisher-transformed z-value in each brain voxel. Two-way between-group (nonrelapsed vs relapsed) t-tests were run for each of the matrices to discover whether differences in connectivity strength were present between groups. Treatment group and FTND scores were entered as covariates of no interest in each of the t-tests to control for potential differences between pre-quit treatment and tobacco dependence.
Patterns of whole-brain functional connectivity with each seed region were first examined across all subjects using family-wise error correction (P<0.001, KE=100 voxels). Then, between-group analyses (nonrelapsed vs relapsed) were performed and differences were considered significant if they passed a statistical threshold of P<0.05, cluster corrected. Cluster correction was determined using AlphaSim and running 10 000 Monte Carlo simulations on a whole-brain mask composed of 233 847 voxels (Ward 2000). This produced an individual voxel threshold of P<0.001; along with an extent threshold of ⩾70 contiguous significant voxels required for an overall corrected false-positive detection rate of P<0.05. Figures were made using MRIcron.
RESULTS
Participants
A total of 95 participants were scanned; however, nine participants withdrew from the study before initiating a quit attempt and were not included in the analyses. One additional participant was removed from the data set due to poor imaging data quality. Thus, the final sample size consisted of 85 participants (54% female, 22% non-white, age mean±SD=38±10). Participants smoked 19±6 cigarettes/day, had smoked for 19±9 years, and had FTND scores of 5±2. Forty-four participants were assigned to treatment group 1 and 41 participants were assigned to treatment group 2.
Smoking Cessation Outcomes
The nonrelapsed group consisted of 44 participants who did not relapse throughout the 10-week follow-up period, and the relapsed group consisted of 41 participants who relapsed; see Table 1. Of the 44 participants who did not relapse, the median number of cigarettes reportedly smoked during the 10-week follow-up period was 2.0 (SD=28.8). Of the 41 participants who relapsed, 14 were lost to contact before the 10-week period had ended and were presumed to have relapsed. Differences in demographical characteristics between the two groups were analyzed with χ2 and independent samples t-tests. At baseline, the nonrelapsed group smoked fewer cigarettes/day (16±5 vs 22±6) and had lower FTND scores (4.6±2.0 vs 5.5±1.6) than the relapsed group (P-value <0.05). No other differences were found between the groups.
Whole-Insula ROI: Overall Connectivity
As shown in Supplementary Figure S1, both right and left whole-insula seed regions showed robust functional connectivity with bilateral insula, extending into supramarginal gyrus, planum temporale, central operculum cortex, and frontal operculum cortex; bilateral anterior cingulate cortex, extending into the paracingulate cortex, and supplementary motor area; bilateral frontal poles; bilateral posterior cingulate; and bilateral thalamus.
Insula Subdivisions: Overall Connectivity
As shown in Figure 2, and consistent with prior literature (e.g., Cauda et al, 2011; Deen et al, 2011), the three insula segments (da-INS, va-INS, and p-INS) showed distinct patterns of connectivity with other brain regions. The right da-INS segment connected with bilateral insula, extending into the supramarginal gyrus and frontal poles; bilateral anterior cingulate, extending into the paracingulate cortex and supplementary motor cortex; right posterior cingulate and right precuneus cortex. The right va-INS segment connected with bilateral insula, extending into the posterior middle temporal gyrus and frontal orbital cortex; bilateral anterior cingulate, extending into the paracingulate and superior frontal gyrus; bilateral posterior cingulate; bilateral angular gyrus; and right frontal pole. The right p-INS segment connected with bilateral insula, extending into parietal operculum cortex; bilateral precentral and postcentral gyrus; bilateral anterior cingulate, extending into supplementary motor cortex; and right posterior cingulate. The left insula segments revealed similar patterns of connectivity.

Functional connectivity with the right posterior (shown in blue), dorsoanterior (shown in red), and ventroanterior (shown in yellow) insula segments. Regions of overlapping connectivity between the dorsoanterior and ventroanterior segments are shown in orange, between the dorsoanterior and posterior segments are shown in purple, and among the posterior, dorsoanterior, and ventroanterior segments are shown in brown. FWE-corrected P<0.001, KE >100 voxels.
Whole-Insula ROI: Group Differences in Connectivity
Between-group t-tests were conducted to investigate differences between the nonrelapsed and relapsed groups, controlling for the effects of treatment group and FTND scores. When comparing relapse outcomes using the right whole-insula seed, the nonrelapsed group had greater connectivity than the relapsed group with right precentral and postcentral gyri, right lingual gyrus, bilateral putamen, and bilateral p-INS; see Supplementary Figure S2 and Table 2. The left whole-insula seed region showed a similar pattern of greater connectivity for the nonrelapsed group than the relapsed group with bilateral precentral and postcentral gyri, although no differences were seen in bilateral putamen or p-INS; see Table 2.
Insula Subdivisions: Group Differences in Connectivity
Using the right p-INS as a seed, the nonrelapsed group had greater connectivity than the relapsed group with bilateral pre- and postcentral gyri, and bilateral putamen. Similarly, the left p-INS also showed greater connectivity in the nonrelapsed group than the relapsed group with bilateral precentral and postcentral gyri; see Figure 3.

Group differences (nonrelapsed group >relapsed group) in functional connectivity using the right posterior (shown in blue), dorsoanterior (shown in red), and ventroanterior (shown in yellow) insula segments as a seed regions of interest. Cluster corrected, P<0.05. Color bars indicate T-values. A full color version of this figure is available at the Neuropsychopharmacology journal online.
The right and left va-INS showed greater connectivity in the left superior frontal gyrus and the left lingual gyrus, respectively, among the nonrelapsed group than in the relapsed group.
Finally, the right da-INS segment had greater connectivity with the left p-INS in the nonrelapsed group than in the relapsed group.
There was no connectivity greater in the relapsed group than in the nonrelapsed group with any of the seed regions. See Supplementary Information for post hoc analyses.
DISCUSSION
The results of this study provide novel evidence that smokers who relapse during a quit attempt have weaker functional connectivity between the posterior insula and the primary sensorimotor cortices. These effects were observed in resting-state functional imaging scans taken 30 days before a quit attempt, among smokers who desired to quit smoking. Approximately half of the participants relapsed (defined as seven consecutive days of smoking⩾1 cigarette) within the 10-week post-quit follow-up period, and the other half of participants were considered nonrelapsed. Three insula segments (p-INS, da-INS, and va-INS) were used as seed ROIs in a connectivity analysis. We hypothesized that the da-INS segment would reveal the greatest difference in connectivity between groups. Instead, our primary results implicate the p-INS. This region is thought to be related to the visceral interoception of craving and sensorimotor action. Our results support the idea that insula connectivity is related to relapse vulnerability, and a stronger connection between the primary interoceptive region of the insula and the motor control regions improves the likelihood of abstinence among smokers during a quit attempt.
In general, the insula interfaces between physiological sensations and cognition, and it is believed to process affective states in order to detect salient information then recruit attentional resources and cognitive control (Chang et al, 2013). Naqvi et al (2014) proposed that the function of the insula in addiction is to ‘represent the interoceptive effects of drug taking in the service of goal-directed drug seeking’. These interoceptive effects may include aversive drug-craving sensations; for instance, the insula response to negative emotional information has been shown to correlate with tobacco withdrawal-induced negative affect (Froeliger et al, 2012). The p-INS, in particular, may be specifically involved in processing somatosensory stimuli with motivational or affective significance (Augustine 1996; Craig 2009; Deen et al, 2011), and in a meta-analysis it has been shown to be more activated by smoking cues than by neutral cues among smokers (Engelmann et al, 2012). We found that the p-INS is functionally connected to the pre- and postcentral gyri, which are the primary motor and somatosensory cortices, and the putamen, which is part of the basal ganglia that regulates movement. Putamen activation has been related to cigarette cue reactivity during smoking abstinence (McClernon et al, 2009), and the basal ganglia and primary motor cortices are also vital for inhibiting a prepotent motor response, as shown in go/no go and stop-signal tasks (Kelly et al, 2004; Garavan et al, 2006). In fact, a combined analysis of functional connectivity during stop-signal studies revealed that response inhibition involves insula, inferior frontal cortex, pre-supplementary motor area, basal ganglia, and thalamus (Congdon et al, 2010).
Poor inhibitory control, considered a form of impulsivity, has an important role in drug abuse, and there is evidence of poor inhibitory control on go/no-go tasks among smokers compared with controls (for review see Perry and Carrol 2008). A study by Berkman et al (2011) showed that response inhibition-related activation in inferior frontal gyrus, pre-supplementary motor area, and basal ganglia related to a decreased association between cigarette cravings and smoking behavior during a quit attempt. One interpretation of this evidence is that interoceptive cues related to cravings and the urge to smoke are processed by the p-INS, which then relays this information to the motor and somatosensory regions. These regions may represent the sensorimotor aspects of craving and/or motor preparation for smoking behavior. However, it is these same motor regions that may help inhibit smoking behavior when it conflicts with a different cognitive goal, namely to resist smoking. Stronger connectivity within this network could bolster the inhibition of smoking behavior elicited by cravings, or inhibit the cravings themselves. Alternatively, perhaps the motor/somatosensory cortices, basal ganglia, and insula are inhibited by yet other brain regions (e.g., frontal cortex and thalamus) to effectively block relapse; direct pathways between the insula and the motor/somatosensory cortex may not be critical. Future studies could investigate the directionality of connectivity within this circuit during cigarette craving and the suppression of craving to address these questions.
Other functional connectivity studies have investigated differences between smokers and nonsmokers, or differences between smoking satiated and abstinent conditions (e.g., Huang et al, 2014; Lerman et al, 2014; Wang et al, 2014; Weiland et al, 2014). However, to our knowledge, only one previous study has investigated differences between lapsed/relapsed and abstinent smokers. Janes et al (2010) used independent component analysis to examine functional connectivity during a cue reactivity paradigm; the authors reported weaker connectivity among smokers who lapsed shortly after a quit attempt in a network including the insula, prefrontal cortex, anterior cingulate cortex, primary motor and sensory cortices, putamen, and other regions (Janes et al, 2010). The authors suggest that lapsed subjects had poorer cognitive control of interoceptive awareness and were less able to regulate the emotional response to smoking-related cues (Janes et al, 2010). Despite methodological differences, both of these studies report that weaker functional connectivity in an insula-containing network is associated with an increased likelihood of relapse during a quit attempt.
The finding of weakened connectivity as a marker of relapse is somewhat unexpected, given the literature on smoking cessation among stroke patients. In 2007, Naqvi et al reported that insula lesions in smokers could result in abrupt smoking cessation without subsequent cravings or urges to smoke (Naqvi et al, 2007). This was followed-up by a prospective study that showed a trend toward smoking cessation following insula strokes (Suner-Soler et al, 2012), and a recent study that showed lesions encompassing both the basal ganglia and the insula lead to a greater likelihood of quitting smoking (Gaznick et al, 2014). In addition, a lesion specifically to the right posterior cingulate reportedly resulted in an immediate and complete disruption of a patient’s tobacco addiction (Jarraya et al, 2010). Based on this evidence, Naqvi et al (2014) proposed that the insula lesions interfered with conscious craving and suggested that inhibiting insula function (e.g., via transcranial magnetic stimulation, TMS) could be an effective treatment for addiction (Naqvi et al, 2014).
However, if insula lesions disrupted tobacco addiction, then one may expect weaker connectivity with the insula to be associated with increased likelihood of maintaining abstinence among smokers, which we have shown is not the case. The outcomes of the lesion and functional connectivity studies are not mutually exclusive though. If the p-INS is the seat of conscious craving, then lesioning this region could result in a loss of craving and thus abolish smoking behavior. On the other hand, in an intact brain, the insula communicates the craving sensation to motor and cognitive control regions that must respond or inhibit a response according to one’s smoking goal. Therefore, enhanced connectivity throughout this network could help resist relapse. In fact, a recent study found that high-frequency TMS, which is believed to increase cortical excitability, reduced cigarette consumption and increased smokers’ quitting rate compared with low frequency TMS, which is believed to decrease cortical excitability (Dinur-Klein et al, 2014). Thus, using TMS to enhance functional brain networks could become a new smoking cessation treatment.
This study has a number of strengths, including the large sample size, a pre-randomization fMRI baseline assessment, and a specific focus on a brain region previously implicated in smoking relapse. A limitation of our study was the infrequent biochemical verification of abstinence outcomes. We primarily relied on self-report diaries; however, when a discrepancy occurred between the diaries and biochemical measures of smoking, we preferentially relied on the biochemical measures. These discrepancies may not have always been the result of inaccurate self-reports, but additional verification of the diaries was beyond the scope of the investigation. Also, to confirm our hypothesis that smokers who did not relapse were better at inhibiting or suppressing urges to smoke, it would be useful if we had electronic monitoring of smoking urges throughout the cessation treatment period. Some studies have investigated the functional activation in response to resisting an urge to smoke (Brody et al, 2007; Hartwell et al, 2011). Future research could combine these approaches and correlate functional insula connectivity with resisting an urge to smoke. Finally, we based our insula subdivisions on previous work (Deen et al, 2011), and these subdivisions do not reflect individual differences in anatomy or functional connectivity among our sample. Our results could have been strengthened by drawing ROI segments on individuals’ anatomical images, or by using a sample-based functional connectivity map of the insula similar to previous studies (Deen et al, 2011).
In conclusion, smokers who relapsed during the 10-week follow-up period had less connectivity between the right p-INS, the bilateral sensorimotor cortices, and the bilateral putamen. This result indicates that participants who relapsed had reduced functional connectivity between interoceptive awareness and motor control regions, and suggests that these participants had reduced ability to inhibit a prepotent motor response (i.e., taking out a cigarette and smoking it) to cravings or an urge to smoke. Taken together, this evidence supports the hypothesis that the insula is both a driver of conscious drug urges and a regulator of drug use: the insular cortex connects the interoceptive experience of craving with goal-directed behavior, which could either be drug-seeking behavior or the willful suppression of drug-seeking behavior during a quit attempt. Although this study examined baseline, pre-randomization insula connectivity as a predictor of cessation outcomes, future studies will investigate treatment effects on connectivity and how changes in connectivity relate to risk of relapse.
FUNDING AND DISCLOSURE
This study was supported by R01 DA025876 (FJM) and K01 DA033347 (MAA). FJM is a Site PI on an investigator initiated grant (GRAND) from Pfizer, Inc. JER has consulting and patent purchase agreements with Philip Morris International for nicotine inhaler technology, and has a license agreement with Idea Logic, Inc. for an oral nicotine solution patent. The other authors declare no conflict of interest.
References
Augustine JR (1996). Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res Rev 22: 229–244.
Behzadi Y, Restom K, Liau J, Liu TT (2007). A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage 37: 90–101.
Benowitz NL (2010). Nicotine addiction. N Engl J Med 362: 2295–2303.
Berkman ET, Falk EB, Lieberman MD (2011). In the trenches of real-world self-control: neural correlates of breaking the link between craving and smoking. Psychol Sci 22: 498–506.
Brody AL, Mandelkern MA, London ED, Childress AR, Lee GS, Bota RG et al (2002). Brain metabolic changes during cigarette craving. Arch Gen Psychiatry 59: 1162–1172.
Brody AL, Mandelkern MA, Olmstead RE, Jou J, Tiongson E, Allen V et al (2007). Neural substrates of resisting craving during cigarette cue exposure. Biol Psychiatry 62: 642–651.
Cauda F, D'Agata F, Sacco K, Duca S, Geminiani G, Vercelli A (2011). Functional connectivity of the insula in the resting brain. Neuroimage 55: 8–23.
Chang LJ, Yarkoni T, Khaw MW, Sanfey AG (2013). Decoding the role of the insula in human cognition: functional parcellation and large-scale reverse inference. Cereb Cortex 23: 739–749.
Congdon E, Mumford JA, Cohen JR, Galvan A, Aron AR, Xue G et al (2010). Engagement of large-scale networks is related to individual differences in inhibitory control. Neuroimage 53: 653–663.
Craig AD (2009). How do you feel - now? The anterior insula and human awareness. Nat Rev Neurosci 10: 59–70.
Deen B, Pitskel NB, Pelphrey KA (2011). Three systems of insular functional connectivity identified with cluster analysis. Cereb Cortex 21: 1498–1506.
Dinur-Klein L, Dannon P, Hadar A, Rosenberg O, Roth Y, Kotler M et al (2014). Smoking cessation induced by deep repetitive transcranial magnetic stimulation of the prefrontal and insular cortices: a prospective, randomized controlled trial. Biol Psychiatry 76: 742–749.
Engelmann JM, Versace F, Robinson JD, Minnix JA, Lam CY, Cui Y et al (2012). Neural substrates of smoking cue reactivity: a meta-analysis of fMRI studies. Neuroimage 60: 252–262.
Franklin TR, Lohoff FW, Wang Z, Sciortino N, Harper D, Li Y et al (2009). DAT genotype modulates brain and behavioral responses elicited by cigarette cues. Neuropsychopharmacology 34: 717–728.
Froeliger B, Modlin LA, Kozink RV, Wang L, McClernon FJ (2012). Smoking abstinence and depressive symptoms modulate the executive control system during emotional information processing. Addict Biol 17: 668–679.
Garavan H (2010). Insula and drug cravings. Brain Struct Funct 214: 593–601.
Garavan H, Hester R, Murphy K, Fassbender C, Kelly C (2006). Individual differences in the functional neuroanatomy of inhibitory control. Brain Res 1105: 130–142.
Gaznick N, Tranel D, McNutt A, Bechara A (2014). Basal ganglia plus insula damage yields stronger disruption of smoking addiction than basal ganglia damage alone. Nicotine Tob Res 16: 445–453.
Grusser SM, Wrase J, Klein S, Hermann D, Smolka MN, Ruf M et al (2004). Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology (Berl) 175: 296–302.
Hartwell KJ, Johnson KA, Li XB, Myrick H, LeMatty T, George MS et al (2011). Neural correlates of craving and resisting craving for tobacco in nicotine dependent smokers. Addict Biol 16: 654–666.
Heatherton TF, Kozlowski LT, Frecker RC, Fagerstrom KO (1991). The Fagerstrom test for nicotine dependence: a revision of the Fagerstrom tolerance questionnaire. Br J Addict 86: 1119–1127.
Huang W, King JA, Ursprung WW, Zheng S, Zhang N, Kennedy DN et al (2014). The development and expression of physical nicotine dependence corresponds to structural and functional alterations in the anterior cingulate-precuneus pathway. Brain Behav 4: 408–417.
Janes AC, Pizzagalli DA, Richardt S, Frederick BD, Chuzi S, Pachas G et al (2010). Brain reactivity to smoking cues prior to smoking cessation predicts ability to maintain tobacco abstinence. Biol Psychiatry 67: 1002–1002.
Jarraya B, Brugieres P, Tani N, Hodel J, Grandjacques B, Fenelon G et al (2010). Disruption of cigarette smoking addiction after posterior cingulate damage. J Neurosurg 113: 1219–1221.
Kelly AMC, Hester R, Murphy K, Javitt DC, Foxe JJ, Garavan H (2004). Prefrontal-subcortical dissociations underlying inhibitory control revealed by event-related fMRI. Eur J Neurosci 19: 3105–3112.
Klein TA, Ullsperger M, Danielmeier C (2013). Error awareness and the insula: links to neurological and psychiatric diseases. Front Hum Neurosci 7: 14.
Kosten TR, Scanley BE, Tucker KA, Oliveto A, Prince C, Sinha R et al (2006). Cue-induced brain activity changes and relapse in cocaine-dependent patients. Neuropsychopharmacology 31: 644–650.
Kurth F, Zilles K, Fox PT, Laird AR, Eickhoff SB (2010). A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct Funct 214: 519–534.
Lerman C, Gu H, Loughead J, Ruparel K, Yang YH, Stein EA (2014). Large-scale brain network coupling predicts acute nicotine abstinence effects on craving and cognitive function. JAMA Psychiatry 71: 523–530.
Lester RAJ (2011). Cognitive mechanisms underlying relapse to nicotine. Rev Neurosci 22: 467–470.
Maldjian JA, Laurienti PJ, Kraft RA, Burdette JH (2003). An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 19: 1233–1239.
McClernon FJ, Kozink RV, Lutz AM, Rose JE (2009). 24-h smoking abstinence potentiates fMRI-BOLD activation to smoking cues in cerebral cortex and dorsal striatum. Psychopharmacology 204: 25–35.
Morales AM, Ghahremani D, Kohno M, Hellemann GS, London ED (2014). Cigarette exposure, dependence, and craving are related to insula thickness in young adult smokers. Neuropsychopharmacology 39: 1816–1822.
Moran-Santa Maria MM, Hartwell KJ, Hanlon CA, Canterberry M, Lematty T, Owens M et al (2014). Right anterior insula connectivity is important for cue-induced craving in nicotine-dependent smokers. Addict Biol 20: 407–414.
Naqvi NH, Gaznick N, Tranel D, Bechara A (2014). The insula: a critical neural substrate for craving and drug seeking under conflict and risk. Ann NY Acad Sci 1316: 53–70.
Naqvi NH, Rudrauf D, Damasio H, Bechara A (2007). Damage to the insula disrupts addiction to cigarette smoking. Science 315: 531–534.
Perry JL, Carrol ME (2008). The role of impulsive behavior in drug abuse. Psychopharmacology 200: 1–26.
Redcay E, Moran JM, Mavros PL, Tager-Flusberg H, Gabrieli JDE, Whitfield-Gabrieli S (2013). Intrinsic functional network organization in high-functioning adolescents with autism spectrum disorder. Front Hum Neurosci 7: 1–11.
Suner-Soler R, Grau A, Gras ME, Font-Mayolas S, Silva Y, Davalos A et al (2012). Smoking cessation 1 year poststroke and damage to the insular cortex. Stroke 43: 131–136.
U.S. Department of Health and Human Services (2014) The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General. U.S. Department of Health and Human Services: Rockville, MD, USA (accessed 24 March 2015).
Ullsperger M, Harsay HA, Wessel JR, Ridderinkhof KR (2010). Conscious perception of errors and its relation to the anterior insula. Brain Struct Funct 214: 629–643.
van den Heuvel MP, Hulshoff Pol HEH (2010). Exploring the brain network: a review on resting-state fMRI functional connectivity. Eur Neuropsychopharmacol 20: 519–534.
Volkow ND, Baler RD (2014). Addiction science: Uncovering neurobiological complexity. Neuropharmacology 76: 235–249.
Wang KC, Yang JY, Zhang SY, Wei DT, Hao X, Tu S et al (2014). The neural mechanisms underlying the acute effect of cigarette smoking on chronic smokers. PLoS One 9.
Ward BD (2000) Simultaneous inference for fMRI datahttp://homepage.usask.ca/~ges125/fMRI/AFNIdoc/AlphaSim.pdf.
Weiland BJ, Sabbineni A, Calhoun VD, Welsh RC, Hutchison KE (2014). Reduced executive and default network functional connectivity in cigarette smokers. Hum Brain Mapp 36: 872–882.
Whitfield-Gabrieli S, Nieto-Castanon A (2012). Conn: a functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect 2: 125–141.
Acknowledgements
We thank Rachel Kozink, Matt Hallyburton, and Nicole Kaiser for their assistance with data collection.
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Rights and permissions
About this article

Cite this article
Addicott, M., Sweitzer, M., Froeliger, B. et al. Increased Functional Connectivity in an Insula-Based Network is Associated with Improved Smoking Cessation Outcomes. Neuropsychopharmacol 40, 2648–2656 (2015). https://doi.org/10.1038/npp.2015.114
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2015.114
This article is cited by
-
Nicotine dependence and insula subregions: functional connectivity and cue-induced activation
Neuropsychopharmacology (2023)
-
Not all smokers are alike: the hidden cost of sustained attention during nicotine abstinence
Neuropsychopharmacology (2022)
-
Functional connectivity of the anterior insula during withdrawal from cigarette smoking
Neuropsychopharmacology (2021)
-
Abnormal white matter tracts of insula in smokers
Brain Imaging and Behavior (2021)
-
Increased interregional functional connectivity of anterior insula is associated with improved smoking cessation outcome
Brain Imaging and Behavior (2020)

{kind=link}
{kind=link}
{kind=link}