
Abstract
The glucagon-like peptide-1 (GLP-1) receptor and the glucose-dependent insulinotropic polypeptide (GIP) receptor transduce nutrient-stimulated signals to control beta cell function1. Although the GLP-1 receptor (GLP-1R) is a validated drug target for diabetes1, the importance of the GIP receptor (GIPR) for the function of beta cells remains uncertain2,3,4. We demonstrate that mice with selective ablation of GIPR in beta cells (MIP-Cre:GiprFlox/Flox; Gipr−/−βCell) exhibit lower levels of meal-stimulated insulin secretion, decreased expansion of adipose tissue mass and preservation of insulin sensitivity when compared to MIP-Cre controls. Beta cells from Gipr−/−βCell mice display greater sensitivity to apoptosis and markedly lower islet expression of T cell–specific transcription factor-1 (TCF1, encoded by Tcf7), a protein not previously characterized in beta cells. GIP, but not GLP-1, promotes beta cell Tcf7 expression via a cyclic adenosine monophosphate (cAMP)-independent and extracellular signal–regulated kinase (ERK)-dependent pathway. Tcf7 (in mice) or TCF7 (in humans) levels are lower in islets taken from diabetic mice and in humans with type 2 diabetes; knockdown of TCF7 in human and mouse islets impairs the cytoprotective responsiveness to GIP and enhances the magnitude of apoptotic injury, whereas restoring TCF1 levels in beta cells from Gipr−/−βCell mice lowers the number of apoptotic cells compared to that seen in MIP-Cre controls. Tcf7−/− mice show impaired insulin secretion, deterioration of glucose tolerance with either aging and/or high-fat feeding and increased sensitivity to beta cell injury relative to wild-type (WT) controls. Hence the GIPR-TCF1 axis represents a potential therapeutic target for preserving both the function and survival of vulnerable, diabetic beta cells.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
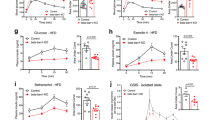
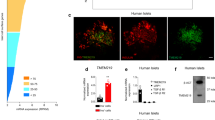

Accession codes
References
Campbell, J.E. & Drucker, D.J. Pharmacology physiology and mechanisms of incretin hormone action. Cell Metab. 17, 819–837 (2013).
Saxena, R. et al. Genetic variation in GIPR influences the glucose and insulin responses to an oral glucose challenge. Nat. Genet. 42, 142–148 (2010).
Speliotes, E.K. et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 42, 937–948 (2010).
Miyawaki, K. et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat. Med. 8, 738–742 (2002).
Mulvihill, E.E. & Drucker, D.J. Pharmacology, physiology and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 35, 992–1019 (2014).
Finan, B. et al. Unimolecular dual incretins maximize metabolic benefits in rodents, monkeys and humans. Sci. Transl. Med. 5, 209ra151 (2013).
Finan, B. et al. A rationally designed monomeric peptide triagonist corrects obesity and diabetes in rodents. Nat. Med. 21, 27–36 (2015).
Shu, L. et al. Decreased TCF7L2 protein levels in type 2 diabetes mellitus correlate with downregulation of GIP- and GLP-1 receptors and impaired beta-cell function. Hum. Mol. Genet. 18, 2388–2399 (2009).
Wessel, J. et al. Low-frequency and rare exome chip variants associate with fasting glucose and type 2 diabetes susceptibility. Nat. Commun. 6, 5897 (2015).
Miyawaki, K. et al. Glucose intolerance caused by a defect in the entero-insular axis: a study in gastric inhibitory polypeptide receptor knockout mice. Proc. Natl. Acad. Sci. USA 96, 14843–14847 (1999).
Meier, J.J. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 8, 728–742 (2012).
Nauck, M.A. et al. Preserved incretin activity of glucagon-like peptide 1 [7–36 amide] but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J. Clin. Invest. 91, 301–307 (1993).
Højberg, P.V. et al. Four weeks of near-normalisation of blood glucose improves the insulin response to glucagon-like peptide-1 and glucose-dependent insulinotropic polypeptide in patients with type 2 diabetes. Diabetologia 52, 199–207 (2009).
Brouwers, B. et al. Impaired islet function in commonly used transgenic mouse lines due to human growth hormone minigene expression. Cell Metab. 20, 979–990 (2014).
Oropeza, D. et al. Phenotypic characterization of MIP-CreERT1Lphi mice with transgene-driven islet expression of human growth hormone. Diabetes 64, 3798–3807 (2015).
Mehran, A.E. et al. Hyperinsulinemia drives diet-induced obesity independently of brain insulin production. Cell Metab. 16, 723–737 (2012).
Chen, S., Okahara, F., Osaki, N. & Shimotoyodome, A. Increased GIP signaling induces adipose inflammation via a HIF-1alpha-dependent pathway and impairs insulin sensitivity in mice. Am. J. Physiol. Endocrinol. Metab. 308, E414–E425 (2015).
Lamont, B.J. & Drucker, D.J. Differential anti-diabetic efficacy of incretin agonists vs. DPP-4 inhibition in high fat fed mice. Diabetes 57, 190–198 (2008).
Ali, S., Lamont, B.J., Charron, M.J. & Drucker, D.J. Dual elimination of the glucagon and GLP-1 receptors in mice reveals plasticity in the incretin axis. J. Clin. Invest. 121, 1917–1929 (2011).
Pamir, N. et al. Glucose-dependent insulinotropic polypeptide receptor null mice exhibit compensatory changes in the enteroinsular axis. Am. J. Physiol. Endocrinol. Metab. 284, E931–E939 (2003).
Pederson, R.A. et al. Enhanced glucose-dependent insulinotropic polypeptide secretion and insulinotropic action in glucagon-like peptide 1 receptor −/− mice. Diabetes 47, 1046–1052 (1998).
Mayo, K.E. et al. International Union of Pharmacology. XXXV. The glucagon receptor family. Pharmacol. Rev. 55, 167–194 (2003).
Kim, S.J. et al. GIP stimulation of pancreatic beta-cell survival is dependent upon phosphatidylinositol 3-kinase (PI3-K)/ protein kinase B (PKB) signaling, inactivation of the Forkhead transcription factor Foxo1 and downregulation of bax expression. J. Biol. Chem. 280, 22297–22307 (2005).
Maida, A., Hansotia, T., Longuet, C., Seino, Y. & Drucker, D.J. Differential importance of GIP versus GLP-1 receptor signaling for beta cell survival in mice. Gastroenterology 137, 2146–2157 (2009).
Shu, L. et al. Transcription factor 7-like 2 regulates beta-cell survival and function in human pancreatic islets. Diabetes 57, 645–653 (2008).
Yusta, B. et al. GLP-1 receptor activation improves β-cell function and survival following induction of endoplasmic reticulum stress. Cell Metab. 4, 391–406 (2006).
Li, Y. et al. Glucagon-like peptide-1 receptor signaling modulates beta cell apoptosis. J. Biol. Chem. 278, 471–478 (2003).
Van de Wetering, M., Castrop, J., Korinek, V. & Clevers, H. Extensive alternative splicing and dual promoter usage generate Tcf-1 protein isoforms with differential transcription control properties. Mol. Cell. Biol. 16, 745–752 (1996).
Ioannidis, V., Beermann, F., Clevers, H. & Held, W. The beta-catenin–TCF-1 pathway ensures CD4+CD8+ thymocyte survival. Nat. Immunol. 2, 691–697 (2001).
Vlotides, G., Eigler, T. & Melmed, S. Pituitary tumor-transforming gene: physiology and implications for tumorigenesis. Endocr. Rev. 28, 165–186 (2007).
Lyssenko, V. et al. Pleiotropic effects of GIP on islet function involves osteopontin. Diabetes 60, 2424–2433 (2011).
Grant, S.F. et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 38, 320–323 (2006).
Florez, J.C. et al. TCF7L2 polymorphisms and progression to diabetes in the Diabetes Prevention Program. N. Engl. J. Med. 355, 241–250 (2006).
Boj, S.F. et al. Diabetes risk gene and Wnt effector Tcf7l2/TCF4 controls hepatic response to perinatal and adult metabolic demand. Cell 151, 1595–1607 (2012).
Kaur, K. et al. Elevated hepatic miR-22–3p expression impairs gluconeogenesis by silencing the Wnt-responsive transcription factor, Tcf7. Diabetes 64, 3659–3669 (2015).
Columbus, J. et al. Insulin treatment and high-fat diet feeding reduces the expression of three Tcf genes in rodent pancreas. J. Endocrinol. 207, 77–86 (2010).
Noble, J.A. et al. A polymorphism in the TCF7 gene, C883A, is associated with type 1 diabetes. Diabetes 52, 1579–1582 (2003).
Erlich, H.A., Valdes, A.M., Julier, C., Mirel, D. & Noble, J.A. Evidence for association of the TCF7 locus with type I diabetes. Genes Immun. 10 (suppl. 1), S54–S59 (2009).
Wang, Z., Moro, E., Kovacs, K., Yu, R. & Melmed, S. Pituitary tumor transforming gene-null male mice exhibit impaired pancreatic beta cell proliferation and diabetes. Proc. Natl. Acad. Sci. USA 100, 3428–3432 (2003).
Chesnokova, V. et al. Diminished pancreatic beta-cell mass in securin-null mice is caused by beta-cell apoptosis and senescence. Endocrinology 150, 10 (suppl. 1) 2603–2610 (2009).
Wicksteed, B. et al. Conditional gene targeting in mouse pancreatic ss-cells: analysis of ectopic Cre transgene expression in the brain. Diabetes 59, 3090–3098 (2010).
Scrocchi, L.A. et al. Glucose intolerance but normal satiety in mice with a null mutation in the glucagon-like peptide receptor gene. Nat. Med. 2, 1254–1258 (1996).
Verbeek, S. et al. An HMG-box-containing T-cell factor required for thymocyte differentiation. Nature 374, 70–74 (1995).
Patterson, J.T. et al. A novel human-based receptor antagonist of sustained action reveals body weight control by endogenous GLP-1. ACS Chem. Biol. 6, 135–145 (2011).
Lamont, B.J. et al. Pancreatic GLP-1 receptor activation is sufficient for incretin control of glucose metabolism in mice. J. Clin. Invest. 122, 388–402 (2012).
Acknowledgements
The authors thank H. Bates for excellent technical assistance and B. Yusta for thoughtful discussions and critical reading of the manuscript; H. Clevers for Tcf7−/− mice; and R. DiMarchi for critical reagents. This work in Toronto was supported by the Canada Research Chairs Program, the Banting and Best Diabetes Centre Novo Nordisk Chair in Incretin Biology, and Canadian Institute for Health Research (CIHR) grants 82700 and 123391 (DJD). We thank the Human Organ Procurement and Exchange (HOPE) program and the Trillium Gift of Life Network (TGLN) for their assistance in obtaining pancreases and islets from human organ donors for research, and we thank J. Lyon (Alberta Diabetes Institute; http://www.bcell.org/isletcore.html) and T. Kin and A.M.J. Shapiro (University of Alberta, Clinical Islet Isolation Facility) for human islet isolation. The Alberta Diabetes Foundation funded the human islet isolations. Work at the University of Alberta was funded by a grant from the Canadian Diabetes Association (to P.E.M.). P.E.M. holds a Canada Research Chair in Islet Biology. Postdoctoral fellowship funding was provided by the Canadian Institute for Health Research (J.E.C., J.R.U., and E.E.M.), Banting and Best Diabetes Centre (J.E.C.), the Canadian Diabetes Association (E.E.M.) and Alberta Innovates Health Solutions (J.R.U.). D.J.D. is the main guarantor of this work and takes responsibility for all content.
Author information
Authors and Affiliations
Contributions
J.E.C. and D.J.D. designed and directed the study, analyzed data and wrote the manuscript. J.R.U., E.E.M., L.L.B. and P.E.M. contributed to the study design and the preparation of the manuscript. J.E.C., J.R.U., E.E.M., J.K., L.L.B., X.C., B.J.L. and T.M. performed experiments. J.K. and P.E.M. carried out experiments on human islets. Y.L. and J.L.W. provided assistance with the RNA-seq data. N.T. and L.H.P. provided MIP-Cre mice. C.J.S. performed the electron microscopy. All authors reviewed the manuscript and provided final approval for submission.
Corresponding author
Ethics declarations
Competing interests
D.J.D. has been a consultant to Novo Nordisk Inc. and other companies that develop and/or sell incretin-based therapies, including Arisaph Pharmaceuticals Inc., Intarcia Therapeutics, Merck Research Laboratories, MedImmune, Receptos, Sanofi, Takeda and Transition Pharmaceuticals Inc. Neither D.J.D. nor his family members hold stock directly or indirectly in any of these companies.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 and Supplementary Tables 1 and 2 (PDF 17197 kb)
Rights and permissions
About this article

Cite this article
Campbell, J., Ussher, J., Mulvihill, E. et al. TCF1 links GIPR signaling to the control of beta cell function and survival. Nat Med 22, 84–90 (2016). https://doi.org/10.1038/nm.3997
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nm.3997
This article is cited by
-
Relevance and consequence of chronic inflammation for obesity development
Molecular and Cellular Pediatrics (2023)
-
The incretin co-agonist tirzepatide requires GIPR for hormone secretion from human islets
Nature Metabolism (2023)
-
The IGFBP3/TMEM219 pathway regulates beta cell homeostasis
Nature Communications (2022)
-
Mechanisms controlling pancreatic islet cell function in insulin secretion
Nature Reviews Molecular Cell Biology (2021)
-
A circular RNA generated from an intron of the insulin gene controls insulin secretion
Nature Communications (2020)
