
Abstract
Oxygen minimum zones (OMZs) currently impinge upon >1 million km2 of sea floor and are predicted to expand with climate change. We investigated how changes in oxygen availability, macrofaunal biomass and retention of labile organic matter (OM) regulate heterotrophic bacterial C and N incorporation in the sediments of the OMZ-impacted Indian continental margin (540–1100 m; [O2]=0.35–15 μmol l−1). In situ pulse-chase experiments traced 13C:15N-labelled phytodetritus into bulk sediment OM and hydrolysable amino acids, including the bacterial biomarker D-alanine. Where oxygen availability was lowest ([O2]=0.35 μmol l−1), metazoan macrofauna were absent and bacteria assimilated 30–90% of the labelled phytodetritus within the sediment. At higher oxygen levels ([O2]=2–15 μmol l−1) the macrofaunal presence and lower phytodetritus retention with the sediment occur concomitantly, and bacterial phytodetrital incorporation was reduced and retarded. Bacterial C and N incorporation exhibited a significant negative relationship with macrofaunal biomass across the OMZ. We hypothesise that fauna–bacterial interactions significantly influence OM recycling in low-oxygen sediments and need to be considered when assessing the consequences of global change on biogeochemical cycles.
Similar content being viewed by others


Introduction
The bathyal continental margins (200–2000 m) are regions of high biogeochemical activity in the deep sea accounting for ∼7% of global sea floor area, but recycle up to 30% of oceanic sedimentary organic matter (OM) (Middelburg et al., 1997). Episodic deposition of phytoplankton-derived detritus (phytodetritus) represents a major OM input to the deep-sea floor (Billett et al., 1983), stimulating the faunal and microbial feeding responses that mediate OM cycling (Witte et al., 2003; Moodley et al., 2005, 2011; Buhring et al., 2006). Oxygen minimum zones (OMZs) are geologically stable water bodies where oxygen concentrations are persistently <22 μmol l−1, occuring in oceanic upwelling regions, where high surface primary productivity combines with poor advective mixing of the water column. At present, OMZs impinge upon >1 million km2 of the sea floor, producing depth-dependent gradients in oxygen availability across the continental margins of the eastern Pacific, northern Indian and southwest Atlantic Oceans (Helly and Levin, 2004). OMZ-impacted sediments experience high OM accumulation and burial rates (for example, Cowie et al., 1999). The low-oxygen conditions within OMZs provide suitable habitat for denitrifying and anaerobic ammonia-oxidising bacteria, with between 30% and 50% of marine nitrogen loss occurring in OMZ-impacted regions (reviewed in Francis et al., 2007). In future climate projections, OMZs are predicted to expand with far-reaching implications for global biogeochemical cycles (Stramma et al., 2010). Changes in oxygen availability and sediment geochemistry across an OMZ influence sediment community composition, producing changes in the faunal and bacterial assemblages that control the rates and pathways for OM recycling (Woulds et al., 2007; Andersson et al., 2008; Gooday et al., 2009; Hunter et al., 2011, 2012; Moodley et al., 2011). Therefore, an understanding of the effects of oxygen limitation upon OM availability, sediment community structure and ecosystem processes is required in order to predict the effects of climate change upon global biogeochemical cycles.
The measurement of N dynamics in OMZs has primarily focussed upon quantifying inorganic N production rates and fluxes, from the sediment and through the water column (for example, Schwartz et al., 2009; Ward et al., 2009; Woulds et al., 2009). Little information is available regarding bacterial utilisation of organic N within OMZs, yet the lability of sedimentary OM is partially determined by the availability of nitrogenous organic compounds, such as amino acids (Dauwe and Middelburg, 1998; Vandewiele et al., 2009). The ecological mechanisms for OM recycling should, therefore, be considered in terms of both C and N processing (Thingstad, 1987). Stable-isotope labelling is a powerful experimental technique that allows C and N to be traced through an ecosystem. To date, application of this technique in OMZ-impacted sediments has focused upon the fate of organic carbon, within shipboard incubations (for example, Woulds et al., 2007; Andersson et al., 2008; Moodley et al., 2011). The advent of a method to trace 15N incorporation into hydrolysable amino acids (HAAs), including the bacteria-specific amino-acid D-alanine, (Veuger et al., 2005, 2007b) makes it possible to empirically assess bacterial N utilisation and de-coupling of C and N in marine sediments (Cook et al., 2007; Veuger and Middelburg, 2007; Evrard et al., 2008).
In situ methods overcome many of the challenges associated with working in deep-sea hypoxic environments, ensuring that neither oxygen contamination nor changes in temperature and pressure influence experimental designs (Glud et al., 1994; Hall et al., 2007). In this study, the deep-sea submersible Shinkai 6500 deployed dual-labelling (13C:15N) pulse-chase experiments at the deep-sea floor. Phytodetrital C and N were traced into sediment bulk OM and HAAs, including the bacterial biomarker D-alanine (D-Ala). We investigate the changes in bacterial C and N incorporation and C:N coupling, across an OMZ-impacted continental margin; testing the hypotheses that bacterial activity is controlled by changes in macrofaunal biomass and the retention of labile OM within the OMZ-impacted sediments of the Indian continental margin.
Materials and methods
Study area
The Arabian Sea OMZ impinges upon the western Indian continental margin between 150 and 1500 m producing depth-dependent changes in oxygen availability, sediment geochemistry and the mega- and macrofauna (Hunter et al., 2011, 2012). The present study was carried out between September and November 2008, during R/V Yokosuka (JAMSTEC, Japan) cruise YK08-11. In situ pulse-chase experiments were conducted at four experimental stations (Figure 1) in the OMZ core (540 m) and lower OMZ boundary (800–1100 m) that differed in terms of oxygen availability (Table 1).

Bathymetric map of the experimental stations.
Culture of labelled phytodetritus
Phytodetritus was produced from an axenic clone of the diatom Thalassiosira weissflogii (NCMA, Bigelow Marine Laboratories, East Boothbay, ME, USA), cultured at 16 °C (light: dark=16:8; salinity=35; duration=28 days) in artificial seawater enriched with L1 medium (Guillard, 1975). The medium contained 99% 13C-bicarbonate (NaH13CO3, Cambridge Isotope Laboratories, Andover, MA, USA), 50% 15N-sodium nitrate (Na15NO3, Cambridge Isotope Laboratories). Algae were harvested by centrifugation (500 g; 30 min), sonicated (2000 Hz; 5 min); washed three times in ultra-pure water (Milli-Q, Merck Millipore, Billerica, MA, USA) to remove inorganic salts and dissolved organic carbon; and lyophilised (−60 °C; −0.0001 mbar; 24 h) to produce phytodetritus. Phytodetrital labelling was 27.75 atom% 13C; 33.70 atom% 15N, with a C:N mass ratio of 4.04, measured via a Flash 1112 elemental analyser linked to a Thermo Delta Plus isotope ratio mass spectrometer (EA-IRMS; Thermo Scientific, Waltham, MA, USA).
Stable-isotope labelling experiments
In situ pulse-chase experiments were conducted using Oceanlab spreader mesocosms. Each spreader consisted of a transparent polycarbonate tube (l=30 cm; d=25 cm) and acetal plastic lid, which was gently pushed into the sediment to ∼10 cm depth. The submersible then pushed a plunger through the lid to release a known dose of isotopically labelled phytodetritus onto the enclosed sediment (Figure 2). At each of the four stations, three replicate spreaders were deployed 2 m apart, by the submersible Shinkai 6500. Each spreader delivered a 650-mg C m−2, 160-mg N m−2 dose of 13C:15N-labelled phytodetritus onto the enclosed sediments. These doses were equivalent to 0.76% (±0.14%) of the 0–1 cm bulk sediment OM pool at 540 m; 0.82% (±0.22%) at 800 m; and 1.07% (±0.7%) at T2 1100 m. Lids were left in place for a minimum of 2 h, to allow phytodetritus to settle onto the sediment, and then removed by Shinkai 6500. This provides semi-enclosed meso-cosms (h=20 cm; d=25 cm), exposing each experimental treatment to the ambient conditions at each station. In contrast to enclosed incubations, DI13C and DI15N fluxes could not be quantified. However, lid-removal mitigates against experimentally-induced anoxia, which may occur in low-oxygen environments (Riedel et al., 2008). Mean current speeds were ∼0.1 m s−1 across study area (K Oguri pers. comm.), unlikely to result in sediment resuspension within the experiments. Experiments were left in situ for 4-day incubation periods. Three replicate spreaders were also deployed at stations T2 800 m and T2 1100 m for 7-day incubations. Following elapse of the incubation time each experiment was sampled using 8.2-cm-diameter push cores by Shinkai 6500, and the polycarbonate tubes recovered. Background sediment was sampled by three replicate push cores at each station. Only two spreaders could be sampled at T1 540 m, and two replicate background cores recovered at stations T1 540 m and T1 800 m. Upon recovery, cores were processed in a constant temperature room, set to sea floor ambient temperatures (Table 1). The upper 2 cm of the sediment cores were sliced into 0–1 and 1–2 cm sections, homogenised and deep frozen (−80 °C) for microbial and geochemical analysis.

Deployment of a spreader mesocosm by Shinkai 6500: (a) A spreader placed onto the sediment surface; (b) the submersible recovers the spreader lid, leaving the mesocosm in situ. All images were taken by UW during Shinkai 6500 dive SK1106 and are used courtesy of the Japan Agency Marine Science and Technology.
Analytical techniques
The particulate organic carbon, total nitrogen concentrations and isotopic labelling (13C and 15N) in the sediment bulk OM pool was measured by elemental analyser-IRMS. Before analysis, bulk sediment samples were lyophilised (−60 °C; −0.0001 mbar; 24 h), acidified with excess 1 mol l−1 hydrochloric acid (HClaq) to remove carbonates and dried to constant weight at 60 °C (following Hunter et al., 2011).
15N and 13C incorporation into the sediment HAAs and the bacterial biomarker D-alanine (D-Ala) were analysed using the method developed by Veuger et al. (2005, 2007b). Briefly, 0.2 g samples of lyophilised sediment were washed in 2 mol l−1 HClaq and Milli-Q to remove soluble material. Washed samples were hydrolysed with 6 mol l−1 HClaq (110 °C; 20 h) and purified by cation-exchange chromatography (DOWEX 50WX8-100 resin, Dow Water & Process Solutions, Midland, MI, USA). HAAs were derivatized with iso-propanol and pentafluoropropionic anhydride; further purified by solvent extraction (chloroform: phosphate buffer, 0.5: 1.0 v/v); and evaporated to dryness. Relative abundances of 15N and 13C in derivatized D-and L-amino acids and their concentrations were analysed on a Hewlett Packard 6890 gas chromatograph (Hewlett Packard, Palo Alto, CA, USA) and Thermo Delta Plus IRMS linked via a Thermo type III combustion interface (GC-c-IRMS; Thermo Scientific). HAAs were also extracted from the labelled phytodetritus in order to determine their 13C and 15N enrichment.
Phospholipid-derived fatty acids (PLFAs) were extracted from spreader and background cores to quantify bacterial biomass. PLFAs were extracted from 1.5 g of lyophilised sediment using a single-phase extraction mixture of chloroform, methanol and citrate buffer (1: 2: 0.8 v/v/v), over 2 h, and fractionated on silicic acid columns (6 ml ISOLUTE SIS PE columns, International Sorbent Technologies Ltd, Ystrad Mynach, UK) by sequential elution with chloroform (neutral lipids), acetone (glycolipids) and methanol (phospholipids). Phospholipids were transmethylated fatty acid methyl esters obtained by alkaline methanolysis, and concentrations were determined by gas chromatography flame ionisation detection (Agilent 6890N, Agilent Technologies, Santa Clara, CA, USA) (following Thornton et al., 2011).
Data treatment
Concentrations of carbon and nitrogen within sediment bulk OM and HAAs, and their isotopic ratios (13C/12C and 15N/14N) were determined by IRMS. Isotope ratios were used to calculate the atom% 13C and 15N for each sample by

where X is the 13C or 15N content (in atom%) and Rsample is the isotopic ratio of the sample. 15N and 13C enrichments of both bulk OM and HAAs, expressed per gram dry sediment, were calculated by

where E is either the concentration of excess 13C or 15N within a HAA or sediment sample; Xsample to be the atom% 13C or 15N of the sample; Xcontrol to be the natural abundance (atom%) of 13C or 15N of background HAAs or sediment OM; and Sconc. is the concentration of HAAs (in nmol g−1) or OM (in μmol g−1). 13C concentrations of the HAAs were corrected for addition of unlabelled carbon during derivatization, as described by Veuger et al. (2006). 13C and 15N enrichments of the bulk OM and HAAs were normalised to phytodetrital C (phytoC) and N (phytoN) incorporation by

where phytoE is the concentration of phytoC or phytoN, isoE is the excess 13C and 15N concentration of the sample and isoF is the 13C or 15N fraction of the phytodetritus (13C=0.2775; 15N=0.3370).
Incorporation of phytoC and phytoN into the bacterial biomarker D-Ala is expressed relative to that in the amino acid L-alanine (L-Ala), as the D/L-Ala ratio calculated by

where ID-Ala is the phytoC or phytoN incorporation by D-alanine and IL-Ala is the phytoC or phytoN incorporation by L-Ala. D-Ala data were not corrected for racemisation (formation of D-Ala from L-Ala during sample hydrolysis). Instead a racemisation background D/L-Ala ratio of 0.015 was measured for the phytodetritus (axenic T. weissflogii), and is applied in the calculation of the bacterial phytoN and phytoC incorporation as

Where bactIncorporation is the bacterial contribution (%) to label incorporation in the HAAs (%) and ExcessD/L-Ala is the D/L-Ala ratio for excess phytoC or phytoN.
D/L-Ala ratios of 0.05 and 0.1 were used as minimum and maximum estimates of the bacterial contribution (bactD/L-Ala) (Veuger et al., 2007b). These estimates are displayed graphically.
Bacterial biomass was calculated from the most common bacterial PLFAs across the four stations, i15:0, ai15:0, i16:0 and 16:1ω7c (Boschker and Middelburg, 2002), following Middelburg et al. (2000)

where Ibiomass is the bacterial biomass; IPLFA is the concentration of a bacteria-specific PLFA; a is the PLFA concentration of bacteria in hypoxic sediments (∼0.038) (Brinch-Iversen and King, 1990); and b is the fraction of bacterial PLFAs recovered from sediment samples at each station (T1 540 m, 0.28; T1 800 m, 0.25;T2 800 m, 0.22; T2 1100 m, 0.21).
Relationships between bacterial C incorporation (phytoC D/L-Ala), and changes in macrofaunal biomass and phytoC retention in the HAAs were tested for significance by robust median-based regression (Sen, 1968), using the mblm package (Komsta, 2007) in R 2.14.1 (R Development Core Team, 2009). Bacterial N incorporation data (phytoN D/L-Ala) were not amenable to statistical analysis because of missing values.
Results
Sediment geochemistry and microbial biomass
Environmental conditions at each station are described by Hunter et al. (2011) and summarised in Table 1. HAAs made up 1.3–3.0% of the sediment organic C content and 15–30% of the sediment N content, across all stations. Highest values were recorded in 0–1 cm sediment layers at T1 800 m (HAA-C∼3.1%; HAA-N∼30%). Minimal values were observed in the 1–2 cm sediment layers at T1 540 m (HAA-C∼1.3%; HAA-N∼15%). Mean bacterial biomass ranged from 19 to 72 μmol g−1. Sediment HAA concentrations and bacterial biomass exhibited high within-station variability but no trends across the study area (Table 1).
Phytodetrital C and N within sediment bulk OM and amino-acid pools
phytoC and phytoN labels within bulk OM and HAA pools were primarily restricted to the 0–1 cm sediment layers (Figure 3). Only at the core of the OMZ (T1 540 m) were phytoC and phytoN detectable within the 1−2-cm HAA pool, after 4 days (Figures 3c and d). Higher concentrations of phytoC and phytoN were retained in the bulk OM and HAAs at stations on transect 1 (T1 540 m, T1 800 m) compared with transect 2 (T2 800 m, T2 1100 m). No changes in phytoC or phytoN retention are observed in sediment bulk OM or HAAs between 4 and 7 days (Figures 3a and b).

Mean (± data ranges) phytodetrital carbon (a, c) and nitrogen (b, d) retention in the sediment bulk OM (a, b) and HAAs (c, d), over 4 and 7 days.
Bacterial incorporation of phytodetrital C and N
Incorporation of phytoC and phytoN into D-Ala were highest after 4 days at the core of the OMZ (T1 540 m): ranging from 0.036 to 0.045 in 00–01 cm sediments; and 0.039–0.048 in 01–02 cm sediments. Here, bacterial incorporation accounted for 30–90% of the phytoC and phytoN retained within the HAA pool (Figure 4). In the OMZ lower boundary (800–1100 m), phytoC and phytoN D/L-Ala ratios were close to the racemisation background, with bacterial contribution to phytoC in the total HAAs accounted for only 6–11%. Bacterial incorporation of phytoN was low across the OMZ lower boundary (800–1100 m), with the exception of one incubation at T2 800 m, where bacteria accounted for 22–54% of the phytoN in HAAs. After 7 days, D/L-Ala ratios show bacterial incorporation accounted for maxima of 28 and 22% of the phytoC in the HAA pool at T2 800 m and T2 1100 m. After 7 days, bacterial phytoN incorporation accounted for between 23% and 50% of the phytoN within the sediment HAA pool, at T2 1100 m (Figure 4).

D/L-Ala ratios and corresponding minimum and maximum estimates of contribution of bacterial assimilation to total labelled HAA pools of (a) phytodetrital carbon (phytoC in D-Ala/ phytoC in L-Ala) and (b) phytodetrital nitrogen (phytoN in D-Ala/phytoN in L-Ala). Dashed line indicates the racemisation background. Missing data points occur where phytoC or phytoN D-alanine concentrations fell below detectable thresholds.
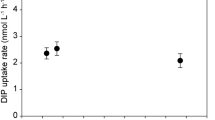
A significant negative relationship between bacterial phytoC incorporation and changes in macrofaunal biomass occurred across the study area (Figure 5). The estimated R2 value indicates that this relationship explained approximately 26% of the variation in bacterial phytoC incorporation. No relationship was observed between bacterial phytoC incorporation and retention of phytoC within the sediment HAA pool (P=0.991).

Robust linear regression of phytoC D/L-Ala ratios, in 0–1 cm sediment, against macrofaunal biomass. Dashed line indicates the racemisation background.
De-coupling of phytodetrital C and N
The phytoC:phytoN ratios of the sediment bulk OM exhibited little difference between stations and incubation times (Figure 6a). Values ranged between 8 and 12 and were consistently greater than the C:N ratio of the bulk phytodetrital material added (4.04). In the OMZ core, the phytoC:phytoN ratios of the sediment HAA pool was substantially lower than that of the added phytodetrital HAAs, revealing a preferential retention of phytoN within the HAAs (Figure 6b). phytoC:phytoN ratios were higher at the OMZ boundary stations. At oxygen concentrations ∼2 μmol l−1 phytoC:phytoN ratios ranged across the C:N ratio of the phytodetrital HAAs (16.65), with peak values of 18.8 and 21.8 after 4 days, and a value of 18.8 recorded after 7 days. At oxygen concentrations of 15 μmol l−1, phytoC:phytoN ratios were lower than the C:N ratio of the added phytodetritus, indicating enhanced phytoN retention within sediment HAAs. Between 4 and 7 days changes in phytoC:phytoN ratios at T2 1100 m, suggest a progressive loss of phytoN (Figure 6b).

phytoC:phytoN ratio of (a) the sediment bulk OM and (b) hydrolysable amino-acid pools. Dashed line indicates the C:N mass ratios of (a) the bulk phytodetrital OM and (b) the phytodetrital HAAs.
Discussion
Methodological considerations
Previous studies have used isotopic labelling of HAAs as a proxy for microbial 13C and 15N incorporation (Tobias et al., 2003; Veuger et al., 2007a), assuming sediment HAAs to account for between 50% and 60% of bacterial biomass (Cowie and Hedges, 1992). However, low oxygen mediates the preservation of biomolecules resulting in a relatively large amino-acid pool within OMZ-impacted sediments (Cowie et al., 1999; Vandewiele et al., 2009). In the present study, HAA concentrations were used as an indicator of labile OM availability and bacterial incorporation of phytoC and phytoN were traced using the bacteria-specific amino acid D-Ala. Unlike its stereoisomer L-Ala, which is present in all organisms, D-Ala is specific to bacteria. Thus, 13C and 15N labelling of D-Ala provides direct evidence of bacterial incorporation of phytoC and phytoN (Veuger et al., 2005). Ratios between D and L-Ala subsequently reflect the fraction of the total labelled HAA pool incorporated into bacterial biomass. Therefore, D/L-Ala ratios provide biomass specific indices of bacterial phytoC and phytoN (Veuger and Middelburg, 2007; Veuger et al., 2007a). While D-Ala is bacteria-specific, it also forms by the racemisation of L-Ala during the hydrolysis and extraction of amino acids. As a result, D/L-Ala ratios were plotted against a racemisation background value obtained from D/L-Ala ratio of the 13C:15N-labelled phytodetritus (following Kaiser and Benner, 2005; Veuger et al., 2007b).
It was not feasible to determine sediment OM stocks a priori, and so a fixed phytodetritus dose (650 mg C m−2) was used. Between stations, differences in relative phytodetritus dose between stations were small and unlikely to have an impact upon experimental results. The use of semi-enclosed mesocosms potentially exposed the experimental treatments to sediment resuspension and intrusion by epi-benthic megafauna. While this may introduce additional error sources into the experiments, it ensures each experiment approximates the natural disturbances within the sea-floor environmental regime. Megafaunal abundances were co-linear to macrofaunal biomass. Therefore, we cannot exclude potential roles for megafauna–macrofauna and megafauna–bacterial interactions as regulators of bacterial activity.
Phytodetrital C and N retention and bacterial assimilation across the OMZ-impacted Indian margin
In the present study, phytoC and phytoN retention within the sediment bulk OM and HAA pools changed across the study area, reflecting differences in the availability of oxygen and the presence of macrofauna and megafauna (Table 1). Sedimentary phytoC and phytoN concentrations were highest in the OMZ core, after 4 days. In this region, oxygen limitation and the absence of mega- and macrofauna are important factors promoting OM preservation (for example, Woulds et al., 2007; Jeffreys et al., 2009 and references therein). In contrast, phytoC and phytoN retention in the OMZ boundary (800–1100 m) was variable, with high phytoC and phytoN concentrations in bulk OM and HAA pools at T1 800 m, compared with the relatively low values at T2 800 m and T2 1100 m. This increased variability occured concomitantly with the presence of macrofauna (Table 1) and oxygen concentrations high enough to support nitrification but still sufficiently low to sustain denitrification and anaerobic ammonia-oxidising pathways (Devol, 1978; Jensen et al., 2011). D/L-Ala ratios demonstrate that in the OMZ core, bacterial assimilation accounted for 30–90% of the phytoC and phytoN labelling of the HAAs. This occurs concomitantly with peaks in phytoC and phytoN retention in the bulk OM and HAA pools (Figures 3 and 4), and oxygen concentrations ∼0.2% of those outside the OMZ (Hunter et al., 2011). In the OMZ boundary, bacterial phytoC and phytoN incorporation were low after 4 days, despite oxygen levels between 6 and 42 times higher than those OMZ core. However, increased phytoC and phytoN incorporation between 4 and 7 days indicates retardation of bacterial feeding activity.
Besides the difference in oxygen, there were differences in temperature between stations (Table 1). While ambient temperature is an important control on deep-sea bacterial metabolism (Moodley et al., 2005, Andersson et al., (2008) observed little change in bacterial activity over a similar temperature range at the OMZ-impacted Pakistan margin. In the present study, the primary factor controlling bacterial phytoC and phytoN incorporation appears to be oxygen availability, either directly or indirectly. A direct effect of oxygen is unlikely because anaerobic bacteria predominate within the sediments of the OMZ-impacted Indian margin (Gonsalves et al., 2011), while facultative anaerobes can sustain aerobic metabolism at oxygen concentrations ∼3 nmol l−1 (Stolper et al., 2010). Therefore, consistent with Moodley et al. (2011) and Andersson et al. (2008), we propose that oxygen availability did not directly influence bacterial activity.
Bacterial C and N incorporation were indirectly controlled by changes in oxygen availability through its influence upon benthic macrofaunal assemblages. Hunter et al. (2012) report that macrofauna consumed up to 455 μmol C m−2, and 25 μmol N m−2, after 4 days at 800 and 1100 m stations. These observations, in combination with the loss of phytoC and phytoN from the sediment HAAs indicate that fauna were active in processing phytodetrital OM. Absence of fauna at T1 540 m, however, prevent us from directly testing the effects of faunal activity upon the bacteria. However, macrofaunal biomass is strongly correlated to faunal OM uptake (for example, Middelburg et al., 2000; Woulds et al., 2007), and it is likely that differences in macrofaunal activity regulate bacterial phytoC and phytoN incorporation. Bacterial activity can also be limited by labile OM availability (Arrieta and Herndl, 2001; Nausch and Kerstan, 2001) with episodic OM depositions driving the feeding response of deep-sea sediment communities (Witte et al., 2003; Moodley et al., 2005, 2011). While bacterial activity decreased concomitantly with decreases in retention of phytoC and phytoN in the sediment HAA pools, this exhibited no significant relationship. This may be partially accounted for by high within-station variation in the sediment HAA pools and limited replication within the present study. Nevertheless, changes in bacterial feeding responses were attributed to differences in macrofaunal biomass across the OMZ. It has been shown experimentally that macrofauna feed more efficiently upon concentrated phytodetritus depositions than bacteria, making OM available to the microbial assemblages through excretion of faecal material (van Nugteren et al., 2009). Thus, stimulation of macrofaunal feeding responses may also result in retardation of phytodetrital C and N incorporation by the bacteria, consistent with increases in bacterial incorporation of phytoC and phytoN, which may be observed between 4 and 7 days.
De-coupling of phytodetrital C and N in OMZ-impacted sediments
Excess phytoC:phytoN ratios of the bulk sediment and HAAs at the four stations reveal differences in C:N coupling across the oxygen gradient (Figure 6). At the OMZ core phytoC:phytoN ratios of the sediment HAA pool were substantially lower than that of the added phytodetrital HAAs, indicating stronger phyoN retention probably because of preferential bacterial incorporation of phytoN. By contrast, phytoC:phytoN ratios of bulk sediment OM changed little between stations, indicating a preferential loss of phytoN, from the bulk sediment OM pool, at all stations. The large difference between the C:N ratios of the bulk sediment OM and added phytodetritus suggest the presence of a 15N-labelled pool of dissolved inorganic nitrogen within the isotopically labelled phytodetritus. As such, the preferential loss of phytoN from the bulk OM pool is likely to be an experimental artefact caused by DI15N leaching from phytodetrital cells.
At the OMZ core, high bacterial phytoC and phytoN incorporation, demonstrates the importance of bacteria in OM recycling pathways. Organic C is available in a variety of forms (lipids; carbohydrates; amino acids), but the main sources of organic N are amino acids. The phytoC:phytoN ratios of the HAAs at this station demonstrate preferential phytoN incorporation by the bacteria, at this station. Previous studies emphasise the importance of extracellular amino-acid oxidation as a pathway for N incorporation in marine systems (Pantoja and Lee, 1994; Mulholland et al., 2003; Veuger and Middelburg, 2007). In the extracellular amino-acid oxidation pathway bacteria liberate NH4+ from amino acids outside their cell walls (Pantoja and Lee, 1994). While we cannot determine the importance of direct and indirect N uptake pathways, extracellular amino-acid oxidation represents an efficient method for microbial N incorporation because the energetic costs of transporting NH4+ across the cell membrane are lower than those of amino acid transport (Berman et al., 1999). Thus, extracellular amino-acid oxidation represents a mechanism by which deep-sea bacteria could preferentially incorporate nitrogen from episodic OM depositions.
In the OMZ boundary, excess phytoC:phytoN ratios of the sediment HAAs reveal differences in C:N coupling linked to the changes in oxygen availability and the presence of fauna. At the low-oxygen stations (800 m, [O2]=2.20–2.36 μmol l−1) phytoC:phyto N ratios indicate reduced biological transformation of the HAAs remaining within the sediment, characterised by a weak phytoN loss. Concomitantly, the significant relationship between bacterial activity and faunal biomass, with reduced phytoC and phytoN retention within faunated sediments, suggests that the macrofauna regulate labile OM availability. Macrofaunal grazing has been observed to control the release of dissolved organic nitrogen in marine sediments (Gilbert et al., 1991) and provides pathways for OM mineralisation, to DIC and NH4+ (for example, Woulds et al., 2007, 2009). In marine systems, macrofaunal invertebrates are ammonotelic, mineralising relatively large quantities of organic N to NH4+, compared with the N retained in their body tissues (Wright, 1995). Macrofaunal feeding may provide the mechanism for the loss of phytoN loss observed at 800 m, while faunal–bacterial interactions limit bacterial activity. Potentially, this may facilitate utilisation of the phytodetrital N by heterotrophic denitrifying bacteria (Devol, 1978); and both aerobic and anaerobic ammonia-oxidising microbes (Pitcher et al., 2011), whose distributions overlap in oxygen concentrations between 1 and 3 μmol l−1. This hypothesis is supported by observations of enhanced inorganic N fluxes and nitrite production following deposition of phytodetritus at the 800 m stations (C Woulds and H Suga, personal communication).
At the higher oxygen station (1100 m; [O2]=15 μmol l−1) increased retention of phytoN within the sediment THAAs after 4 days, occurred concomitantly with low bacterial activity level. Between 4 and 7 days, net phytoN loss was observed from sediment HAAs, as bacterial phytoC and phytoN incorporation increased. This suggests that bacterial activity is retarded, where the OMZs influence upon the sediment is weaker. Lower macrofaunal biomass at this station (Table 1) may retard phytoN loss by remineralisation, potentially reducing faunal–bacterial competition for the phytodetrital N source. This would allow increased niche differentiation between macrofaunal and bacterial assemblages (van Nugteren et al., 2009). We propose that macrofaunal activity makes OM available for bacterial utilisation and provides pathway for gradual phytoN loss. This is consistent with previous observations from abyssal sediments (>4000 m), where faunal activity mediated bacterial C incorporation (Witte et al., 2003). These effects may be enhanced by megafaunal activity, with high densities present at both 800 and 1100 m. Epi-benthic megafauna are known to have an important role controlling availability of labile OM in OMZ-impacted sediments (for example, Smallwood et al., 1999; Jeffreys et al., 2009), potentially contributing to phytoN loss across study area.
Bacterial activity may also be regulated by changes in other faunal assemblages across the OMZ, such as the foraminifera and metazoan meiofauna. Both these assemblages exhibit strong zonation across OMZs, with foraminiferal distributions controlled by sediment OM availability and high foraminiferal densities observed where metazoans are absent (for example, Gooday et al., 2009). By contrast, meiofaunal densities are positively correlated to macrofauna densities, potentially relying on macrofaunal bioturbation to provide suitable microhabitats (Cook et al., 2000). Both foraminfera and meiofauna may graze upon bacteria (for example, Nomaki et al., 2006; Pascal et al., 2008). However, they may also enter into symbiotic relationships with heterotrophic bacteria; for example, in low-oxygen sediments bacterial metabolism provides an inorganic N source for nitrate-respiring foraminifera (Risgaard-Petersen et al., 2006). Interactions between the bacteria, foraminifera, and both metazoan meio- and macrofauna are potentially complex. Therefore, targeted experiments are necessary to disentangle these trophic linkages.
The present study demonstrates that macrofaunal responses to oxygen limitation regulate the heterotrophic bacterial C and N cycling pathways in marine sediments. While macrofaunal presence was linked to lower retention of phytodetrital OM and preferential loss of organic N from sediments, the relationship between bacterial phytoC incorporation and the retention of phytodetrital OM within the sediment was non-significant. We hypothesise that macrofaunal regulation of sediment bacterial activity is complex, potentially occurring via niche-partitioning and interactions with other faunal groups, such as the megafauna, foraminifera and meiofauna. OMZ expansion is likely to have implications upon sea floor biogeochemical processes, with reduced oxygen availability associated with increased nitrogen loss from the sediments. Furthermore, exclusion of fauna at extreme low-oxygen levels is associated with a shift to OM processing by heterotrophic bacteria and retention of organic N within sediments. These hypotheses require testing over longer time periods within a manipulative experimental framework, to further explore faunal–bacterial interactions.
References
Andersson JH, Woulds C, Schwartz M, Cowie GL, Levin LA, Soetaert K et al. (2008). Short-term fate of phytodetritus in sediments across the Arabian Sea Oxygen Minimum Zone. Biogeosciences 5: 43–53.
Arrieta JM, Herndl GL . (2001). Changes in bacterial beta-gucosidase diversity during a coastal phytoplankton bloom. Limnol Oceanogr 47: 594–599.
Berman T, Bechemin C, Maestrini SY . (1999). Release of ammonium and urea from dissolved organic nitrogen in aquatic ecosystems. Aq Micr Ecol 16: 295–302.
Billett DSM, Lampitt RS, Rice AL, Mantoura RFC . (1983). Seasonal sedimentation of phytoplankton to the deep-sea benthos. Nature 302: 520–522.
Boschker HTS, Middelburg JJ . (2002). Stable isotopes and biomarkers in microbial ecology. FEMS Microbiol Ecol 40: 85–95.
Brinch-Iversen J, King GM . (1990). Effects of substrate concentration, growth state, and oxygen availability on relationships among bacterial carbon, nitrogen and phospholipid phosphorus content. FEMS Microbiol Lett 74: 345–355.
Buhring SI, Lampadariou N, Moodley L, Tselepides A, Witte U . (2006). Benthic microbial and whole-community responses to different amounts of C-13-enriched algae: In situ experiments in the deep Cretan Sea (Eastern Mediterranean). Limnol Oceanogr 51: 157–165.
Cook AA, Lambshead PJD, Hawkins LE, Mitchell N, Levin LA . (2000). Nematode abundance at the oxygen minimum zone in the Arabian Sea. Deep Sea Res Pt II 47: 75–85.
Cook PLM, Veuger B, Boer S, Middelburg JJ . (2007). Effect of nutrient availability on carbon and nitrogen incorporation and flows through benthic algae and bacteria in near-shore sandy sediment. Aq Micr Ecol 49: 165–180.
Cowie GL, Calvert SE, Pedersen TF, Schulz H, Von Rad U . (1999). Organic content and preservational controls in surficial shelf and slope sediments from the Arabian Sea (Pakistan margin). Mar Geol 161: 23–38.
Cowie GL, Hedges JI . (1992). Sources and reactivities of amino acids in a coastal marine environment. Limnol Oceanogr 37: 703–724.
Dauwe B, Middelburg JJ . (1998). Amino acids and hexosamines as indicators of organic matter degradation state in North Sea sediments. Limnol Oceanogr 43: 782–798.
Devol AH . (1978). Bacterial oxygen uptake kinetics as related to biological processes in the oxygen deficient zones of the oceans. Deep Sea Res 25: 137–146.
Evrard V, Cook PLM, Veuger B, Huettel M, Middelburg JJ . (2008). Tracing carbon and nitrogen incorporation and pathways in the microbial community of a photic subtidal sand. Aquat Microb Ecol 53: 257–269.
Francis CA, Beman JM, Kuypers MMM . (2007). New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation. ISME Journal 1: 19–27.
Gilbert PM, Garside C, Fuhrman JA, Roman MR . (1991). Time-dependent coupling of inorganic and organic nitrogen uptake and regeneration in the plume of the Chesapeake Bay estuary and its regulation by large heterotrophs. Limnol Oceanogr 36: 895–909.
Glud RN, Gundersen JK, Jorgensen BB, Revesbech NP, Schulz HD . (1994). Diffusive and total oxygen uptake of deep-sea sediments in the eastern South Atlantic Ocean: In situ and laboratory measurements. Deep Sea Res Pt I 41: 1767–1788.
Gooday AJ, Levin LA, Aranda da Silva A, Bett BJ, Cowie GL et al. (2009). Faunal responses to oxygen gradients on the Pakistan Margin: a comparison of foraminiferans, macrofauna and megafauna. Deep Sea Res Pt II 56: 488–502.
Gonsalves MJ, Paropkari AL, Fernandes CEG, Loka Bharathi PA, Krishnakumari L, Fernando V et al. (2011). Predominance of anaerobic bacterial community over aerobic community contribute to intensify ‘oxygen minimum zone’ in the eastern Arabian Sea. Cont Shelf Res 31: 1224–1235.
Guillard RRL . (1975). Culture of phytoplankton for feeding marine invertebrates. In: Smith WL, Chanley MH (eds). Culture of Marine Invertebrate Animals. Plenum: New York, USA, pp 26–60.
Hall POJ, Brunnegard J, Hulthe G, Martin WR, Stahl H, Tengberg A . (2007). Dissolved organic matter in abyssal sediments: core recovery artifacts. Limnol Oceanogr 52: 19–31.
Helly JJ, Levin LA . (2004). Global distribution of naturally occurring marine hypoxia on continental margins. Deep Sea Res Pt I 51: 1159–1168.
Hunter WR, Levin LA, Kitazato H, Witte U . (2012). Macrobenthic assemblage structure and organismal stoichiometry control faunal uptake of phytodetritus in OMZ-impacted sediments. Biogeosci 9: 993–1006.
Hunter WR, Oguri K, Kitazato H, Ansari ZA, Witte U . (2011). Epi-benthic megafaunal zonation across an oxygen minimum zone at the Indian continental margin. Deep Sea Res Pt I 58: 699–710.
Jeffreys RM, Wolff GA, Cowie GL . (2009). Influence of oxygen on heterotrophic reworking of sedimentary lipids at the Pakistan Margin. Deep Sea Res Pt II 56: 358–375.
Jensen MM, Lam P, Revsbech NP, Nagel B, Gaye B, Jetten MSM et al. (2011). Intensive nitrogen loss over the Omani Shelf due to anammox coupled with dissimilatory nitrite reduction to ammonium. ISME Journal 5: 1660–1670.
Kaiser K, Benner R . (2005). Hydrolysis-induced racemization of amino acids. Limnol Oceanogr Meth 3: 318–325.
Komsta L . (2007) Median-based Linear Models 0.11.
Middelburg JJ, Barranguet C, Boschker HTS, Herman PMJ, Moens T, Heip CHR . (2000). The fate of intertidal microphytobenthos carbon: An in situ13C-labeling study. Limnol Oceanogr 45: 1224–1234.
Middelburg JJ, Soetaert K, Herman PMJ . (1997). Empirical relationships for use in global diagenetic models. Deep Sea Res Pt I 44: 327–344.
Moodley L, Middelburg JJ, Soetaert K, Boschker HTS, Herman PMJ, Heip CHR . (2005). Similar rapid response to phytodetritus deposition in shallow and deep-sea sediments. J Mar Res 63: 457–469.
Moodley L, Nigam R, Ingole B, Prakash Babu C, Panchang R, Nanajkar M et al. (2011). Oxygen minimum seafloor ecological (mal) functioning. J Exp Mar Biol Ecol 398: 91–100.
Mulholland MR, Lee C, Glibert PM . (2003). Extracellular enzyme activity and uptake of carbon and nitrogen along an estuarine salinity and nutrient gradient. Mar Ecol Prog Ser 258: 3–17.
Nausch M, Kerstan E . (2001). The relationship between dissolved carbohydrates and carbohydrate-degrading enzymes in the salinity gradient of the Pomeranian Bight (southern Baltic). Oceanol 45: 437–452.
Nomaki H, Heinz P, Nakatsuka T, Shimanaga M, Ohkouchi N, Ogawa NO et al. (2006). Different ingestion pattern of 13C-labeled bacteria and algae by deep-sea benthic foraminifera. Mar Ecol Prog Ser 310: 95–108.
Pantoja S, Lee C . (1994). Cell-surface oxidation of amino acids in seawater. Limnol Oceanogr 39: 1718–1726.
Pascal P-Y, Dupuy C, Richard P, Rzeznik-Orignac J, Nipuil N . (2008). Bacterivory of a mudflat nematode community under different environmental conditions. Mar Biol 154: 671–682.
Pitcher A, Villanueva L, Hopmans EC, Schouten S, Reichart GJ, Damste JSS . (2011). Niche segregation of ammonia-oxidising archaea and anammox bacteria in the Arabian Sea oxygen minimum zone. ISME J 5: 1896–1904.
R Development Core Team. (2009) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria.
Riedel B, Zuschin M, Haselmair A, Stachowitsch M . (2008). Oxygen depletion under glass: Behavioural responses of benthic macrofauna to induced anoxia in the Northern Adriatic. J Exp Mar Biol Ecol 367: 17–27.
Risgaard-Petersen N, Langezaal AM, Ingvardsen S, Schmid MC, Jetten MSM, Op den Camp HJM et al. (2006). Evidence for complete denitrification in a benthic foraminifer. Nature 443: 93–96.
Sen PK . (1968). Estimates of regression coefficient based on Kendall's tau. J Am Stat Ass 63: 1379–1389.
Schwartz MC, Woulds C, Cowie GL . (2009). Sedimentary denitrification rates across the Arabian Sea oxygen minimum zone. Deep Sea Res Pt II 56: 324–332.
Smallwood BJ, Wolff GA, Bett BJ, Smith CR, Hoover D, Gage JD et al. (1999). Megafauna can control the quality of organic matter in marine sediments. Naturwiss 86: 320–324.
Stolper DA, Revsbech NP, Canfield DE . (2010). Aerobic growth at nanomolar oxygen concentrations. Proc Natl Acad Sci 107: 18755–18760.
Stramma L, Schmidtko S, Levin LA, Johnson GC . (2010). Ocean oxygen minima expansions and their biological impacts. Deep Sea Res Pt I 57: 587–595.
Thingstad TF . (1987). Utilization of N, P and organic C by heterotrophic bacteria. I. Outline of a chemostat theory with a consistent concept of ‘maintenance’ metabolism. Mar Ecol Prog Ser 35: 99–109.
Thornton B, Zhang Z, Mayes RW, Hogberg MN, Millwood AJ . (2011). Can gas chromatography combustion isotope ratio mass spectrometry be used to quantify organic compound abundance? Rapid Commun Mass Spectrom 25: 2433–2438.
Tobias C, Giblin A, McClelland J, Tucker J, Peterson B . (2003). Sediment DIN fluxes and preferential recycling of benthic microalgal nitrogen in a shallow macrotidal estuary. Mar Ecol Prog Ser 257: 25–36.
van Nugteren P, Herman PMJ, Moodley L, Middelburg JJ, Vos M, Heip CHR . (2009). Spatial distribution of detrital resources determines the outcome of competition between bacteria and a facultative detritivorous worm. Limnol Oceanogr 54: 1413–1419.
Vandewiele S, Cowie G, Soetaert K, Middelburg JJ . (2009). Amino acid biogeochemistry and organic matter degradation state across the Pakistan margin oxygen minimum zone. Deep Sea Res Pt II 56: 318–334.
Veuger B, Eyre BD, Maher D, Middelburg JJ . (2007a). Nitrogen incorporation and retention by bacteria, algae, and fauna in a subtropical intertidal sediment: an in situ15N-labeling study. Limnol Oceanogr 52: 1930–1942.
Veuger B, Middelburg JJ . (2007). Incorporation of nitrogen from amino acids and urea by benthic microbes: role of bacteria versus algae and coupled incorporation of carbon. Aquat Microb Ecol 48: 35–46.
Veuger B, Middelburg JJ, Boschker HTS, Houtekamer M . (2005). Analysis of 15N incorporation into D-alanine: A new method for tracing nitrogen uptake by bacteria. Limnol Oceanogr: Meth 3: 230–240.
Veuger B, Middelburg JJ, Boschker HTS, Houtekamer M . (2007b). Update of ‘Analysis of 15N incorporation into D-alanine: A new method for tracing nitrogen uptake by bacteria’ (Veuger et al., 2005, Limnol. Oceanogr. Methods 3: 230–240). Limnol Oceanogr Meth 5: 192–194.
Veuger B, Van Oevelen D, Boschker HTS, Middelburg JJ . (2006). Fate of peptidoglycan in an intertidal sediment: an in situ13C-labeling study. Limnol Oceanogr 51: 1572–1580.
Ward BB, Devol AH, Rich JJ, Chang BX, Bulow SE, Naik H et al. (2009). Denitrification as the dominant nitrogen loss process in the Arabian Sea. Nature 461: 78–81.
Witte U, Wenzhofer F, Sommer S, Boetius A, Heinz P, Aberle N et al. (2003). In situ experimental evidence of the fate of a phytodetritus pulse at the abyssal sea floor. Nature 424: 763–766.
Woulds C, Cowie GL, Levin LA, Andersson JH, Middelburg JJ, Vandewiele S et al. (2007). Oxygen as a control on seafloor biological communities and their roles in sedimentary carbon cycling. Limnol Oceanogr 52: 1698–1709.
Woulds C, Schwartz MC, Brand T, Cowie GL, Law G, Mowbray SR . (2009). Porewater nutrient concentrations and benthic nutrient fluxes across the Pakistan margin OMZ. Deep Sea Res Pt II 56: 333–346.
Wright PA . (1995). Nitrogen excretion: three end products, many physiological roles. J Exp Biol 198: 273–281.
Acknowledgements
We thank the editor and anonymous referees for an efficient and constructive review. We are deeply indebted to Professor Hiroshi Kitazato (JAMSTEC, Japan), the officers and crew of the R/V Yokosuka and Shinkai 6500, and the team of scientists during cruise YK08-11. Without their dedication and support, this study could not have been conducted. We acknowledge invaluable pre- and post-cruise logistical support by Professor Waji Naqvi (NIO, India), Dr Hidetaka Nomaki (JAMSTEC, Japan) and Dr Markus Moeseneder (NOCS, UK). This study could not have been completed without the help of Marco Houtekamer, Peter van Breugel and Pieter van Rijswijk at NIOZ, and Barry Thornton and Maureen Procee at the James Hutton Institute. The project was supported by the Carnegie Trust (United Kingdom) (Grant no. ERI 008427 to U Witte), WRH is funded by a NERC Doctoral Training Grant (NE/G523904/1). Analytical work for this paper was supported by a Travel Grant from the Challenger Society for Marine Science to WRH. BV was financially supported by the Netherlands Organization for Scientific Research (Pionier 833.02.2002) and the Darwin Center for Biogeology (project 142.16.1052).
Author contributions
Experiments were designed by UW, and conducted by UW and WRH. Analytical work was carried out by WRH and BV. WRH, BV and UW all contributed to the writing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Hunter, W., Veuger, B. & Witte, U. Macrofauna regulate heterotrophic bacterial carbon and nitrogen incorporation in low-oxygen sediments. ISME J 6, 2140–2151 (2012). https://doi.org/10.1038/ismej.2012.44
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2012.44
Keywords
This article is cited by
-
17α-ethynylestradiol (EE2) limits the impact of ibuprofen upon respiration by streambed biofilms in a sub-urban stream
Environmental Science and Pollution Research (2020)
-
On a non-parametric confidence interval for the regression slope
METRON (2017)
