
Abstract
Death receptors, including the TNF receptor-1 (TNF-RI), have been shown to be able to initiate caspase-independent cell death. This form of “necrotic cell death” appears to be dependent on the generation of reactive oxygen species. Recent data have indicated that superoxide generation is dependent on the activation of NADPH oxidases, which form a complex with the adaptor molecules RIP1 and TRADD. The mechanism of superoxide generation further establishes RIP1 as the central molecule in ROS production and cell death initiated by TNFα and other death receptors. A role for the sustained JNK activation in necrotic cell death is also suggested. The sensitization of virus-infected cells to TNFα indicates that necrotic cell death may represent an alternative cell death pathway for clearance of infected cells.
Similar content being viewed by others
ROS and death receptors
Levels of cellular reactive oxygen species (ROS) are modulated by the balance between the cellular processes that produce ROS and the processes that eliminate them 1. In most situations, ROS are attenuated by systems of antioxidant enzymes, including superoxide dismutases, catalase, thioredoxin reductase, glutathione peroxidases, and peroxiredoxins, as well as non-enzymatic antioxidants 1, 2. ROS are produced primarily by three sources within the cell. First, and foremost in ROS production is the mitochondria, from which some electrons leak at a rate that has been estimated to be between 2 to 5% as they are transferred between electron transport chain complexes 3, 4. A second major source of ROS in the cell are the NADPH oxidase complexes, which use NADPH as a substrate to bring about the direct reduction of molecular oxygen to produce superoxide anions 5 which play an important role in defense against environmental pathogens 6 as well as in signal transduction pathways. Third, many other enzymes produce ROS in many different cell types, although less robustly, including xanthine oxidase, cyclooxygenases, lipoxygenases, myeloperoxidases, heme oxygenase, monoamine oxidases, and aldehyde oxidase, as well as cytochrome P450-based enzymes. Cytokines and growth factors that bind to receptors of different classes have also been reported to lead to the generation of ROS that function as second messengers in their signal transduction pathways 7, 8, however, the production of ROS may not be intrinsic to the cytokines and growth factors and may actually be achieved by the activation of other enzymes, such as NADPH oxidases. This appears to be the case in signaling and death induced by death receptors.
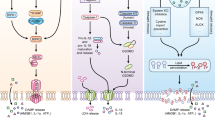
The death receptors are a subfamily of receptors in the TNF Receptor superfamily (for reviews, 9, 10). These receptors contain an intracellular “death domain” through which they mediate their downstream signaling pathways by means of homotypic interactions with death domain-containing adaptor proteins, such as FADD, TRADD, and RIP1. These death receptors, including TNF-R1 (Figure 1), Fas, Death Receptor 3, and the TRAIL receptors, DR4 and DR5, induce apoptosis in many cell types through FADD-dependent activation of caspase-8. When activated, caspase-8 may act in the apoptotic process by directly cleaving cellular proteins, or may act indirectly to induce apoptosis through cleavage of the BH3-only protein BID. Truncated Bid protein acts on the mitochondria to cause the release of cytochrome c from the mitochondria, which activates further downstream caspases through an Apaf-1/caspase-9 mediated pathway.

(A) TNFα signaling proceeds by the binding of the TNFα trimer to the TNF-R1 receptor. This initiates a change in the receptor that brings about the binding of the TRADD and RIP1 adaptor proteins to the death domain of the receptor. These adaptor proteins are then able to recruit TRAF2 and the IKK machinery (not shown) to the complex. (B) Recruitment of TRADD, RIP1, TRAF2, and the downstream machinery causes the degradation of the IκBα protein and activation of MAP kinase kinase kinases (MAP3Ks). The proteolytic degradation of IκBα, which typically retains NF-κB in the cytoplasm, allows NF-κB to translocate to the nucleus where it acts as a transcription factor. The MAP3Ks phosphorylate and activate downstream kinases that result in the activation of the MAP Kinases, JNK, p38 and ERK. Activation of these kinases in combination with NF-κB activation results in the transcription of pro-inflammatory and pro-survival genes. (C) When NF-κB activation is diminished or prevented, TNFα signaling results in the recruitment of FADD and caspase-8 to a secondary complex that triggers caspase-8 activation, which leads to apoptosis. One of the substrates for caspase-8 is RIP1. Cleavage of RIP1 further potentiates apoptosis. (D) When caspases are inhibited, apoptosis is prevented and the RIP1 protein is stabilized. RIP1 and TRADD form a complex with the NOXO1, which recruits Nox1 and Rac1 to form an active superoxide producing complex. RIP1 is proposed to be important for NOXO1 recruitment, while TRADD may be important in the activation of the complex through its interaction with the SH3 domain of NOXO1. The presence of NOXA1 as an activator is predicted, but has not yet been shown. The presence of p22 in the complex is unknown. Production of superoxide is proposed to promote the sustained activation of JNK, which leads to necrotic cell death.
While the pathways leading to caspase-dependent apoptosis are reasonably well understood, death receptors also have been shown under some circumstances to initiate cell death through less-defined pathways that are not dependent on caspase activity 11. In the case of the TRAIL receptors, caspase-independent cell death has been reported to take the form of autophagic cell death 12, 13, a variant of programmed cell death that requires specific genetically defined pathways and which occurs following a failed attempt by the cell to survive by meeting its energetic needs through the lysosomal degradation of cellular proteins and/or organelles 14, 15. In most cases, however, caspase-independent cell death induced by death receptors does not appear to be the result of autophagy, as autophagy inhibitors such as 3-methyladenine, chloroquine, and PI 3-kinase inhibitors, or energy supplementation with methyl pyruvate fail to inhibit cell death. The morphology of this form of death resembles that of necrosis, with organelle and cellular swelling, large-scale mitochondrial damage and loss of membrane integrity 15, 16. Therefore, we (and others in the field) have referred to caspase-independent cell death induced by death receptors as “necrotic cell death”, although this is clearly not the passive, non-programmed process that necrosis is usually depicted as 17. Although necrosis has been described classically as accidental cell death occurring only in cases of severe pathological damage, there is now evidence that necrotic cell death may also play a role in normal physiology and development 11, 14.
Necrotic cell death induced by death receptors has been proposed to involve the generation of ROS derived from either mitochondrial or non-mitochondrial sources 14, 16. Caspase inhibition under these circumstances actually leads to an enhancement of cell death, rather than its inhibition 18. This may be due, in part, to caspase-8 cleavage of RIP1 during the apoptotic process (Figure 1C, 19), and may imply a certain level of basal cleavage of RIP1 in some cell types 20. From the data of many groups, RIP1 appears to be a central molecule in the initiation of caspase-independent cell death by death receptors, as well as by other stimuli 11, 18, 21, 22. The RIP1 protein is necessary for the generation of ROS by TNFα and is required for the initiation of caspase-independent cell death 22, 23.
FasL, TRAIL, and TNFα have been shown to lead to ROS generation in response to receptor stimulation 16, 24, 25, 26, 27, 28. The production of ROS in many cases has been implicated to come from downstream events involving mitochondria 16. It is now becoming clear, however, that both TNFα and FasL can stimulate the production of superoxide by activating NADPH oxidases 25, 29, 30, 31.
NADPH oxidases
NADPH oxidases are one of the few enzymes specifically dedicated to the production of ROS. Originally thought to be a single enzyme, NADPH oxidases have now been found to comprise a large family of enzymes, with the expression of different isoforms being tissue specific 5. Activated neutrophils and macrophages produce large quantities of superoxide for host defense via the phagocytic form of NADPH oxidase, Nox2 (previously known as gp91phox). A number of other NADPH oxidase family members (Nox1, Nox3, and Nox4, Nox5, DUOX1, DUOX 2) have been characterized 5 in non-phagocytic cell types, including fibroblasts, vascular smooth muscle cells, cardiac myocytes, and endothelial cells. These cell types produce small amounts of ROS by NADPH oxidases to regulate intracellular signaling cascades 5, 6. NOXs 1-4 exist as heterodimers with a 22 kDa subunit (p22phox), and require the presence of this small subunit for activity. In the case of Nox2, two other activator subunits are required for activity. Upon phosphorylation in response to stimulus, the p47phox subunit binds to membrane phospholipids, interacts with p22phox, and recruits the p67phox subunit to the complex 5. The p67phox activator binds and stabilizes an interaction of the complex with the small GTPase Rac (Rac1 or Rac2, depending on cell type), resulting in an active enzyme complex 5. The regulatory p41NOXO1 and p51NOXA1 subunits may function in other oxidase complexes, such as Nox1, similarly to p47phox and p67phox, respectively, with some small differences 32, 33, 34. NOXO1, unlike p47phox, lacks an autoinhibitory region, and does not require phosphorylation for membrane translocation. Hence it requires no new activation stimulus for activity. Its PX domain also has a slightly different lipid preference 35. NOXA1, unlike p67phox, has a single SH3 domain rather than two SH3 domains. In overexpression studies, when p47phox and/or p67phox were exchanged for NOXO1 and/or NOXA1 and vice-versa in the Nox1 and Nox2 complexes, the complexes were still functional, though the superoxide generation was less potent 32, 33, 34. It is unclear as to whether this actually occurs in endogenous complexes, but, if true, it may indicate that there is some redundancy in the use of subunits by Nox enzymes, or that the amount of superoxide produced may be modulated by subunit expression in a cell type specific manner.
TNFα has long been known to potentiate NADPH oxidase activity in macrophages, monocytes, and neutrophils, however, a second stimulus with another agonist, such as PMA was considered necessary to actually activate the oxidase. While it has been clear that TNFα is a powerful enabler of Nox2 activation, the mechanism by which this occurs is uncertain. Several reports have shown that TNFα leads to increased expression of various NADPH oxidase components 36, 37, 38, and this appears to contribute to increased oxidase activity, especially in the long term. However, more direct potential mechanisms for potentiation of superoxide production have been proposed, some of which involve the phosphorylation of p47phox by different kinases, including PKCζ 39, a tyrosine kinase 40, 41, 42, or p38 MAPK 43. TNFα has also been proposed to regulate Nox2 activity though regulation of the proposed associated hydrogen ion channel that balances the electronic charge translocated by NADPH oxidase 44.
In contrast to the indirect activation of Nox2, we have recently shown that TNFα is a direct activator of the Nox1 NADPH oxidase in the L929 murine fibrosarcoma cell line, as well as in p65−/− MEFs subjected to caspase inhibition 31. Treatment of L929 cells with TNFα alone is sufficient to induce superoxide production within 30-45 minutes of treatment, and the superoxide generated by Nox1 contributes substantially to TNFα-induced necrotic cell death. Knockdown of Nox1 with siRNA prevents both superoxide generation and cell death in response to TNFα. Therefore, Nox1 appears to be the primary source of ROS involved in TNFα-induced cell death in L929 cells.
Nox1, NOXO1 and Rac1 form a signaling complex with components of the TNF receptor complex in a TNF-dependent manner 31. The mechanism of activation of Nox1 by TNFα in these cells is suggested by protein interactions between the death domain-containing TNF-R1 adaptor proteins RIP1 and TRADD, and the Nox1 organizer protein, NOXO1 (Figure 1D). RIP1 interacts strongly with NOXO1 protein through an as yet undefined domain, while the polyproline-rich region of TRADD interacts weakly with the SH3 domain of NOXO1. RIP1 is required for formation of the induced signaling complex, as Nox1 fails to immunoprecipitate with TRADD in RIP−/− MEFs under TNFα-mediated necrotic conditions. Overexpression of a mutant TRADD protein with a mutation in its proline region that abrogates binding to NOXO1 diminishes superoxide formation and cell death in response to TNFα, and thus the mutant protein functions as a dominant negative molecule with regard to Nox1 activation, without affecting other TNFα signaling pathways. Based on the relative affinities of RIP1 and TRADD for NOXO1, we have proposed a model in which RIP1 recruits NOXO1 and Nox1/NOXO1/Rac1 to the complex, where a second, lower affinity interaction, between NOXO1 and TRADD is required to promote oxidase activation 31.
Consistent with the hypothesis that NOXO1 but not p47phox was involved in oxidase activation, we found that the tyrosine kinase inhibitor genestein, the general PKC inhibitor, bisindolymaleimide, or a specific PKCζ peptide inhibitor had no effect on the superoxide production in L929 cells in response to TNFα. Therefore it appears that induction of this protein complex is a completely novel mechanism by which TNFα affects oxidase activation.
Although previous work has established a role for ROS in the pathway of TNFα–induced necrotic cell death 23, 45, 46, 47, 48, some previous data have suggested that the process involves mitochondrial derived ROS 16, 45, 49, 50. While we have established that the non-mitochondrial oxidase Nox1 appears to be involved, the source of ROS in TNF-stimulated cell death is by no means exclusive. In other experiments, we have detected a large amount of what appears to be mitochondrial-derived ROS generated several hours after the addition of TNFα, and at a much later time point than the earliest appearance of superoxide (unpublished data). While this could mark the appearance of non-specific ROS as a result of the beginning of cell death, it could also suggest the participation of both Nox1-produced superoxide and mitochondrial-produced ROS in the process. As the mechanism of production of mitochondrial-derived ROS has not yet been established, it may be that the superoxide produced by non-mitochondrial oxidases leads to the generation of mitochondrial-derived ROS. Oxidative radical stress itself is known to further amplify the production of ROS within the mitochondria through protein damage 51, which may act as a positive feedback loop. One other group has suggested that Nox1 plays a role in necrotic cell death induced by serum withdrawal. In this case, they propose that Nox1 activation is actually downstream of the production of mitochondrial ROS 52. Though we cannot at this time eliminate this possibility, the kinetics of complex formation suggests to us that the Nox1-mediated superoxide generation is an early event.
Contribution of ROS to cell death
How do the superoxide and other ROS generated downstream of TNF stimulation contribute to cell death? The molecular targets of ROS have begun to be defined, but much is not yet known. ROS production may have various roles in cell death, namely, ROS may directly oxidize cellular proteins, lipids, or nucleic acids and cause general damage and dysfunction, or ROS may initiate cell death processes through affecting various signaling cascades 17. One of the major ways by which ROS affect signaling pathways is through the direct reaction of these compounds with the catalytic sites of phosphatases. Classical protein tyrosine phosphatases have long been known to be inactivated by ROS-mediated oxidation of their catalytic cysteine 53, 54. More recent data suggest that ROS are also capable of inactivating dual specificity phosphatases 55, which can dephosphorylate tyrosine and serine/threonine residues, as well as phospholipids. Depending on the oxidation state of the catalytic cysteine, inactivation of phosphatases by ROS may be reversible or irreversible 54, 56. Having a prolonged phosphorylation status affects the activity of many proteins within the cell, including the stress-activated MAP kinases, p38 and JNK 55.
JNK activity has been reported to be positively regulated by ROS in a number of ways. Primarily, JNK is positively affected by the direct inactivation of JNK phosphatase activity by ROS 55. The monomeric form of glutathione S-transferase Pi (GSTπ) binds directly to the C-terminus of JNK and inhibits its activation 57. ROS induces oligomerization of GSTπ and causes its dissociation from JNK, resulting in JNK activation 58. ASK1 is a MAP3K that has been reported to be important in promoting downstream sustained phosphorylation of JNK under conditions of oxidative stress 59. The reduced form of thioredoxin binds to ASK1 and blocks its kinase activity in a large complex, which has been termed the ASK1 signalosome 60, 61, 62. Upon oxidation, thioredoxin is released and TRAF2 and/or TRAF6 are recruited to the signalosome along with downstream kinases, which then are involved in the activation of JNK 62.
JNK has many different functions in cell death 63. Sustained JNK activation is seen in L929 cells after TNF treatment, which appears to correlate with superoxide generation and necrotic cell death 31. Suppression of JNK by genetic or pharmacological approaches offers significant protection against cell death induced by ROS and TNF 64, and inhibition of JNK completely blocks TNF-induced necrotic cell death in L929 cells 31. This is consistent with the finding that exogenously applied ROS induce necrotic cell death via JNK activation 21. Therefore, JNK is proposed as a critical mediator of necrotic cell death 64, and sustained JNK activation is thought to be one of the key events in necrotic cell death induced by TNF-α 31, 47, 55.
Physiological relevancy of TNF-induced necrotic death
In most cell types under physiological conditions, TNFα treatment does not result in cell death (Figure 1B). This is due in large measure to the activation of NF-κB, which stimulates the transcription of genes for pro-survival proteins 65, such as the caspase-8 inhibitory protein cFLIP, the cytoplasmic zinc finger protein A20, the antiapoptotic Bcl-2 family protein A1, and the cellular inhibitor of apoptosis proteins (cIAPs). Other NF-κB regulated genes have been reported to affect JNK activation, such as GADD45β and XIAP 66, 67. TNF-α-induced NF-κB activation can reduce ROS levels via increased expression of the antioxidant proteins FHC and MnSOD 68, 69. Thus, NF-κB prevents both apoptosis and sustained JNK activation 70, 71, 72, 73. Therefore, under normal conditions, both apoptotic and necrotic cell death are inhibited; and under most circumstances where NF-κB is inhibited, apoptosis generally prevails due to the activation of caspases (Figure 1C). Under what conditions, then, does TNFα initiate necrotic cell death? The key to this question may lie in the increased sensitization of cells to necrotic cell death in the presence of caspase-inhibitors, as it has long been known that many pathogens encode proteins that inhibit caspase activation. They consequently prevent apoptosis of an infected cell in response to TNFα, which, as an inflammatory cytokine, would be present at the highest amounts during an infection. Thus, it has been proposed that necrotic cell death may represent an alternative pathway to eliminate infected cells from the body 11. This hypothesis is borne out by the observation in several cases that cells infected with virus are actually sensitized to TNFα-induced killing, and that viruses lacking the caspase-inhibitory proteins do not lead to sensitization 74, 75. RIP1 deficiency, however, rescued infected cells from TNFα-induced cytotoxicity 75. These reports do suggest that necrotic cell death induced by TNFα is likely to be physiologically relevant under conditions of infection and that RIP1 is a central molecule in initiation of the TNFα-induced necrotic cell death pathway.
Conclusion
In conclusion, TNFα induces a necrotic-like cell death when caspases are inhibited within the cell. This death is mediated, at least in part, by ROS generated by the activation of the NADPH oxidase, Nox1, through complex formation between Nox1 subunits and the TNF receptor adaptors RIP1 and TRADD 31. The model we have proposed (Figure 1D) is that RIP1 recruits NOXO1 and Nox1/NOXO1/Rac1 to a complex where TRADD is required to promote oxidase activation. ROS likely mediate some of their downstream effects on necrotic cell death through the sustained activation of JNK. TNFα-induced necrotic cell death is most likely to play a physiological role in the clearance of infected cells during an immune response.
References
Sies H . Oxidative stress: oxidants and antioxidants. Exp Physiol 1997; 82:291–295.
Rhee SG, Yang KS, Kang SW, Woo HA, Chang TS . Controlled elimination of intracellular H(2)O(2): regulation of peroxiredoxin, catalase, and glutathione peroxidase via post-translational modification. Antioxid Redox Signal 2005; 7:619–626.
Boveris A, Chance B . The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem J 1973; 134:707–716.
Boveris A, Cadenas E . Mitochondrial production of superoxide anions and its relationship to the antimycin insensitive respiration. FEBS Lett 1975; 54:311–314.
Lambeth JD . NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol 2004; 4:181–189.
Quinn MT, Ammons MC, Deleo FR . The expanding role of NADPH oxidases in health and disease: no longer just agents of death and destruction. Clin Sci (Lond) 2006; 111:1–20.
Sauer H, Wartenberg M, Hescheler J . Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell Physiol Biochem 2001; 11:173–186.
Shen HM, Pervaiz S . TNF receptor superfamily-induced cell death: redox-dependent execution. Faseb J 2006; 20:1589–1598.
Wajant H . Death receptors. Essays Biochem 2003; 39:53–71.
Thorburn A . Death receptor-induced cell killing. Cell Signal 2004; 16:139–144.
Festjens N, Vanden Berghe T, Vandenabeele P . Necrosis, a well-orchestrated form of cell demise: signalling cascades, important mediators and concomitant immune response. Biochim Biophys Acta 2006; 1757:1371–1387.
Thorburn J, Moore F, Rao A, et al. Selective inactivation of a Fas-associated death domain protein (FADD)-dependent apoptosis and autophagy pathway in immortal epithelial cells. Mol Biol Cell 2005; 16:1189–1199.
Park KJ, Lee SH, Kim TI, et al. A human scFv antibody against TRAIL receptor 2 induces autophagic cell death in both TRAIL-sensitive and TRAIL-resistant cancer cells. Cancer Res 2007; 67:7327–7334.
Edinger AL, Thompson CB . Death by design: apoptosis, necrosis and autophagy. Curr Opin Cell Biol 2004; 16:663–669.
Kroemer G, El-Deiry WS, Golstein P, et al. Classification of cell death: recommendations of the Nomenclature Committee on Cell Death. Cell Death Differ 2005; 12 Suppl 2:1463–1467.
Fiers W, Beyaert R, Declercq W, Vandenabeele P . More than one way to die: apoptosis, necrosis and reactive oxygen damage. Oncogene 1999; 18:7719–7730.
Morgan MJ, Kim YS, Liu Z . Lipid rafts and oxidative stress-induced cell death. Antioxid Redox Signal 2007; 9:1471–1483.
Vandenabeele P, Vanden Berghe T, Festjens N . Caspase inhibitors promote alternative cell death pathways. Sci STKE 2006; pe44:.
Lin Y, Devin A, Rodriguez Y, Liu ZG . Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis. Genes Dev 1999; 13:2514–2526.
Yu L, Alva A, Su H, et al. Regulation of an ATG7-beclin 1 program of autophagic cell death by caspase-8. Science 2004; 304:1500–1502.
Shen HM, Lin Y, Choksi S, et al. Essential roles of receptor-interacting protein and TRAF2 in oxidative stress-induced cell death. Mol Cell Biol 2004; 24:5914–5922.
Festjens N, Vanden Berghe T, Cornelis S, Vandenabeele P . RIP1, a kinase on the crossroads of a cell's decision to live or die. Cell Death Differ 2007; 14:400–410.
Lin Y, Choksi S, Shen HM, et al. Tumor necrosis factor-induced nonapoptotic cell death requires receptor-interacting protein-mediated cellular reactive oxygen species accumulation. J Biol Chem 2004; 279:10822–10828.
Devadas S, Hinshaw JA, Zaritskaya L, Williams MS . Fas-stimulated generation of reactive oxygen species or exogenous oxidative stress sensitize cells to Fas-mediated apoptosis. Free Radic Biol Med 2003; 35:648–661.
Zhang AY, Yi F, Zhang G, Gulbins E, Li PL . Lipid raft clustering and redox signaling platform formation in coronary arterial endothelial cells. Hypertension 2006; 47:74–80.
Meurer R, MacIntyre DE . Lack of effect of pertussis toxin on TNF-alpha-induced formation of reactive oxygen intermediates by human neutrophils. Biochem Biophys Res Commun 1989; 159:763–769.
Lee MW, Park SC, Kim JH, et al. The involvement of oxidative stress in tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)-induced apoptosis in HeLa cells. Cancer Lett 2002; 182:75–82.
Shakibaei M, Schulze-Tanzil G, Takada Y, Aggarwal BB . Redox regulation of apoptosis by members of the TNF superfamily. Antioxid Redox Signal 2005; 7:482–496.
Reinehr R, Becker S, Eberle A, Grether-Beck S, Haussinger D . Involvement of NADPH oxidase isoforms and Src family kinases in CD95-dependent hepatocyte apoptosis. J Biol Chem 2005; 280:27179–27194.
Suzuki Y, Ono Y, Hirabayashi Y . Rapid and specific reactive oxygen species generation via NADPH oxidase activation during Fas-mediated apoptosis. FEBS Lett 1998; 425:209–212.
Kim YS, Morgan MJ, Choksi S, Liu ZG . TNF-induced activation of the Nox1 NADPH oxidase and its role in the induction of necrotic cell death. Mol Cell 2007; 26:675–687.
Banfi B, Clark RA, Steger K, Krause KH . Two novel proteins activate superoxide generation by the NADPH oxidase NOX1. J Biol Chem 2003; 278:3510–3513.
Geiszt M, Lekstrom K, Witta J, Leto TL . Proteins homologous to p47phox and p67phox support superoxide production by NAD(P)H oxidase 1 in colon epithelial cells. J Biol Chem 2003; 278:20006–20012.
Takeya R, Ueno N, Kami K, et al. Novel human homologues of p47phox and p67phox participate in activation of superoxide-producing NADPH oxidases. J Biol Chem 2003; 278:25234–25246.
Cheng G, Lambeth JD . NOXO1, regulation of lipid binding, localization, and activation of Nox1 by the Phox homology (PX) domain. J Biol Chem 2004; 279:4737–4742.
Newburger PE, Dai Q, Whitney C . In vitro regulation of human phagocyte cytochrome b heavy and light chain gene expression by bacterial lipopolysaccharide and recombinant human cytokines. J Biol Chem 1991; 266:16171–16177.
De Keulenaer GW, Alexander RW, Ushio-Fukai M, Ishizaka N, Griendling KK . Tumour necrosis factor alpha activates a p22phox-based NADH oxidase in vascular smooth muscle. Biochem J 1998; 329 (Pt 3):653–657.
Moe KT, Aulia S, Jiang F, et al. Differential upregulation of Nox homologues of NADPH oxidase by tumor necrosis factor-alpha in human aortic smooth muscle and embryonic kidney cells. J Cell Mol Med 2006; 10:231–239.
Frey RS, Rahman A, Kefer JC, Minshall RD, Malik AB . PKCzeta regulates TNF-alpha-induced activation of NADPH oxidase in endothelial cells. Circ Res 2002; 90:1012–1019.
Utsumi T, Klostergaard J, Akimaru K, et al. Modulation of TNF-alpha-priming and stimulation-dependent superoxide generation in human neutrophils by protein kinase inhibitors. Arch Biochem Biophys 1992; 294:271–278.
Akimaru K, Utsumi T, Sato EF, et al. Role of tyrosyl phosphorylation in neutrophil priming by tumor necrosis factor-alpha and granulocyte colony stimulating factor. Arch Biochem Biophys 1992; 298:703–709.
Dewas C, Dang PM, Gougerot-Pocidalo MA, El-Benna J . TNF-alpha induces phosphorylation of p47(phox) in human neutrophils: partial phosphorylation of p47phox is a common event of priming of human neutrophils by TNF-alpha and granulocyte-macrophage colony-stimulating factor. J Immunol 2003; 171:4392–4398.
Dang PM, Stensballe A, Boussetta T, et al. A specific p47phox -serine phosphorylated by convergent MAPKs mediates neutrophil NADPH oxidase priming at inflammatory sites. J Clin Invest 2006; 116:2033–2043.
Chenevier-Gobeaux C, Simonneau C, Therond P, et al. Implication of cytosolic phospholipase A2 (cPLA2) in the regulation of human synoviocyte NADPH oxidase (Nox2) activity. Life Sci 2007; 81:1050–1058.
Goossens V, Grooten J, De Vos K, Fiers W . Direct evidence for tumor necrosis factor-induced mitochondrial reactive oxygen intermediates and their involvement in cytotoxicity. Proc Natl Acad Sci USA 1995; 92:8115–8119.
Sakon S, Xue X, Takekawa M, et al. NF-kappaB inhibits TNF-induced accumulation of ROS that mediate prolonged MAPK activation and necrotic cell death. EMBO J 2003; 22:3898–3909.
Ventura JJ, Cogswell P, Flavell RA . Baldwin AS Jr, Davis RJ. JNK potentiates TNF-stimulated necrosis by increasing the production of cytotoxic reactive oxygen species. Genes Dev 2004; 18:2905–2915.
Festjens N, Kalai M, Smet J, et al. Butylated hydroxyanisole is more than a reactive oxygen species scavenger. Cell Death Differ 2006; 13:166–169.
Sidoti-de Fraisse C, Rincheval V, Risler Y, Mignotte B, Vayssiere JL . TNF-alpha activates at least two apoptotic signaling cascades. Oncogene 1998; 17:1639–1651.
Goossens V, De Vos K, Vercammen D, et al. Redox regulation of TNF signaling. BioFactors (Oxford, England) 1999; 10:145–156.
Ott M, Gogvadze V, Orrenius S, Zhivotovsky B . Mitochondria, oxidative stress and cell death. Apoptosis 2007; 12:913–922.
Lee SB, Bae IH, Bae YS, Um HD . Link between mitochondria and NADPH oxidase 1 isozyme for the sustained production of reactive oxygen species and cell death. J Biol Chem 2006; 281:36228–36235.
Nakashima I, Kato M, Akhand AA, et al. Redox-linked signal transduction pathways for protein tyrosine kinase activation. Antioxid Redox Signal 2002; 4:517–531.
Groen A, Lemeer S, van der Wijk T, et al. Differential oxidation of protein-tyrosine phosphatases. J Biol Chem 2005; 280:10298–10304.
Kamata H, Honda S, Maeda S, et al. Reactive oxygen species promote TNFalpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 2005; 120:649–661.
den Hertog J, Groen A, van der Wijk T . Redox regulation of protein-tyrosine phosphatases. Arch Biochem Biophys 2005; 434:11–15.
Wang T, Arifoglu P, Ronai Z, Tew KD . Glutathione S-transferase P1-1 (GSTP1-1) inhibits c-Jun N-terminal kinase (JNK1) signaling through interaction with the C terminus. J Biol Chem 2001; 276:20999–21003.
Adler V, Yin Z, Fuchs SY, et al. Regulation of JNK signaling by GSTp. EMBO J 1999; 18:1321–1334.
Tobiume K, Matsuzawa A, Takahashi T, et al. ASK1 is required for sustained activations of JNK/p38 MAP kinases and apoptosis. EMBO Rep 2001; 2:222–228.
Saitoh M, Nishitoh H, Fujii M, et al. Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1. EMBO J 1998; 17:2596–2606.
Liu H, Nishitoh H, Ichijo H, Kyriakis JM . Activation of apoptosis signal-regulating kinase 1 (ASK1) by tumor necrosis factor receptor-associated factor 2 requires prior dissociation of the ASK1 inhibitor thioredoxin. Mol Cell Biol 2000; 20:2198–2208.
Noguchi T, Takeda K, Matsuzawa A, et al. Recruitment of tumor necrosis factor receptor-associated factor family proteins to apoptosis signal-regulating kinase 1 signalosome is essential for oxidative stress-induced cell death. J Biol Chem 2005; 280:37033–37040.
Liu J, Lin A . Role of JNK activation in apoptosis: a double-edged sword. Cell Res 2005; 15:36–42.
Shen HM, Liu ZG . JNK signaling pathway is a key modulator in cell death mediated by reactive oxygen and nitrogen species. Free Radic Biol Med 2006; 40:928–939.
Karin M, Lin A . NF-kappaB at the crossroads of life and death. Nat Immunol 2002; 3:221–227.
Tang G, Minemoto Y, Dibling B, et al. Inhibition of JNK activation through NF-kappaB target genes. Nature 2001; 414:313–317.
Papa S, Zazzeroni F, Bubici C, et al. Gadd45 beta mediates the NF-kappa B suppression of JNK signalling by targeting MKK7/JNKK2. Nat Cell Biol 2004; 6:146–153.
Pham CG, Bubici C, Zazzeroni F, et al. Ferritin heavy chain upregulation by NF-kappaB inhibits TNFalpha-induced apoptosis by suppressing reactive oxygen species. Cell 2004; 119:529–542.
Jones PL, Ping D, Boss JM . Tumor necrosis factor alpha and interleukin-1beta regulate the murine manganese superoxide dismutase gene through a complex intronic enhancer involving C/EBP-beta and NF-kappaB. Mol Cell Biol 1997; 17:6970–6981.
Tang F, Tang G, Xiang J, et al. The absence of NF-kappaB-mediated inhibition of c-Jun N-terminal kinase activation contributes to tumor necrosis factor alpha-induced apoptosis. Mol Cell Biol 2002; 22:8571–8579.
Reuther-Madrid JY, Kashatus D, Chen S, et al. The p65/RelA subunit of NF-kappaB suppresses the sustained, antiapoptotic activity of Jun kinase induced by tumor necrosis factor. Mol Cell Biol 2002; 22:8175–8183.
Papa S, Zazzeroni F, Pham CG, Bubici C, Franzoso G . Linking JNK signaling to NF-kappaB: a key to survival. J Cell Sci 2004; 117:5197–5208.
Nakano H, Nakajima A, Sakon-Komazawa S, et al. Reactive oxygen species mediate crosstalk between NF-kappaB and JNK. Cell Death Differ 2006; 13:730–737.
Li M, Beg AA . Induction of necrotic-like cell death by tumor necrosis factor alpha and caspase inhibitors: novel mechanism for killing virus-infected cells. J Virol 2000; 74:7470–7477.
Chan FK, Shisler J, Bixby JG, et al. A role for tumor necrosis factor receptor-2 and receptor-interacting protein in programmed necrosis and antiviral responses. J Biol Chem 2003; 278:51613–51621.
Acknowledgements
The authors' research is supported by the Intramural Research Program of the National Institutes of Health, National Cancer Institute, Center for Cancer Research, USA.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is cited by
-
Targeting necroptosis in fibrosis
Molecular Biology Reports (2023)
-
Intravital electrochemical nanosensor as a tool for the measurement of reactive oxygen/nitrogen species in liver diseases
Journal of Nanobiotechnology (2022)
-
OTUD1 deubiquitinase regulates NF-κB- and KEAP1-mediated inflammatory responses and reactive oxygen species-associated cell death pathways
Cell Death & Disease (2022)
-
Expression of TXNIP is associated with angiogenesis and postoperative relapse of conventional renal cell carcinoma
Scientific Reports (2021)
-
GKN2 promotes oxidative stress-induced gastric cancer cell apoptosis via the Hsc70 pathway
Journal of Experimental & Clinical Cancer Research (2019)
