


















« Prev Next »
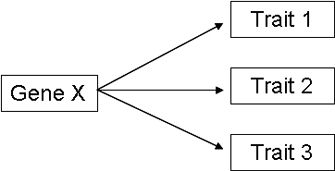
Figure 1: Diagram of pleiotropy.
A pleiotropic gene is a single gene that controls more than one trait.
© 2008 Nature Education All rights reserved. 
Today, we know that Mendel's observations were the result of pleiotropy, or the phenomenon in which a single gene contributes to multiple phenotypic traits. In this case, the seed coat color gene, denoted a, was not only responsible for seed coat color, but also for flower and axil pigmentation (Fairbanks & Rytting, 2001).
The term pleiotropy is derived from the Greek words pleio, which means "many," and tropic, which means "affecting." Genes that affect multiple, apparently unrelated, phenotypes are thus called pleiotropic genes (Figure 1). Pleiotropy should not be confused with polygenic traits, in which multiple genes converge to result in a single phenotype.
Examples of Pleiotropy
In some instances of pleiotropy, the influence of the single gene may be direct. For example, if a mouse is born blind due to any number of single-gene traits (Chang et al., 2002), it is not surprising that this mouse would also do poorly in visual learning tasks. In other instances, however, a single gene might be involved in multiple pathways. For instance, consider the amino acid tyrosine. This substance is needed for general protein synthesis, and it is also a precursor for several neurotransmitters (e.g., dopamine, norepinephrine), the hormone thyroxine, and the pigment melanin. Thus, mutations in any one of the genes that affect tyrosine synthesis or metabolism may affect multiple body systems. These and other instances in which a single gene affects multiple systems and therefore has widespread phenotypic effects are referred to as indirect or secondary pleiotropy (Grüneberg, 1938; Hodgkin, 1998). Other examples of both direct and indirect pleiotropy are described in the sections that follow.
Fruit Flies and the Vestigial Gene
In the fruit fly Drosophila, the vestigial gene plays a critical role in wing development. In fact, if these flies are homozygous for the recessive form of the vestigial gene (vg), they will develop short wings, and they will be unable to fly as a direct result. Along with regulating wing development, the vg gene is also pleiotropic. Indirectly, the gene changes the number of egg strings in a fly's ovaries, alters the position of bristles on a fly's scutellum, and decreases the length of a fly's life (Caspari, 1952; Miglani, 2002).
Chickens and the Frizzle Trait
In 1936, researchers Walter Landauer and Elizabeth Upham observed that chickens that expressed the dominant frizzle gene produced feathers that curled outward rather than lying flat against their bodies (Figure 2). However, this was not the only phenotypic effect of this gene — along with producing defective feathers, the frizzle gene caused the fowl to have abnormal body temperatures, higher metabolic and blood flow rates, and greater digestive capacity. Furthermore, chickens who had this allele also laid fewer eggs than their wild-type counterparts, further highlighting the pleiotropic nature of the frizzle gene.

Figure 2: A chicken with the frizzle gene
© 2004 Richard Blatchford, Dept. of Animal Science UC Davis. All rights reserved.
Pigmentation and Deafness in Cats
Approximately 40% of cats with white fur and blue eyes are deaf (Hartl & Jones, 2005). An initial hint at the link between pigmentation and deafness was the observation that white cats with one blue eye and one yellow eye were deaf only on the blue-eyed side. Interestingly, this phenomenon isn't exclusive to cats; for example, Waardenburg syndrome is a disorder that acts in a similar manner in humans. Because the gene responsible for both of these phenotypes affects pigmentation as well as the ability to hear, the gene is pleiotropic. Research continues into the exact mechanisms by which this gene affects hearing. For instance, in mice, experiments involving pigment cells have shown that pigmentation plays a role in maintaining fluid in ear canals. Animals that lack the pigment also lack ear canal fluid, which causes their ear canals to collapse. In turn, this collapse contributes to degeneration of the auditory nerves, which results in deafness (Sunquist, 2007).
Antagonistic Pleiotropy
Not all instances of pleiotropy are so straightforward, however. For example, in humans, the p53 gene directs damaged cells to stop reproducing, thereby resulting in cell death. This gene helps avert cancer by preventing cells with DNA damage from dividing, but it also suppresses the division of stem cells, which allow the body to renew and replace deteriorating tissues during aging (Rodier et al., 2007). This situation is therefore an example of antagonistic pleiotropy, in which the expression of a single gene causes competing effects, some of which are beneficial and some of which are detrimental to the fitness of an organism. The idea of antagonistic pleiotropy is central to the theory of aging proposed by American biologist G. C. Williams in 1957. In particular, Williams suggested that while some genes, like p53, increase the odds of successful reproduction and fitness early in life, they actually decrease fitness later in life. Moreover, because the gene's harmful effects appear after reproduction is complete, the gene is not eliminated through natural selection.
Yet another example of antagonistic pleiotropy can be found in Drosophila. In their research, scientists Carla Sgrò and Linda Partridge (1999) observed that flies with high fecundity early in their lives also had decreased lifespans. Based on these observations, Sgrò and Partridge suggested that aging in Drosophila evolved in response to the damaging effects of reproduction earlier in life. It is still not known exactly which genes determine this fecundity-mortality connection. Nevertheless, this example highlights the idea that antagonistic pleiotropy can be a trade-off between beneficial and detrimental effects.
Pleiotropy in Humans
As touched upon earlier in this article, there are many examples of pleiotropic genes in humans, some of which are associated with disease. For instance, Marfan syndrome is a disorder in humans in which one gene is responsible for a constellation of symptoms, including thinness, joint hypermobility, limb elongation, lens dislocation, and increased susceptibility to heart disease. Similarly, mutations in the gene that codes for transcription factor TBX5 cause the cardiac and limb defects of Holt-Oram syndrome, while mutation of the gene that codes for DNA damage repair protein NBS1 leads to microcephaly, immunodeficiency, and cancer predisposition in Nijmegen breakage syndrome.
One of the most widely cited examples of pleiotropy in humans is phenylketonuria (PKU). This disorder is caused by a deficiency of the enzyme phenylalanine hydroxylase, which is necessary to convert the essential amino acid phenylalanine to tyrosine. A defect in the single gene that codes for this enzyme therefore results in the multiple phenotypes associated with PKU, including mental retardation, eczema, and pigment defects that make affected individuals lighter skinned (Paul, 2000).
The phenotypic effects that single genes may impose in multiple systems often give us insight into the biological function of specific genes. Pleiotropic genes can also provide us valuable information regarding the evolution of different genes and gene families, as genes are "co-opted" for new purposes beyond what is believed to be their original function (Hodgkin, 1998). Quite simply, pleiotropy reflects the fact that most proteins have multiple roles in distinct cell types; thus, any genetic change that alters gene expression or function can potentially have wide-ranging effects in a variety of tissues.
References and Recommended Reading
Caspari, E. Pleiotropic gene action. Evolution 6, 1–18 (1952)
Chang, B., et al. Retinal degeneration mutants in the mouse. Vision Research 42, 517–525 (2002)
Fairbanks, D. J., & Rytting, B. Mendelian controversies: A botanical and historical review. American Journal of Botany 88, 737–752 (2001)
Grüneberg, H. An analysis of the pleiotropic effects of a new lethal mutation in the rat (Mus norvegicus). Proceedings of the Royal Society of London: Series B, Biological Sciences 125, 123–144 (1938)
Hartl, D. L., & Jones, E. W. Genetics: Analysis of Genes and Genomes, 6th ed. (Boston, Jones and Bartlett, 2005)
Hodgkin, J. Seven types of pleiotropy. International Journal of Developmental Biology 42, 501–505 (1998)
Landauer, W., & Upham, E. Weight and size of organs in frizzle fowl. Storrs Agricultural Experiment Station Bulletin 210 (1936)
Miglani, G. S. Advanced Genetics ( Pangbourne, England, Alpha Science, 2002)
Paul, D. A double-edged sword. Nature 405, 515 (2000) doi:10.1038/35014676 (link to article)
Rodier, F., et al. Two faces of p53: Aging and tumor supression. Nucleic Acids Research 35, 7475–7484 (2007) doi:10.1093/nar/gkm744
Sgrò, C. M., & Partridge, L. A delayed wave of death from reproduction in Drosophila. Science 286, 2521–2524 (1999) doi:10.1126/science.286.5449.2521
Sunquist, F. Malaysian mystery leopards. National Wildlife 45 (2007) (Accessed September 19, 2008)
Williams, G. C. Pleiotropy, natural selection, and the evolution of senescence. Evolution 11, 398–411 (1957)










