


 Figure 1
Figure 1
















« Prev Next »
Although Gregor Mendel developed his groundbreaking principles of inheritance in the mid-1800s, the importance of his work went largely unnoticed by the scientific community until the early 1900s. At that time, numerous researchers began to conduct experiments that upheld many of Mendel's ideas; however, they also discovered several situations that represented apparent deviations from these principles.
For example, one of the earliest exceptions to normal Mendelian ratios was observed by geneticists William Bateson, Edith Rebecca Saunders, and Reginald C. Punnett in 1905. In their work with pea plants, these researchers noticed that not all of their crosses yielded results that reflected the principle of independent assortment--specifically, some phenotypes appeared far more frequently than traditional Mendelian genetics would predict. Based on these findings, the trio proposed that certain alleles must somehow be coupled with one another, although they weren't sure how this linkage occurred. The answer to this question came just seven years later, when Thomas Hunt Morgan used fruit flies to demonstrate that linked genes must be real physical objects that are located in close proximity on the same chromosome.
Bateson, Saunders, and Punnett Suspect Linkage
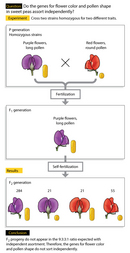
In 1905, William Bateson, Edith Rebecca Saunders, and Reginald Punnett were examining flower color and pollen shape in sweet pea plants (Bateson et al., 1905) by performing dihybrid crosses similar to those carried out by Gregor Mendel. In particular, these researchers crossed homozygous pea plants that had purple flowers and long pollen grains with homozygous plants that had red flowers and round pollen grains (Pierce, 2005). Prior to the cross, the trio noted that purple flowers (P) were dominant over red flowers (p), and that long pollen grains (L) were dominant over round pollen grains (l). The F1 generation of plants resulting from the PPLL x ppll cross was therefore doubly heterozygous (PpLl), and all of the F1 plants had purple flowers and long pollen grains.
Next, Bateson, Saunders, and Punnett decided to cross the F1 plants with each other. After this cross, the researchers expected the F2 generation to have a 9:3:3:1 ratio (nine plants with purple flowers and long pollen grains, to three plants with purple flowers and round pollen grains, to three plants with red flowers and long pollen grains, to one plant with red flowers and round pollen grains). Instead, they observed the results shown in Table 1 (Bateson et al., 1905), and these results were found to be statistically significant with a chi-square (c2) value of 969.
Table 1: Characteristics of the F2 Generation
| Phenotype | Expected | Observed | (Observed-Expected)2/Expected |
| Purple, long | 1199 | 1528 | 90.3 |
| Purple, round | 400 | 106 | 216.1 |
| Red, long | 400 | 117 | 200.2 |
| Red, round | 133 | 381 | 462.4 |
| Total | 2132 | 2132 | c2 = 969.0 |
As Table 1
indicates, Bateson, Saunders, and Punnett observed that their crosses produced
a deviation from the predicted Mendelian independent assortment ratios (Bateson
et al., 1905). The researchers
considered several explanations for this result, including epistasis, and they even tried to fit their data with
a 7:1:1:7 ratio. However, epistasis in F2 generations generally
results in a modified Mendelian ratio and is observed when genes affect the
same trait, such as flower color, so epistasis could not explain the deviation
seen here.
During their analysis, the researchers realized that there was an excess in the number of parental phenotypes (purple-long and red-round) in the F2 results. In particular, of the 2,132 F2 plants, 1,199 were expected to have purple flowers and long pollen grains, but instead, there were a whopping 1,528 plants with this phenotype. Similarly, only 133 plants were expected to have red flowers and round pollen grains, but the researchers observed nearly three times that many (381). It is now understood that the differences between the expected and observed results were statistically significant (P < 0.005), which means that the data could not be explained solely by chance. (Pearson's chi-square [c2] test can be used to statistically test for independent assortment of the phenotypes for flower color and pollen shape, as shown by the c2 value in Table 1.)
Because the parental phenotypes reappeared more frequently than expected, the three researchers hypothesized that there was a coupling, or connection, between the parental alleles for flower color and pollen grain shape (Bateson et al., 1905), and that this coupling resulted in the observed deviation from independent assortment. Indeed, Figure 1 shows an example of a cross between homozygous pea plants with purple flowers and long pollen grains and homozygous plants with red flowers and round pollen grains that exhibits linkage of the parental alleles.
But why are certain alleles linked? Bateson, Saunders, and Punnett weren't sure. In fact, it was not until the later work of geneticist Thomas Hunt Morgan that this coupling, or linkage, could be fully explained.
Morgan Finds Answers in the White-Eyed Fly
At the beginning of the twentieth century, Thomas Hunt Morgan's famous "fly room" at Columbia University was the site of many discoveries and "eureka" moments in the field of genetics. Morgan chose to use the prolific fruit fly Drosophila melanogaster as a model to study genetics. Then, for a period of three years, Morgan and his students struggled to find a way to create a fly that looked different from normal flies by treating these flies with heat, cold, X-rays, acids, bases, sugars, and other chemicals.
Finally, in 1910, Morgan fortuitously discovered a single fly with white eyes that did not result from any of his treatments. (Normal fruit flies have red eyes, not white eyes.) Morgan immediately crossed this white-eyed male fly to its red-eyed sisters (Morgan, 1910). Interestingly, when Morgan later inbred the heterozygous F1 red-eyed flies, the traits of the F2 progeny did not assort independently. Morgan expected a 1:1:1:1 ratio of red-eyed females, red-eyed males, white-eyed males, and white-eyed females. Instead, he observed the following phenotypes in his F2 generation:
- 2,459 red-eyed females
- 1,011 red-eyed males
- 782 white-eyed males
There were no white-eyed females, and Morgan wondered whether this was because the trait was sex-limited and only expressed in male flies. To test whether this was indeed the case, Morgan completed a second cross between the original white-eyed male fly and some of his F1 daughters. These crosses produced an F2 generation with the following phenotypes:
- 129 red-eyed females
- 132 red-eyed males
- 88 white-eyed females
- 86 white-eyed males
Thus, the results of this cross did produce white-eyed females, and the groups had approximately equal numbers. Morgan therefore hypothesized that the eye-color trait was connected with the sex factor (Morgan, 1910). This in turn led to the idea of genetic linkage, which means that when two genes are closely associated on the same chromosome, they do not assort independently (Morgan, 1911).
Morgan's proposal was an early suggestion that genes were real, physical objects that were located on chromosomes (Robbins, 2000). Indeed, knowledge of genetic linkage was critical to prove that genes were actual objects that could be inherited, undergo recombination, and be mapped to specific locations on chromosomes. For instance, after Morgan's findings were published, Reginald C. Punnett used this information to identify linkage groups in his previous plant studies, and he associated these linkage groups with chromosomes (Punnett, 1923; Punnett, 1927). Also, with this knowledge in place, Morgan and Alfred H. Sturtevant, his student, conducted further studies of linkage that provided information regarding gene location on chromosomes and ultimately resulted in gene mapping.
Why Didn’t Mendel Observe Linkage?
So if linkage
exists, why didn't Mendel detect it while carrying out his crosses in pea
plants? In part, this was the case because Mendel studied seven genes, and the
pea plant has seven chromosomes. Still, Mendel didn't choose pairs of genes
that were always on different chromosomes; some of the pairs of genes that Mendel
studied were actually on the same chromosomes, as shown in Figure 2 (Blixt,
1975).
Since the publication of Mendel's findings, other scientists have performed the pea plant crosses that could have shown linkage: i-a, v-fa, v-le, and fa-le. However, all of the pairs, except v-le, are so distantly located that Mendel would have been unable to detect linkage. In other words, although these pairs of genes are syntenic, they are not statistically linked. Therefore, they behave as though they independently assort. The v-le cross, on the other hand, would have shown linkage if Mendel had completed the cross. Possibly, with just one more cross, Mendel would have discovered linkage himself.
Summary
Thus, through their work with pea plants, Bateson, Saunders, and Punnett discovered an apparent exception to one of Mendel's foundational proposals: the principle of independent assortment. In particular, the trio suspected that certain alleles must somehow be linked to one another, thereby explaining why particular crosses yielded particular phenotypes in unusual numbers. A fuller explanation of this observation came only a few years later, when Thomas Hunt Morgan used fruit flies to show that linkage results when two genes are located near each other on the same chromosome. Since Morgan's time, this idea has served as the basis for continued research in the areas of gene mapping and recombination, to name but a few. Perhaps, if Mendel had carried out just one additional test cross with his plants back in the 1860s, he, rather than Bateson, Saunders, Punnett, and Morgan, would have been the one to uncover this fascinating area of study.
References and Recommended Reading
Bateson, W., et al. Experimental studies in the physiology of heredity. Reports to the Evolution Committee of the Royal Society 2, 1–55, 80–99 (1905)
Blixt, S. Why didn’t Gregor Mendel find linkage? Nature 256, 206 (1975) doi:10.1038/256206a0 (link to article)
Bridges, C. B. Salivary chromosome maps with a key to the banding of the chromosomes of Drosophila melanogaster. Journal of Heredity 26, 60–64 (1935)
———. A revised map of the salivary gland X-chromosome. Journal of Heredity 29, 11–13 (1938)
Hillers, K., & Villeneuve, A. Chromosome-wide control of meiotic crossing over in C. elegans. Current Biology 13, 1641–1647 (2003) doi:10.1016/j.cub.2003.08.026
Mendel, G. Experiments in plant hybridization. Trans. by William Bateson. (1866) (link to article)
Morgan, T. H. (1910). Sex-limited inheritance in Drosophila. Science 132, 120–122 (1910)
———. Random segregation versus coupling in Mendelian inheritance. Science 34, 384 (1911)
Passarge, E., et al. Incorrect use of the term synteny. Nature Genetics 23, 387 (1999) doi:10.1038/70486 (link to article)
Pierce, B. Genetics: A Conceptual Approach (New York, W. H. Freeman, 2005)
Punnett, R. C. Linkage in the sweet pea (Lathyrus odoratus). Journal of Genetics 13, 101–123 (1923)
———. Linkage groups and chromosome number in Lathyrus. Proceedings of the Royal Society of London: Series B, Containing Papers of a Biological Character 102, 236–238 (1927)
Robbins, R. J. Introduction to sex-limited inheritance in Drosophila, by T. H. Morgan (Electronic Scholarly Publishing, 2000) (link to article)
Sturtevant, A. H. The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association. Journal of Experimental Zoology, 14, 43–59 (1913)
Weiner, J. Time, Love, Memory: A Great Biologist and His Quest for the Origins of Behavior (New York, Random House, 1999)










