


















« Prev Next »

The melodic songs of thrushes, warblers, and finches are one of the signs that spring has arrived. With their elaborate songs, birds attract mates and defend their territories against conspecific (other individuals from the same species) rivals. In this article, I discuss why and how birds learn songs and what we can learn from their songs.
Song Diversity Between Species
The sounds that birds produce can be categorized into two distinct classes: calls and songs. A call is usually a short and simple vocalization that signals flight or danger and is produced throughout the year (Catchpole & Slater 2008). A song tends to be a long and complex vocalization produced during a breeding season. Songs are organized into several phrases (or motifs) which consist of series of syllables (Figure 1). Syllables, in turn, are made up of a collection of single notes (or elements). Each individual bird has its own song repertoire, which consists of different versions of a song, called a song type. There is large variation in repertoire size between species. In about a third of all songbird species, birds have only a single song type in their repertoire while in about 20% of all species, the repertoire consists of more than five songs (MacDougall-Shackleton 1997). In some cases, such as brown thrashers (Toxostoma rufum), song types can exceed 2,000!
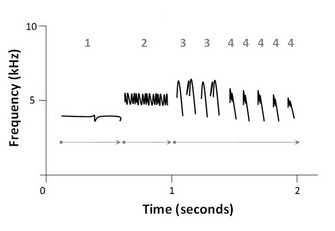
Figure 1: Schematic sound spectrogram of a white-crowned sparrow (Zonotrichia leucophrys) song
Arrows indicate phrase (or motif) and numbers indicate syllables which are made up of notes (or elements), the simplest unit of song.
© 2010 Nature Education All rights reserved. 
Species also vary in when they can learn new songs. Species such as white-crowned sparrows (Zonotrichia leucophrys) have a short period of a few months after hatching when they can learn songs (called close-ended learners) (Figure 2). In contrast, open-ended learners like European starlings (Sturnus vulgaris) and canaries (Serinus canaria) can learn and add new songs to their repertoire throughout their lives, usually on a seasonal basis. Which sex sings tends to depend on who competes to breed. In many species, only males sing; in others, both males and females sing equally in duets.
Why Birds Vary in Repertoire Size
As we already learned, variation in repertoire size among species is extensive, and the characteristic of having a repertoire with multiple song types likely evolved more than once in songbirds (MacDougall-Shackleton 1997). So what favors large and small repertoire size? According to the "repertoire hypothesis," a large repertoire is thought to be under directional sexual selection (Catchpole & Slater 2008, Beecher & Brenowitz 2005) where the choosy sex (often the females) favors one extreme end of a heritable, sexual trait. In fact, in some species females prefer males with large repertoires, and males with larger repertoires have a higher reproductive success (i.e., they produce more offspring) than those with a smaller repertoire (Catchpole & Slater 2008). On the other hand, the "song-sharing hypothesis" suggests that male-male competition selects for simpler, more homogenous songs (Beecher & Brenowitz 2005). When maintaining a breeding territory, familiar neighbors are preferred over newcomers because newcomers, who do not yet have a territory, are more likely to expand their territories than established neighbors. Territorial conflicts are energetically costly, and the song of a local population, or dialect, is thus thought to be favored as it can serve as an indication of regional origin, minimizing physically demanding defense measures.
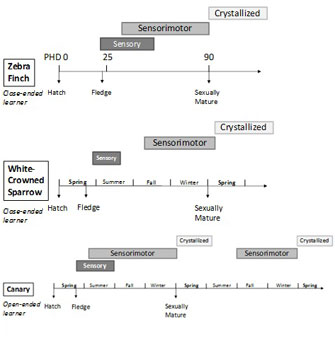
Figure 2: Timeline of song learning in three songbirds: zebra finches (Taeniopygia guttata), white-crowned sparrows (Zonotrichia leucophrys), and canaries (Serinus canaria)
Timing and duration of sensory and sensorimotor phases as well as song crystallization are indicated as a shaded box in relation to post-hatch days (PHD) or seasons.
© 2010 Nature Education All rights reserved.
Development of birdsong
William Thorpe pioneered scientific research on song learning in the late 1950s. He showed that chaffinches (Fringilla coelebs), raised in a laboratory as nestlings without exposure to adult males of the same species, develop abnormal songs (Thorpe 1958). However, when young of the same species were exposed to taped recordings of a wild chaffinch song (tutor songs), they sang species-specific songs as adults. This demonstrated that birds must learn songs early in life. This field of study was advanced further by Peter Marler and colleagues who showed that the song dialect is learned during a sensitive period, and birds have an innate predisposition to learn the songs of conspecifics (Marler 1970).
Auditory and Motor Phase
Similar to human speech development, song learning is a two-stage process (Figure 2). Birds first memorize a tutor song and form an auditory memory, or "template," in their brain (sensory phase). They then translate the inner template into motor activity by practicing, comparing their own vocalization to the template, and refining the songs (sensorimotor phase).
Sensory Phase
With some exceptions (e.g., gray catbirds [Dumetella carolinensis], sedge warblers [Acrocephalus schoenobaenus], brown-headed cowbirds [Molothrus ater]), birds raised in acoustic isolation during the sensory phase produce atypical songs as adults. This is because most young birds learn the species-specific songs during the first year of their lives. In the wild, birds grow up listening to the songs of a variety of different species. So why don't they learn the songs of multiple species? Studies have shown that even without prior experience in hearing their own species' songs, young birds still increase their heart rate and beg more when they hear conspecific songs, indicating that they are born with an innate ability to discriminate their species-specific songs (Brainard & Doupe 2002). If given a choice, young birds preferentially learn conspecific over heterospecific songs, and if birds are raised in acoustic isolation, they sing abnormal songs yet still with species-specific elements. Timing of the sensory phase varies among species, but this window aligns with when adults of the species sing the most (Zeigler & Marler 2008). Thus, together with the genetic predisposition for recognizing and learning species-specific songs, this minimizes the risk of learning wrong songs.
When the sensory phase ends varies among species, but this timing depends partly on experience. If young are raised only with heterospecific songs, birds can learn conspecific songs later than those raised hearing conspecific songs (Brainard & Doupe 2002). Interestingly, if birds are raised in acoustic isolation, the sensory phase can be extended even into adulthood in some species.
Sensorimotor Phase
At the beginning of the sensorimotor phase, young birds first produce generic, variable, and quiet vocalizations called subsong, which is similar to human baby babbling (Brenowitz et al. 1997). They then produce louder, more structured songs called plastic songs, which are still variable but contain some elements of the tutor song. Songs finally crystallize to stable stereotyped songs that are similar to the songs they memorized. During the sensorimotor phase, birds need to hear their own vocalization in order to develop normal songs. If juveniles are deafened after the sensory phase but before the sensorimotor phase, they develop aberrant songs (Konishi 1965).
Birds of certain species produce more sounds during the sensorimotor phase than the sounds they will produce in adulthood. This means that during song crystallization, those species select which sounds are incorporated into the crystallized song. As you might expect, this selection is not random. White-crowned sparrows, for instance, integrate songs that match the dialects of neighboring males into the crystallized song (Nelson & Marler 1994). This can be advantageous because males that sing local dialects have a higher reproductive success than those that sing foreign dialects (MacDougall-Shackleton et al. 2002). Cowbirds, on the other hand, attain songs that are more effective in triggering mating-like behaviors in females (West & King 1988). In either case, song selection during crystallization is based on functional significance to maximize reproductive output.
Hormonal Influences
Neuroplasticity describes the lifelong ability of the brain to form new neural connections depending on season and experience. The hormone testosterone is necessary for song crystallization by reducing plasticity and producing stereotyped songs (Bottjer & Johnson 1997). Testosterone administration during the sensory phase has little or no effect on song memorization. However, testosterone levels are high during song crystallization and in the spring, when songs are more stable. Testosterone administration prior to song crystallization triggers premature crystallization of simple songs, and both castration and blocking testosterone receptors can delay or prevent crystallization. Thus, the timing and degree of elevation in testosterone are essential in proper development of birdsongs.
Neural Control of Song Learning and Production
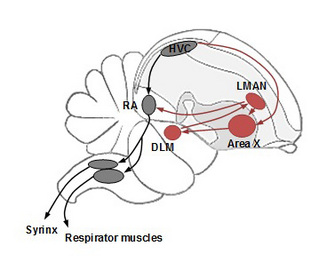
Figure 3: A sagittal view of a schematic bird brain depicting the neural circuit controlling song learning and production
The motor pathway necessary for song production is in black, and the anterior forebrain pathway necessary for song learning and plasticity is in red. Abbreviations for the brain nuclei: Area X (Area X of the medial striatum), DLM (medial portion of the dorsolateral nucleus of the thalamus), HVC (used as a proper name), LMAN (lateral subdivision of the magnocellular nucleus of the anterior nidopallium), and RA (robust nucleus of the arcopallium).
© 2010 Nature Education All rights reserved.
In the motor pathway, HVC projects onto RA (robust nucleus of the arcopallium). RA then sends axons to two separate motor nuclei whose motor neurons innervate the vocal organ in birds, called the syrinx, and the respiratory muscles to produce song while coordinating breathing. The AFP includes Area X (Area X of the medial striatum; homologue to mammalian basal ganglia), DLM (medial portion of the dorsolateral nucleus of the thalamus), and LMAN (lateral subdivision of the magnocellular nucleus of the anterior nidopallium). LMAN converges with the motor pathway at RA.
Studies deactivating parts of the neural circuit for birdsong via lesions highlight functions of the AFP. While lesions of LMAN, DLM, or Area X in adults have little effect on already-learned songs, lesions of these areas before song crystallization lead to abnormal songs (Brenowitz et al. 1997). Further studies have shown that lesions of Area X in juveniles prevent crystallization whereas lesions of LMAN in juveniles result in permanently crystallized, atypical songs. Lesions of LMAN, in fact, make their songs truly stable. It prevents adult birds from learning new songs and also shields already-learned song from deteriorating after deafening. Thus, the AFP facilitates song plasticity in both juveniles and adults, possibly via RA where the AFP and the motor pathway converge (Mooney 2009).
Implications for Research in Human Speech
There are striking similarities between the development of birdsong and human speech. In both cases, dialects and languages are culturally transmitted during a sensitive period of learning. For instance, similar to the sensory phase in song learning, distinguishing and producing sounds of the new language is difficult without an early exposure to the foreign language — before reaching early adolescence in humans (Brainard & Doupe 2002). Both birdsong and human speech are controlled by discrete neural circuitry and auditory feedback is essential in normal learning. These parallels have attracted scientists to use birdsong as a model for research in human speech pathology. In the past ten years, neuroscientists discovered that a mutation in one gene called FOXP2 causes a particular speech and language disorder (White et al. 2006). Using songbirds as a model, researchers found that the FOXP2 gene is necessary to accurately imitate the sounds of a tutor song (Haesler et al. 2007). These recent discoveries demonstrate that birdsong constitutes an excellent model for exploring the molecular basis and behavioral development of human speech.
Acknowledgments
I thank S. MacDougall-Shackleton, A. Diez, T. Farrell, Z. Hall, T. Luloff, S. Nebel, G. Salze, and K. Schmidt for their valuable input on the manuscript and G. Salze for his assistance in figure creation.
References and Recommended Reading
Beecher, M. D. & Brenowitz, E. A. Functional aspects of song learning in songbirds. Trends in Ecology and Evolution 20, 143–149 (2005).
Bottjer, S. W. & Johnson, F. Circuits, hormones, and learning: vocal behavior in songbirds. Journal of Neurobiology 33, 602–618 (1997).
Brainard, M. S. & Doupe, A. J. What songbirds teach us about learning. Nature 417, 351–358 (2002).
Brenowitz, E. A. et al. An introduction to birdsong and the avian song system. Journal of Neurobiology 33, 495–500 (1997).
Catchpole, C. K. & Slater, P. J. B. Bird song: Biological Themes and Variations, 2nd ed. Cambridge, UK: Cambridge University Press, 2008.
Haesler, S. et al. Incomplete and inaccurate vocal imitation after knockdown of FoxP2 in songbird basal ganglia nucleus area X. PLoS Biology 5, e321 2885–2897 (2007).
Konishi, M. The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Zeitschrift fur Tierpsychologie 22, 770–783 (1965).
MacDougall-Shackleton, S. A. "Sexual selection and the evolution of song repertoires." In Current Ornithology, vol. 14, eds. V. J. Nolan, E. D. Ketterson, & C. F. Thompson (New York, NY: Plenum Press, 1997): 81–124.
MacDougall-Shackleton, S. A. et al. Nonlocal male mountain white-crowned sparrows have lower paternity and higher parasite loads than males singing local dialect. Behavioral Ecology 13, 682–689 (2002).
Marler, P. A comparative approach to vocal learning: song development in white-crowned sparrows. Journal of Comparative & Physiological Psychology 71, 1–25 (1970).
Mooney, R. Neurobiology of song learning. Current Opinion in Neurobiology 19, 654–660 (2009).
Nelson, D. A. & Marler, P. Selection-based learning in bird song development. Proceedings of the National Academy of Sciences USA 91, 10498–10501 (1994).
Thorpe, W. H. The learning of song patterns by birds, with especial reference to the song of the chaffinch Fringilla coelebs. Ibis 100, 535–570 (1958).
West, M. J. & King, A. P. Female visual displays affect the development of male song in the cowbird. Nature 334, 244–246 (1988).
White, S. A. et al. Singing mice, songbirds, and more: models for FOXP2 function and dysfunction in human speech and language. Journal of Neuroscience 26, 10376–10379 (2006).
Zeigler, H. P. & Marler, P., eds. Neuroscience of Birdsong. Cambridge, UK: Cambridge University Press, 2008.










