



















« Prev Next »
How Sexual Selection Came To Be Recognized
Charles Darwin proposed that all living species were derived from common ancestors. The primary mechanism he proposed to explain this fact was natural selection: that is, that organisms better adapted to their environment would benefit from higher rates of survival than those less well equipped to do so. However he noted that there were many examples of elaborate, and apparently non-adaptive, sexual traits that would clearly not aid in the survival of their bearers. He suggested that such traits might evolve if they are sexually selected, that is if they increase the individual's reproductive success, even at the expense of their survival (Darwin 1871).
Darwin noted that sexual selection depends on the struggle between males to access females. He recognized two mechanisms of sexual selection: intrasexual selection, or competition between members of the same sex (usually males) for access to mates, and intersexual selection, where members of one sex (usually females) choose members of the opposite sex. The idea of cumbersome traits evolving to aid males in competition during aggressive encounters was readily accepted by scientists shortly after Darwin's publication. However, the idea of female mate choice was received with ridicule, and was not seriously reconsidered until nearly 80 years later (Cronin 1991). In the 40 years since, there has been much progress in our understanding of how sexual selection operates.
Which Sex is Under Stronger Selection?
Sex roles are defined by differences in gametes: females produce relatively few, highly nutritious (usually non-motile) gametes, whereas males produce comparatively abundant, smaller, motile gametes. Because only a single gamete of each type is required to produce an offspring, there will be an excess of male gametes that will not fertilize any eggs. This asymmetry leads to Bateman's principle, whereby female reproduction is primarily limited by their access to resources to nourish and produce these large gametes, whereas male reproduction is mainly limited by access to females (Bateman 1948). Therefore males typically compete among themselves for access to females, whereas females tend to be choosy and mate only with preferred males.
In sexually reproducing species, every offspring has one father and one mother, so the average reproductive success is equal for both males and females. A successful male can potentially sire many offspring. If a male gains a disproportionate share of reproduction, he will take away reproductive opportunities from other males, leading to a high reproductive variance among males. A successful female, on the other hand, will not take away reproductive opportunities from other females, leading to a smaller variance in reproductive success. The higher the reproductive variance, the stronger the effects of sexual selection (Figure 1). Strong sexual selection typically results in sexually dimorphic traits that are exaggerated, or more elaborate, in the sex with highest reproductive variance (Figure 1).
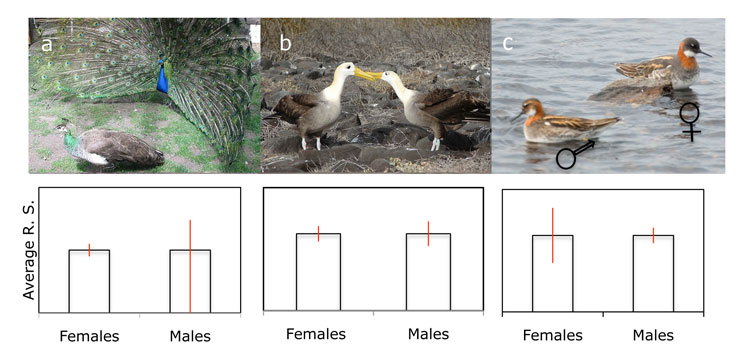
Figure 1: Variance in reproductive success explains which sex is subject to stronger sexual selection
Males and females in a population have the same average reproductive success (R. S., black bars) but they may differ in the reproductive variance among members of each sex (shown in red). Differences in the selection gradient will result in sexual dimorphism. (A) When males are subject to stronger sexual selection than females, males will evolve secondary sexual characters that result in marked differences between the sexes. Peacocks do not provide any parental care, and some males are more successful than others who may never reproduce, leading to marked dimorphism. (B) When males contribute to offspring care, the selection gradient is lower and the sexes will be monomorphic. Many seabirds are monogamous and raise offspring together and the sexes are indistinguishable. (C) When males provide all the parental care, the selection gradient can be reversed and females may have to compete for access to males, leading to reverse sexual dimorphism. Red-necked phalaropes compete for access to males who provide all the parental care. Females are larger and more aggressive than males. (Courtesty of Arthur Grosset)
The Role of Parental Care
Most species provide little or no care to their offspring, but in species where parental care is required, variance in reproductive success will be impacted not only by fertilization success, but also by the contribution of each sex to the care of the offspring (Trivers 1972). The degree and direction of sexual dimorphism can be explained by the relative selection gradients of each sex. If females provide more parental care than males, the variance in male reproductive success can be expected to be large, since females providing offspring care will not be immediately available for further reproduction and competition for available females will increase among males. The exaggerated tail of the (male) peacock compared to the shorter tail of the (female) peahen, indicates that males are under stronger sexual selection than females (Figure 1a). However, in species where biparental care is required to successfully raise offspring, variance in male reproductive success is generally lower, since males that are engaged in providing parental care will not be able to invest as much energy in pursuing additional mating opportunities. This situation often results in the emergence of sexually monomorphic species, in which the male and female look and behave in similar ways (Figure 1b). Finally, in rare situations where only males provide parental care, males can become the limiting resource for females. Under these circumstances, the variance in reproductive success may be high for females who then tend to monopolize access to one or more males to care for their offspring. Males may then become choosy about which females they mate with. This often results in reversed sexual dimorphism, such that females evolve more elaborate secondary sexual characters than males (Figure 1c).
How Does Sexual Selection Operate?
Sexual selection can operate both intra- and inter-sexually, either sequentially or simultaneously (Table 1). During intrasexual selection, members of the same sex attempt to outcompete rivals, often during direct encounters. Intrasexual selection is typically responsible for the evolution of male armaments such as deer antlers, beetle horns, and large body size, that provide individuals with an advantage when fighting off potential competitors. Individuals who are better able to exclude competitors, have a greater chance to acquire mates and father offspring. For example, dominant male red deer monopolize a group of females (also known as harem) by constantly fighting off competitors, and they father most of the offspring produced by the females. By contrast, intersexual selection results from interactions between the sexes, typically involving mate choice. The evolution of elaborate behavioral displays and morphological traits can often be explained as the result of intersexual selection. Usually, females tend to be more choosy, evaluating morphological and behavioral traits from potential mates to determine which will maximize their fitness. Males tend to compete with one another to gain the female's attention. An extreme example of intersexual selection can be found in species where males form leks where multiple males gather to display to females.
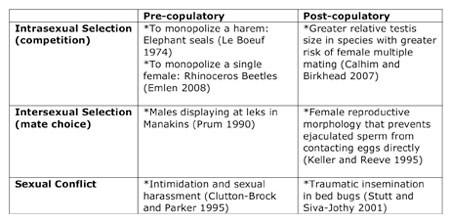
Table 1: Some examples of when and how sexual selection operates
Sexual selection episodes can occur before mating takes place (pre-copulatory), or during and after mating (post-copulatory), and they can occur within a sex (intrasexual) and between the sexes (intersexual).
Choosing a Mate
Why do females choose between males rather than mate at random, or with the first male they encounter? Females can directly increase their reproductive success by mating with certain, select males and acquiring direct benefits. For example, females can gain increased access to food, protection from harassing males, or help in raising offspring, and avoid being infected with parasites or other diseases by choosing healthy males. However there are instances where females do not appear to gain any direct benefit from males, yet they still discriminate among them. Under these conditions, females likely gain indirect benefits via their offspring. These indirect benefits are usually genetic rather than resource based. By choosing certain males, their offspring will likely inherit genes that tend to increase their fitness. Males often evolve traits and displays that advertise their ability to provide direct and indirect benefits, and females evolve preferences for these traits. Two major mechanisms to account for female mate choice have been proposed: good genes, and Fisherian arbitrary processes.
Good Genes
Under the ‘good genes’ scenario, differences among males provide females with information about the genetic qualities of the different males that can be inherited by the offspring. Under the ‘good genes,’ just as in the ‘direct benefits’ models, there is correspondence between the putative roles of natural versus sexual selection, since preferring certain males can result in a female gaining higher viability, fecundity, and reproductive success, for her offspring. Good genes can be those that allow males to carry a ‘handicap,’ yet survive despite having a cumbersome trait (Zahavi 1975), genes that signal resistance to disease (Hamilton & Zuk 1982), or genes that are more compatible with those of the female (Trivers 1972). Evidence of female choice for good genes remains scarce despite decades of studies of female mate choice in many taxa. This apparent lack of success continues to create debate as to the importance of the good genes model in the field.
Fisherian Arbitrary Choice
Named after R. A. Fisher, who originally proposed it, this model suggests that female preference can evolve for arbitrary traits that do not provide information about the male's quality, and that therefore do no reinforce the effects of natural selection. If females evolve a preference for a particular trait, males bearing that trait will be selected as mates. This assortative mating will establish a genetic correlation between the preference and the trait. The fitness advantage of the arbitrary trait exists only as a result of its covariance with the preference. By choosing a male with a particular trait value, the females gain the indirect benefit of producing offspring that will be more sexually attractive to females that carry the preference (Fisher 1930). This type of process can result in a runaway positive feedback loop, whereby the trait becomes more exaggerated as selection on the preference increases (Fisher 1930), but other models have shown that such a feedback loop is only one of many possible evolutionary outcomes of the Fisherian process (Lande 1981; Kirkpatrick 1982).
When Does Sexual Selection Act?
Sexual selection can affect reproductive success at multiple reproductive stages. First, it acts during all the processes that lead to acquiring mating opportunities (i.e., excluding competitors, attracting, selecting and/or retaining mates). Darwin referred exclusively to pre-copulatory sexual selection in his discussions, erroneously assuming that mating would inevitably result in reproductive success. In recent years, evidence that copulatory and post-copulatory events play an important role in determining the outcome of fertilization and reproduction has been increasing. Post-copulatory selection refers to the events that occur during and after mating. Post-copulatory male-male competition is known as sperm competition a term coined by Parker (1970) who recognized that when females mate with multiple males, their ejaculates compete inside the female reproductive tract for access to eggs. Sperm competition has resulted in the evolution of morphologically modified sperm that increase the likelihood of fertilization in many taxa (Birkhead & Moller 1998). Post-copulatory female choice refers to the ability of females to affect the likelihood that sperm from a particular male fertilizes their eggs, and their decision to invest in offspring based on the identity of the male with whom they mate. Females exert this choice via morphological, chemical and behavioral adaptations. This type of selection is called cryptic choice because it occurs inside the female reproductive tract and cannot be detected from behavioral studies alone (Eberhard 1996).
Conflict Between the Sexes
Although both sexes are seeking to optimize their reproductive success, their genetic interests are not aligned, resulting in sexual conflict (Parker 1979). Traits that allow a male to increase his reproductive success at the expense of the female will be positively selected if the female mates with multiple males. These traits will be genetically transmitted and spread in the population, despite their negative effects on female reproductive success, if the reproductive success of these males is higher than that of males lacking such traits (Parker 1979). Sexual conflict can often result in an evolutionary arms race, whereby the evolution of a trait that imposes harm on one sex will result in evolution of a counter-trait to mitigate the harm on the affected sex, with subsequent escalation in both (Chapman et al. 2003). Examples of sexual conflict include traumatic insemination in bed bugs, copulatory grasping and anti-grasping structures in waterstriders, and genital coevolution in waterfowl.
References and Recommended Reading
Bateman, A. J. Inter-sexual selection in Drosophila. Heredity 2, 349-368 (1948).
Birkhead, T. R. & Moller, A. P. Sperm Competition and Sexual Selection. San Diego, CA: Academic Press, 1998.
Calhim, S. & Birkhead, T. R. Testes size in birds: quality versus quantity — assumptions, errors and estimates. Behavioral Ecology 18, 271-275 (2007).
Chapman, T., Arnqvist, G. et al. Sexual conflict. Trends in Ecology and Evolution 3, 41-47 (2003).
Clutton-Brock, T. H. & Parker, G. A. Sexual coercion in animal societies. Animal Behavior 49, 1345-1365 (1995).
Cronin, H. The Ant and the Peacock. Cambridge, UK: Cambridge University Press, 1991.
Darwin, C. The Descent of Man, and Selection in Relation to Sex. London, UK: Murray, 1871.
Eberhard, W. Female Control: Sexual Selection by Cryptic Female Choice. Princeton, NJ: Princeton University Press, 1996.
Emlen, D. J. The Evolution of Animal Weapons. Annual Review of Ecology, Systematics, and Evolution 39, 387-413 (2008).
Fisher, R. A. The Genetical Theory of Natural Selection. Oxford, UK: Clarendon Press, 1930.
Hamilton, W. D. & Zuk, M. Heritable true fitness and bright birds: a role for parasites? Science 218, 384-387 (1982).
Keller, L. & Reeve, H. K. Why do females mate with multiple males? The sexually selected sperm hypothesis. Advanced Studies in Behavior, 24, 291-315 (1997).
Kirkpatrick, M. Sexual selection and the evolution of female choice. Evolution 82, 1-12 (1982).
Lande, R. Models of speciation by sexual selection on polygenic traits. Proceedings of the National Academy of Sciences, USA 78, 3721-3725 (1981).
LeBoeuf, B. Male-male competition and reproductive success in Elephant seals. American Zoologist 14, 163- 176 (1974)
Parker, G. Sperm competition and its evolutionary consequences in the insects. Biological Reviews 45, 525-567 (1970).
Parker, G. Sexual selection and sexual conflict. In Sexual Selection and Reproductive Competition in Insects. eds. Blum, M. S. & Blum, N. A. (New York: Academic Press, 1979): 123-166.
Prum, R. O. Phylogenetic analysis of the evolution of display behavior in the neotropical manakins (Aves: Pipridae). Ethology 84, 202-231 (1990).
Stutt, A. D., Siva-Jothy, M. T. Traumatic insemination and sexual conflict in the bed bug Cimex lectularius. Proceedings of the National Academy of Sciences, U.S.A., 98, 5683-5687 (2001)
Trivers, R. L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man 1871-1971. ed. Campbell, B. (London: Heinemann 1972): 136-179.
Zahavi, A. Mate selection: a selection for a handicap. Journal of Theoretical Biology 53, 205-214 (1975).










