


















« Prev Next »

A mating system describes how males and females pair when choosing a mate. Males and females differ greatly in the investment each makes to reproduce, and may therefore approach mating with differing strategies. To study these differences, scientists observe mating systems and describe how males and females come together. When choosing mates, animals evolve species-typical strategies for maximizing their reproductive success — this results in considerable diversity among animal species in their mating patterns.
In this article we first discuss why sexual reproduction exists, and how differences between males and females affect mating systems. We move on to consider the evolution of mate choice, and then we describe the types of mating systems found in animals.
The Evolution of Sex
Asexually reproducing animals pass on all of their chromosomes, and consequently all copies of each gene, to their offspring. In contrast, due to meiosis, diploid sexually reproducing animals have two copies of each chromosome but only pass one copy of each chromosome on to an egg or sperm cell. This means that a sexually reproducing diploid animal only passes half of its total genes on to its offspring. Despite the cost of losing half of the potential passage of genes to the next generation, sexual reproduction is much more common than asexual reproduction among animals because it provides several evolutionary advantages. The major advantage of sexual reproduction comes from genetic recombination. Genetic recombination allows an organism's offspring to be genetically diverse. Sexual reproduction increases the chances of acquiring favorable mutations and is unlikely to propagate deleterious ones. Genetic diversity within a group of offspring is advantageous as the local environment changes. This idea becomes clear when we examine organisms that can reproduce both sexually and asexually. Aphids, for example, will favor asexual reproduction when their environment is stable. When the environment is going to turn cold, most species of aphids reproduce sexually, because sexual reproduction produces eggs that are freeze tolerant and can diapause during the winter (Simon et al. 2002). Genetic diversity may also lead to evolved defenses against parasites and disease. The mud snail, Potamopyrgus antipodarum, is host to several trematode parasites. Sexual individuals of this species are more common in areas where risk of trematode infection is high. In areas where the risk of infection is low, asexual individuals have displaced sexual ones (King et al. 2009). This suggests that the genetic diversity acquired from sexual reproduction is necessary for this species to defend against parasites, as asexual individuals may not easily survive in areas where parasites are high.
Sexual reproduction often involves evolutionary differentiation of males and females. Females typically produce significantly fewer gametes (eggs) than males and invest heavily in each one. On the other hand, males produce many gametes (sperm) and invest little into each one. These strong differences in gamete investment between the sexes leads to reproductive strategies between the sexes that, in some cases, conflict. Females may spend more care than males selecting a mate due to the high cost of their gametes.

Figure 1: A male bighorn sheep.
The large horns are used in combat between males during mating season, and likely evolved as a result of intrasexual selection.
© 2012 Nature Education Photo courtesy of Michael D. Breed. All rights reserved. 
Variance in Mating Success and Bateman's Principle
A key element of the study of mating systems is understanding how many mates an animal has in its lifetime. Bateman's principle helps to make predictions about mating success and number of mates. Bateman's principle postulates that variance among females in mating success is low, whereas variance among males in mating success is high. This stems from the fact that one mating in females should be enough to fertilize all their eggs whereas in males reproductive success is based on the number of times they have mated. In other words, nearly all females in a population mate and have offspring, but relatively few males mate successfully (Figure 2). Those males that do mate tend to mate with many females-thus a few males have very high reproductive output, but many males have little or no reproductive output (Bateman 1948). This leads to the prediction that sexual selection should act more strongly on males, leading to greater elaboration of behavior and structures used in attracting mates in males than in females.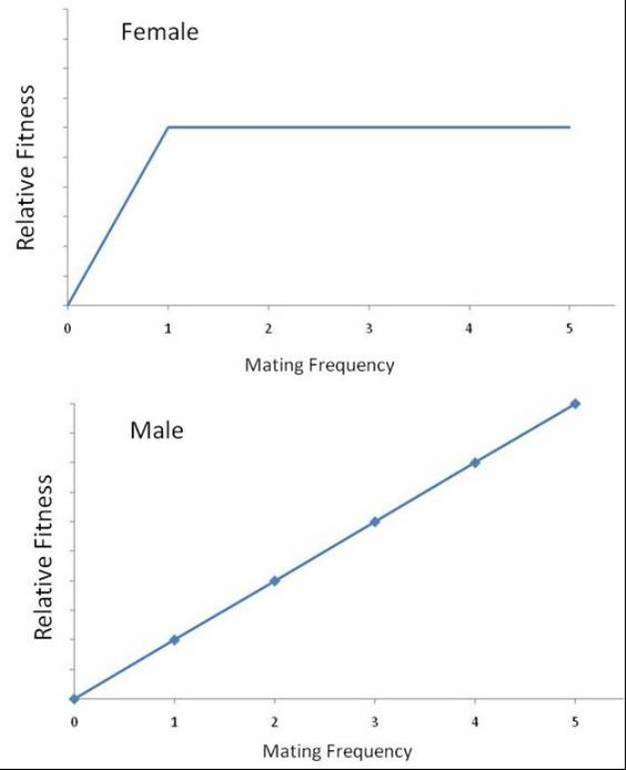
Figure 2: Bateman’s principle.
These figures illustrate Bateman’s principle — after one mating, female mating frequency increases and relative fitness remains constant, as the sperm from one mating is adequate to fertilize all the female’s eggs. In males, as mating frequency increases relative fitness also increases proportionally.
© 2012 Nature Education Modified from Hauber & Lacey 2005. All rights reserved.
Criticisms of Bateman's theory focus on the generality of the predictions. Contrary to the predictions of Bateman's principle, there are several possible advantages to female multiple matings. The female cichlid fish Pseudotropheus spiliopterus mates with any male they meet because they have a high risk of getting predated and a small population. This often leads to multiple matings by a single female (Kellogg et al.1998). Mating with any male that is seen ensures that these cichlids have a chance at producing offspring. The female Malawi blue cichlid has a high population but still participates in multiple matings. In this case multiple matings occur to avoid inbreeding and increase genetic diversity among the offspring (Kellogg et al. 1998). Additionally, multiple matings by females may increase the likelihood that they will find a compatible mate, one that is not sterile, or even help prevent infanticide.
Female Mate Choice
Mate choice is also a key element of mating systems. In most species, females are choosier when picking a mate than males. A significant reason for this is the higher investment females make in each gamete than males. Females may prefer certain males for a variety of reasons, including "good genes", meaning that the male has attributes which predict better survivorship of the offspring, good potential parenting by the male, or possession of resources by the male that will support the offspring during their growth and development.
Additionally, in most species, females are more likely to provide parental care. Females that carefully select their mates are at a lower risk of losing their reproductive investment. Males may be under strong selection for certain traits that are favored by females. Most females look at these traits as indicators of their partner's fitness. Selection favors females that choose males that enhance the likelihood of her offspring's success. Males with more elaborate ornamentation, or that are more colorful, can be displaying a good indicator of value as a mate, and may win the chance to mate with a particular female. (Figure 3). Although mating is important, it can be a costly event — females are predicted to be choosier about selecting their mates than males because of risks during mating, such as aggression or disease transmission, which may negatively impact the female's reproductive output.

Figure 3: Eyespots from peacock tail feathers.
Elaborate ornamentation usually evolves in intrasexual selection and is used in mate choice.
© 2012 Nature Education Photo courtesy of Michael D. Breed. All rights reserved.
Male Mate Choice
The importance of male mate choice is controversial. Older theory predicts that male mate choice should be less common in animals. However it plays an important role in many mating systems, and the cost of mating for males may have been underestimated in earlier studies. Male mate choice occurs most often when males are substantially involved in caring for their offspring, or when there is great variation in the quality of the females as mates within a population. If males are choosy about their mate, then over time females may evolve ornamentation or coloration that is subject to sexual selection.Types of Mating Systems
Monogamy
Social monogamy is the behavioral pairing of a single male with a single female. It is most common in birds and rare in other animals (Figure 4). Theoretically, individuals in monogamous pairs will both contribute to the defense and parental care of offspring. Choosing an inappropriate mate could have a high fitness cost (see the sections above for more on mate choice). Because the costs of poor mate choice in monogamous species can be so high, in some instances organisms engage in strategies of either serial monogamy or extra-pair copulations. Extra-pair copulations are very common in birds (Petrie et al. 1998, Stutchberry 1998). Monogamy reduces the potential for genetic variation among a female's offspring. By mating with more than one male over the course of her lifetime, a female gains higher genetic variation among her offspring. The benefits of monogamy, which are shared parental care and territorial resources, are maintained by having only one mate at a time, or by concealing extra-pair partnerships.
Figure 4: Blue-footed boobies.
Many bird species, such as these blue-footed boobies are monogamous.
© 2012 Nature Education Photo courtesy of Michelle O. Krasnec. All rights reserved.
Polygyny
Polygyny is the association of one male with multiple females. This mating system is found in a few birds and insects, but is most common in mammals. Polygyny is a strategy used by males to increase their reproductive fitness.
Resource Defense Polygyny
In resource defense polygyny, groups of females are attracted to a resource — males then compete for territorial possession of the resource, and, by extension, mating priority with females at the resource (Beletsky 1994). Thus, individual males form territories centered on resources needed for successful mating (McCracken 1981).
Harems
Another common type of polygyny is membership in a harem, a defended group of females associated with one male. Females may initially associate in a harem for group defense, or they may be herded together by a male. Males compete for control of the groups. Harems typically exhibit a dominance hierarchy among the females in the group.
Leks
A lek is an aggregation of males that are each seeking to attract a mate. Within a lek, males typically perform sexual displays. Unlike most other mating systems, leks are not associated with resources. Aggregations of males may be near particularly attractive females or in areas where females are likely to travel (Lank et al. 1995, Aspbury & Gibson 2004). It is thought that males form leks because they attract more females than do isolated males. Attracting more females is a strategy used by males to help increase their reproductive success.
Polyandry
Polyandry is a group with one female and many males. Polyandry is a reproductive strategy that helps a female ensure reproductive success by providing her with multiple mating options.
Resource Defense Polyandry
In the Spotted Sandpiper, females control resources, which in turn controls male mating associations (Oring et al. 1994).
Cooperative Polyandry
The Galapagos hawk exhibits cooperative polyandry. In this case all males in the group copulate with the female and all participate in brood provisioning (Fabborg et al. 1995).
Polygynandry
Some mating systems have looser male-female bonds within groups. In polygynandrous groups, multiple females and males mate with each other, and males may care for the broods of several females. Chimpanzees and bonobos rely on this strategy — it allows groups of males and females to live together and spend less time being concerned with mate competition. Polygynandry may be advantageous from the female's perspective because it causes paternity confusion, which decreases infanticide and allows her to have multiple males care for her brood (Hrdy 1981, 2000).
Promiscuity
In promiscuity there are no pair bonds, and males and females, although sometimes choosy, often seem to mate randomly. As it is typically more advantageous for one or both sexes to pick their mate, promiscuity may occur in species for which the environment is unpredictable (Birkhead 2000, Burton 2002).
Sperm Competition
Although sperm competition is not a type of mating system per se, it is a form of male-male competition that plays an important role in mating systems. If more than one male mates with a female in a short time period, competition can occur after the males have released their sperm (Fisher & Hoekstra 2010). In other words, once a male has released sperm, its sperm must be the first to reach an egg. This is often apparent in animals that use external fertilization. In aquatic animals that release their gametes into the water, animals that release the largest amount of sperm, and sperm that are highly capable of swimming, are likely to produce the most offspring (Stoltz & Neff 2006). Animals with internal fertilization also experience sperm competition. Several mechanisms have evolved to facilitate a male's reproductive success with females that have multiple mates. For example, in one species of damselfly, males physically remove any sperm present from the female before it mates (Waage 1979). Sperm competition adds to the difficulty of obtaining a successful reproductive event by males.
Conclusions
To transfer their genes to the next generation successfully, animals need to choose a suitable mate. Failure to do so leads to low or no reproductive success — that is, poor fitness. But reproductive success can also hinge on the number of mates, and on social interactions that extend beyond mating. By classifying social interactions, scientists have been able to identify different types of mating systems, such as monogamy and polygyny. The mating systems described in this article represent a variety of strategies to achieve reproductive success. The diversity of mating systems in animals is a fascinating example of the incredible variety of solutions that a complex evolutionary problem can yield.
Glossary
Bateman's Principle: The theory that females almost always invest more energy into producing offspring than males, and therefore, in most species, females are a limiting resource over which the other sex will compete.
fitness: The relative measure of the reproductive success of an individual passing its genes to the next generation.
genetic recombination: The process of creating allelic variation in offspring by exchanging DNA; typically happens during sexual reproduction.
harem: A group of females associated with a single male. Typically the male in the harem defends his group of females.
lek: A mating system which consists of an aggregation of males where each is seeking to attract a mate. Leks are not associated with resources; however it is thought that leks attract more females than a single male would attract.
resource defense polygyny: A mating systems in which males establish a territory around resources needed for mating success. In this system multiple females will join the male in his territory.
polyandry: A mating system where one female pairs with many males.
polygyny: A mating system where one male is associated with many females.
promiscuity: A mating system where there are no pair bonds. In this case is seems that males and females mate randomly.
serial monogamy: A mating system in animals where they pair with a mate for one mating season but change mates over the course of a lifetime.
social monogamy: The behavioral pairing of a single female with a single male.
References and Recommended Reading
Aspbury, A. S. & Gibson, R. M. Long range visibility of greater sage-grouse leks: A GIS-based analysis. Animal Behaviour 67, 1128-1132 (2004).
Bateman, A. J. Inter-sexual selection in Drosophila. Heredity 2, 349-368 (1948).
Coleman, S. W. et al. Female preferences drive the evolution of mimetic accuracy in male sexual displays. Biological Letters 3, 463-466 (2007).
Beletsky, L. D. & Orians, G. H. Site fidelity and territorial movements of males in a rapidly declining population of yellow-headed blackbirds. Behavioral Ecology and Sociobiology 34, 257-265 (1994).
Birkhead, T. Promiscuity: An Evolutionary History of Sperm Competition. Cambridge, MA: Harvard University Press, 2000.
Burton, C. Microsatellite analysis of multiple paternity and male reproductive success in the promiscuous snowshoe hare. Canadian Journal of Zoology 80, 1948-1956 (2002).
Faaborg, J. et al. Confirmation of cooperative polyandry in the Galapagos hawk (Buteo galapagoensis). Behavioral Ecology and Sociobiology 36, 83-90 (1995)
Fisher, H. S. & Hoekstra, H. E. Competition drives cooperation among closely related sperm of deer mice. Nature 463, 801-803 (2010).
Grether, G. Intrasexual competition alone favors a sexually dimorphic ornament in the rubyspot damselfly Hetaerina americana. Evolution 50, 1949-1957 (1996).
Hauber, M.E. & Lacey, E.A. Bateman's principle in cooperatively breeding vertebrates: The effects of nonbreeding alloparents on variability in female and male reproductive success. Integrative and Comparative Biology 45, 903-914 (2005).
Hrdy, S. B. The Woman That Never Evolved. Cambridge, MA: Harvard University Press, 1981.
Hrdy, S. B. The optimal number of fathers: Evolution, demography, and history in the shaping of female mate preferences. Annals of the New York Academy of Sciences 907, 75-96 (2000).
Kellogg, K. A. et al. Intraspecific brood mixing and reduced polyandry in a maternal mouth-brooding cichlid. Behavioral Ecology 9, 309-312 (1998).
King, K. C. et al. The geographic mosaic of sex and the Red Queen. Current Biology 19, 1438-1441 (2009).
Lank, D. B. et al. Genetic polymorphism for alternative mating-behavior in lekking male ruff Philomachus pugnax. Nature 378, 59-62 (1995).Lande, R. Models of speciation by sexual selection on polygenic traits. Proceedings of the National Academy of Sciences of the United States of America 78, 3721-3725 (1976).
McCracken, G. F. & Bradbury, W. Social organization and kinship in the polygynous bat Phyllostomus hastatus. Behavioral Ecology and Sociobiology 8, 11-34 (1981).
Oring, L. W. et al. Mate acquisition tactics in polyandrous spotted sandpipers (Actitis macularia): The role of age and experience. Behavioral Ecology 5, 9-16 (1994).
Petrie, M. et al. The degree of extrapair paternity increases with genetic variability could be characterized as cryptic polyandry. Proceedings of the National Academy of Sciences of the United States of America 16, 9390-9395 (1998).
Rosenthal, G. G., & Evans, C. S. Female preference for swords in Xiphophorus helleri reflects a bias for large apparent size. Proceedings of the National Academy of Sciences of the United States of America 95, 4431-4436 (1998).
Simon, J. C. et al. Ecology and evolution of sex in aphids. Trends in Ecology and Evolution 17, 34-39 (2002).
Stoltz, J. A. & Neff, B. D. Sperm competition in a fish with external fertilization: The contribution of sperm number, speed and length. Journal of Evolutionary Biology 19, 1873-1881 (2006).
Stutchberry, B. J. M. Breeding synchrony best explains variation in extrapair mating system among avian species. Behavioral Ecology and Sociobiology 43, 221-222 (1998).
Waage, J. K. Dual function of the damselfly penis: Sperm removal and transfer. Science 203, 916-918 (1979).










