
Abstract
Anaerobic ammonium oxidation (anammox) is recognized as an important process for nitrogen cycling, yet little is known about its role in the subsurface biosphere. In this study, we investigated the presence, abundance, and role of anammox bacteria in upland soil cores from Tianjin, China (20 m depth) and Basel, Switzerland (10 m depth), using isotope-tracing techniques, (q)PCR assays, and 16 S rRNA & hzsB gene clone libraries, along with nutrient profiles of soil core samples. Anammox in the phreatic (water-saturated) zone contributed to 37.5–67.6% of the N-loss (up to 0.675 gN m−2d−1), with anammox activities of 0.005–0.74 nmolN g−1soil h−1, which were even higher than the denitrification rates. By contrast, no significant anammox was measured in the vadose zone. Higher anammox bacterial cell densities were observed (0.75–1.4 × 107copies g−1soil) in the phreatic zone, where ammonia-oxidizing bacteria (AOB) maybe the major source of nitrite for anammox bacteria. The anammox bacterial cells in soils of the vadose zone were all <103copies g−1soil. We suggest that the subsurface provides a favorable niche for anammox bacteria whose contribution to N cycling and groundwater nitrate removal seems considerably larger than previously known.
Similar content being viewed by others


Introduction
Nitrogen (N) is an essential nutrient for all organisms, and it is fundamental to the biochemical processes that define life. The global N cycle has historically been considered a “linear” process: N in the form of atmospheric nitrogen gas (N2) is fixed and transferred as ammonium (NH4+) to the Earth’s surface, oxidized to nitrate (NO3−), and finally reduced and returned back to the atmosphere as N2 gas. The various steps in this process are mediated by microbial activity1. Much has been learned about these steps and many of the microorganisms involved2,3. In today’s consideration of the N cycle, the surface ecosystem is still emphasized, while much less attention is paid to the subsurface biosphere4,5.
For decades, the conversion of fixed nitrogen to dinitrogen gas by heterotrophic bacteria, termed heterotrophic denitrification, was considered the main pathway of N loss in natural ecosystems6. The discovery of anaerobic ammonium oxidation (anammox) mediated by autotrophic anammox bacteria, which were able to oxidize ammonia directly to N2 without nitrous oxide (N2O) emission, greatly changed our view of the marine and terrestrial N cycles7,8,9. Anammox has been detected worldwide in natural ecosystems. In marine systems, it may be responsible for up to 50% of the marine N loss10,11, with hotspots occurring in oxygen minimum zones (OMZ)12,13,14. Anammox has also been detected in many freshwater systems with hotspots occurring in riparian sediments15,16,17,18. Overall, the available data indicates that anammox may be present in many environments, including surface aquatic ecosystems, and that anammox may be responsible for a significant proportion of N2 production. However, the prevalence of anammox in upland fields has not been identified yet19. We speculated that anammox would not occur significantly in the surface soil system but would occur in the subsurface biosphere, especially in the phreatic groundwater-saturated subsurface20,21.
Aquifer soils include soils in the vadose, phreatic, and aquitard zones. The phreatic zone stores groundwater, which plays a central part in drinking water production, agricultural irrigation, ecosystem sustenance, human adaptation to climate change, and global food security22,23. The contribution of anammox to N loss in groundwater was first observed in Massachusetts, USA using isotope tracers24. Subsequently, anammox in groundwater was investigated at several sites, such as Mansfield, UK25, Waterloo, Canada26, and in an in situ experiment at Cape Cod, MA, USA27 where the activity and contribution of anammox varied significantly due to groundwater mobility and allochthonous pollution. Interestingly, anammox in aquifer soils has not been examined to date. Prior studies have not determined whether anammox occurs only in fluent groundwater or whether it occurs in local aquifer soils, where the mechanism of anammox N loss appears different because of the interacting forms.
The aim of the present study was therefore to investigate the distribution, activity, contribution and microbial mechanism of anammox in soil profiles that represent both the vadose water-unsaturated zone and the phreatic zone of two unconfined aquifers in China and in Switzerland, in order to determine whether anammox is significant in subsurface environments. In addition, the potential interspecies relationships among anammox, nitrifiers (ammonia-oxidazing bacteria (AOB) and archaea (AOA)), and even denitrifying anoxic methane oxidizing population (n-DAMO, the fourth oxygen-producing biological pathway28) were also analyzed to find out whether these microorganism could form a novel pathway of anammox synergism in the subsurface biosphere. In this study, the term “aquifer” is defined as the phreatic water-saturated zone below the groundwater table of an unconfined aquifer, while the term “non-aquifer” is used to describe the vadose water-unsaturated zone above the groundwater table.
Results
Identification and quantification of anammox bacterial abundance in soil cores
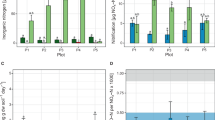
For the Tianjin and Basel core samples, the aquifers were located at a depth of 8–16 m and less than 7.2 m, respectively. The aquifer in Tianjin had characteristics of ammonium and nitrate pollution, containing up to 8.9 and 15.1 mg N kg−1, respectively. The NH4+ and NO3− concentrations in the Basel soil samples were all below 2 mg N kg−1 (Fig. 1). The presence of anammox bacteria was established by amplification of the hzsB and 16 S rRNA gene using primers specific for anammox. The positive PCR products of anammox bacterial gene were only obtained in aquifer soils. Samples from other depths generated no positive PCR results. The BLAST analysis of the gene sequences confirmed that all the clones represented anammox-like sequences.

The aquifer zone is indicated in light blue.
The anammox bacteria abundance was then estimated with qPCR targeting the hzsB gene, which is diagnostic for anammox bacteria. In Tianjin soil cores, the anammox numbers were all below 103 copies g−1 above the aquifer. A significant peak of anammox was detected in the aquifer (8–16 m bgl) where abundance ranged from 9.0 × 105 to 1.3 × 106 copies g−1 dry soil. Below the aquifer, the anammox abundance decreased to the detectable level (~103copies g−1).
The same trend was obtained in the Basel soil core. The peak of anammox bacteria was only distributed in the aquifer (7.2 m bgl) and ranged from 3 × 104 to 1.2 × 106copies g−1 dry soil. At other depths, no anammox abundance was detectable.
Activities, contribution, and role of anammox in the soil cores
The anammox activity and the potential role of anammox as a source of N loss in the soil cores were determined by performing incubations with homogenized soil under in situ temperatures using a 15N-labeled isotope-tracing technique. The depth profiles of Tianjin upland soils containing activated anammox were only in aquifer soils at rates of 0.23–0.74 nmol N g−1h−1 (Fig. 2I), which were in agreement with molecular results. Strikingly, some samples showed anammox rates for N loss that were even higher than those of denitrification (n = 5). The specific cellular anammox activities were then calculated based on total anammox activity and abundance. In the Tianjin aquifer, the specific anammox activity ranged from 4.5–16.0 fmol cell−1d−1 based on analysis of the anammox hzsB gene9, and this activity is at the upper end of reported values (2–20 fmol cell−1d−1 refs 8 and 29). Here, moisture content of soil samples seemed to be the key positive factor influencing anammox rates (r = 0.844, p = 0.000, Supplementary Table S2). The other samples of the Tianjin soil profile showed no significant anammox activity. In the Basel aquifer, the measured anammox ranged from 0.005–0.68 nmol N g−1h−1 with 37.5–58.3% N loss (Fig. 2II). At other depths, no significant anammox activity was measured. The specific activity ranged from 4.5–13.3 fmol cell−1d−1. These data indicate that denitrification is not the only significant pathway for N loss in the aquifer. In the Basel aquifer samples, the water content also had a significant positive correlation with the anammox rate (r = 0.826, p = 0.000, Supplementary Table S3).

The aquifer zone is indicated in light blue. ‘ND’ indicates the abundance of anammox below the detection limit (<103copies g−1), meanwhile, no positive rate of anammox was detected.
Based on the anammox rates and bulk density of all soils (2.6–2.8 g cm−3), the N loss flux attributed to the anammox in the aquifers was estimated. In the soil core samples from the Tianjin aquifer, the N loss via anammox was 0.214–0.675 g N m−2d−1, while the contribution of denitrification to the N loss was 0.187–0.408 g N m−2d−1. It indicated that the anammox contributed a significant percentage (41–67%) to microbial N2 production and N removal in the aquifer. In the soil core samples from the Basel site, the N loss via anammox was 0.005–0.634 g N m−2d−1, while the contribution of denitrification to the N loss was 0.008–0.453 g N m−2d−1. These data indicate that anammox is an important but overlooked alternative process to denitrification for microbial N loss.
Biodiversity and community of anammox bacteria in the aquifers
The biodiversity and community composition of anammox bacteria in the aquifers were investigated by analyzing the hzsB and 16 S rRNA gene sequences retrieved from clone libraries using rarefaction analysis, the Chao1 estimator, and Shannon index calculations (Fig. 3). Phylogenetic analyses of anammox bacterial hzsB and 16 S rRNA gene sequences and related sequences deposited in the GenBank showed that all sequences obtained from soils from the Basel aquifer and Tianjin aquifer were most closely related to “Candidatus Brocadia” (similarity 94.6–97.6%) (Fig. 3I). The analyses of the hzsB and 16 S rRNA gene sequences showed that a total of 10 and 4 OTUs, respectively, were obtained from the Basel aquifer based on the hzsB (98% cut-off) and 16 S rRNA gene sequences (97% cut-off). Nevertheless, the anammox bacteria from the Tianjin aquifer showed a very limited anammox bacterial diversity with a total of 6 and 2 OTUs respectively obtained for the hzsB and 16 S rRNA gene sequences. The hzsB and 16 S rRNA gene sequences both showed a higher biodiversity in soil samples from the Basel aquifer (with little human activity and good groundwater quality) than from the Tianjin aquifer (Fig. 3II).

An evolutionary distance dendrogram (constructed by the neighbor-joining method using the maximum composite likelihood distance with a 1,000 bootstrap in a MEGA 4.0 software package) shows the affiliations of the anammox bacterial hzsB gene sequences (above) and the 16 S rRNA gene sequences (below) retrieved from the aquifer soils in Tianjin and Basel. The OTUs are identified in bold with the number of sequences identified in brackets from the Tianjin aquifer (41 hzsB and 23 16 S rRNA gene sequences in total) and the Basel aquifer (53 hzsB and 43 16 S rRNA gene sequences in total). The known anammox bacterial sequences are identified with solid circles. The designators [J], [K], [S], and [A] indicate the anammox bacteria “Jettenia,” “Kuenenia,” “Scalindua,” and “Anammoxoglobus,” respectively. The details for the anammox clone sequences of each OTU and for the accession numbers in the Genbank are listed in Supplementary Tables S3 and S4. The DOTUR program was used with 2% sequence variation (for hzsB gene sequences) and 3% sequence variation (for 16 S rRNA gene sequences) for OTU determination.
Quantitative analysis of related N cycle processes with anammox in aquifer soils
The biogeochemical mechanism of anammox in the aquifer was investigated in the Tianjin soil cores, and the potential rates of nitrification and denitrification were determined to assess the source of nitrite for anammox. The nitrification and denitrification rates decreased from the surface to the aquifer, and denitrification rates (0.2–0.47 nmol N g−1h−1) were all higher than nitrification rates (0.02–0.13 nmol N g−1h−1) in the vadose zone (Fig. 4). This situation was completely reversed in the aquifer, where both the nitrification and denitrification rates increased, and nitrification rates (0.29–0.66 nmol N g−1h−1) were all higher than denitrification rates (0.21–0.45 nmol N g−1h−1). Therefore, in the aquifer, both partial denitrification and nitrification may provide anammox with nitrite, but nitrification may be the main nitrite source for anammox.

The percentages of nitrification to denitrification and ammonia-oxidizing archaea to ammonia-oxidizing bacteria are shown in the right-hand columns of panel a and panel b, respectively.
The potential of a combined nitrification–anammox process in aquifer soils was further explored by performing qPCR assays on bacterial and archaeal amoA genes in the soil core (0–20 m). The qPCR data for the soils above the aquifer showed higher gene numbers for the archaeal amoA gene (6.7 × 104–2.2 × 106 copies g−1 dry soil) than for the bacterial amoA copies (2.6 × 103–1.1 × 106copies g−1 dry soil). However, in the aquifer, the abundance was higher for AOB (4.4 × 104–9.1 × 104copies g−1) than for AOA (7.1 × 103–2.8 × 104copies g−1 dry soil). Multiple linear stepwise regression analysis also identified the AOB abundance to be the most determining variable regarding nitrification activity (Supplementary Table S5). Furthermore, the n-DAMO bacteria, which conduct the fourth biological pathway that produces oxygen28, were also detected in the samples where AOA, AOB, and anammox bacteria were present. A potential interaction among AOA (‘Nitrososphaera’ dominant), AOB (‘Nitrosomonas’ dominant), n-DAMO (‘Methylomirabilis’), and anammox bacteria (Brocadia) would form a new pathway of anammox synergism in the subsurface biosphere (Fig. 5, Supplementary Figure S4).

The affiliation of AOA&AOB amoA gene and n-DAMO pmoA gene sequences were shown in Supplementary Figure S3.
Discussion
In the present study, the activity, contribution, and role of anammox in subsurface deep upland soils were investigated. Results showed that anammox spieces in the upland soil cores were low biodiversity but highly region-heterogeneous. Below the groundwater level, anammox bacterial abundance and activity were detected at all depths of each soil core. In contrast, above the groundwater level, anammox bacterial abundance was below the detection limit (~103copies g−1). Anammox in the phreatic zone contributed 37.5–67.6% of the N loss with activities of 0.005–0.74 nmol N g−1soil h−1 that were even higher than the denitrification rates (0.337 ± 0.092 nmol N g−1soil h−1, t - test, p = 0.000). The discovery indicate that anammox process actually occurred in local aquifer soils, and significantly contribute in subsurface soil biospheres, especially in the aquifers zone.
The anammox process showed distinct biogeochemical features in aquifer soils and in groundwater. Anammox activity has been reported in groundwater at several sites24,27,29,30. Due to groundwater mobility and allochthonous pollution, the activity and contribution of anammox in groundwater were significantly different at each site. Böhlke et al.24 first detected anammox in groundwater with isotope tracers, with a rate of up to 0.027 μmol L−1d−1, indicating that anammox played only a minor role in the N cycle. Clark et al.26 also provided 15N evidence for anammox in anoxic groundwater, and Moore et al.30 reported bacterial activity of anammox at study sites where groundwater was contaminated by ammonium, with a contribution of 18–36% N loss. In a recent report by Smith et al.25, anammox was found to contribute 39−90% of potential N2 production in wastewater-derived contaminated groundwater. These results suggest the possibility of site-specific heterogeneity in anammox bacterial distributions in groundwater systems. In contrast to the fluidity of groundwater, aquifer soils were stationary, and anammox in aquifers also showed much less fluctuations. In Tianjin aquifer soils, anammox rates were stable at 0.49 ± 0.21 nmol N g−1soil h−1 (t - test, p = 0.000) and contributed 41−67% of the potential N2 production with little variation.
While a high anammox bacterial diversity was reported for groundwater, the aquifers soils of our study exhibited an almost uniform pattern. Anammox bacteria, “Candidatus Brocadia anammoxidans” and “Candidatus Kuenenia stuttgartiensis,” were detected at an ammonium-contaminated site in Mansfield, UK31. In wastewater-derived contaminated groundwater (USA), Brocadia-like bacteria and “Ca. Kuenenia spp.” were also detected27. Most interestingly, in the ammonium-contaminated aquifer, four of the five known genera of anammox sequences were present, i.e. “Ca. Scalindua,” “Ca. Jettenia,” “Ca. Kuenenia,” and “Ca. Brocadia” genus30. Yet in aquifer soils, only one genus, “Candidatus Brocadia,” was identified in both Tianjin and Basel aquifer soils based on phylogenetic analysis of the anammox bacterial 16 S rRNA and hzsB genes. The anammox bacterial biodiversity in the subsurface biosphere was significantly different from that commonly found in the surface biosphere. In surface aquatic systems, the anammox bacteria consistently showed a very low biodiversity11,32. For example, only bacteria of the “Scalindua” genus were detected in marine ecosystems, while “Candidatus Scalindua brodae”’ and “Candidatus Brocadia anammoxidans” were detected in Lake Tanganyika33 and river sediments17, respectively. In wastewater treatment systems, the biodiversity was also low34. In contrast, different anammox genera co-existed in various wetland soils ecosystems. In paddy field soils, four genera of anammox bacteria, “Brocadia,” “Kuenenia,” “Anammoxoglobus,” and “Jettenia” were detected21. In addition, a high biodiversity was observed in various soil ecosystems, such as peat soil, permafrost soil, and agricultural soil19,35. Hence, the mechanism of anammox N loss in local aquifer soil appears to be different from that of fluent groundwater and needs further investigation.
In contrast to numerous reports on the widespread distribution of anammox in aquatic ecosystems, there has been no report about the prevalence of anammox in surface upland fields19 until now. Hence, in the present study, we investigated anammox in deep upland soils. Both the molecular and isotope results showed that anammox did not occur in upland surface soils, but was only mediated in water logged aquifer soils. In aquifer soils, the anammox abundance (above 105copies g−1 soil) and rate (above 0.01 nmol N g−1soil h−1) were significantly above the detection limit compared with the non-aquifer zone where anammox bacterial abundance and rate were undetectable (below 103copies g−1soil). In the Tianjin aquifer, the increases in NH4+ and NOx− were considerably higher than those in non-aquifer soils and were also higher than those in Basel soils. However, the biogeochemical correlation analysis showed that the substrate (ammonia or nitrate) had little positive influence on anammox rates in either the Tianjin or Basel aquifers. In the Basel aquifer, by contrast, the NH4+ and NOx− levels were as low as 1–2 mg kg−1, and were at the same levels as in the non- aquifer samples. Biogeochemical correlation analysis showed that the water contents had the most positive influence on anammox abundance in the Basel aquifer (r = 0.977, p = 0.000). Therefore, the substrate contents may not be the key limiting factors for anammox, while water contents showed a positive relation with anammox occurrence. Our previous studies demonstrated that ubiquitous anammox occurs in freshwater at various levels of substrate concentration18, indicating that water content controls are important for the anammox reactions. Previous studies, including ours, have clearly shown that abundant anammox bacterial cells exist in upland soils after flooding where no anammox bacteria were detected before flooding18,36. It may due to higher material mobility and electron transport caused by water. We speculated, therefore, that as the direct substrate of anammox bacteria, ammonia and nitrate contents in soils were the most influential factors determining anammox rate, whereas the water content was the key factor controlling anammox occurrence. After oxygen was consumed in the water-limited diffusion process, an potential anoxic niche for anammox bacteria was created. This was likely responsible for the observed patterns of occurrence, which need to be further investigated.
The discovery of anammox in aquifer soils provides a new perspective on the N cycle and groundwater N pollution. Since the 1970 s, NO3− contamination (>10 mg L−1, USA EPA) of groundwater has become a significant environmental problem with many parts of the world now reporting groundwater nitrate pollution37. Denitrification has been emphasized as the dominant nitrate attenuation process in the phreatic water-saturated zone with minor roles played by other nitrate depletion mechanisms, such as dissimilatory nitrate reduction to ammonium (DNRA), assimilation of nitrate into microbial biomass, and nitrate removal via phreatophytes38. In the present study, our use of isotopic tracing technology revealed that up to 37.5–67.6% of aquifer soil N2 production, with rates of 0.005–0.74 nmol N g−1soil h−1, was contributed by anammox. At some depths in the aquifer soils, we found that anammox contributed even more than denitrification. The different metabolic pathways in anammox and denitrifying bacteria may give rise to different contributions of anammox and denitrification to nitrate removal. Denitrifiers were mostly facultative anaerobic heterotrophs, which obtain both their energy and carbon from the oxidation of organic compounds39. Oxic aquifers often contain low concentrations of dissolved organic matter37, limiting the energy needed by denitrifying bacteria that use organic carbon as the electron donor. However, the anammox process is catalyzed by the autotrophic anammox bacteria40. Hence, when organic compounds were limited, the anammox bacteria were favored, as has been widely documented in anammox bioreactor research3,20,39. An estimated 0.005–0.675 g N m−3d−1 was lost via anammox in the aquifer soils, indicating that the anammox process is an effective and feasible way to counteract nitrate pollution in groundwater.
Potential interactions have been suggested between particle-associated anammox bacteria and archaeal partners with n-DAMO bacteria in the deep soils. In these oxygen-limited aggregates, cooperation between AOA and AOB can provide nitrite to anammox bacteria39,41 in a partnership similar to those reported for marine snow particles13,14,42. Simultaneous nitrite-dependent anaerobic methane and ammonium oxidation processes have also been reported in various bioreactors with efficient nitrogen removal loadings and rates43,44. Although the oxygen production rates by damo bacteria were still not available, the affinity (Ks) value of AOA was very low (133 nM45), and AOA would be operative at low oxygen to oxidize the ammonia to nitrite for anammox bacteria. The coexistence of ammonia-oxidizing archaea, ammonia-oxidizing bacteria, n-DAMO bacteria, and anammox has been reported in populations ranging from the lab scale41 to the natural systems16.
Methods and Materials
Soil Samples and Background
Two kinds of unconfined aquifers were selected as sampling sites: a Tianjin aquifer in China (N40°07′ E116°28′), which experiences human activity disturbance, and a Basel aquifer in Switzerland (N47°31′ E7°43′), which has little human disturbance. The Tianjin aquifer is located in the North China Plain, which plays a central role in China’s food production by supplying more than half of China’s wheat and one-third of its maize. Agricultural production in the region has grown markedly in the past decades, strongly benefiting from the fast-growing groundwater exploitation. The strong increase in the regional evapotranspiration has led to an imbalance in the groundwater budget46. Moreover, the nitrate pollution here is becoming another difficult issue. By contrast, the Basel aquifer, located in the Canton Basel-Landschaft of northwest Switzerland, has groundwater of drinking water quality with little to moderate human activity near the sampling site.
Depth-dependent soil cores were collected using PowerProbe (at the Tianjin site) and GeoProbe direct push drill rigs (at the Basel site) in June 2013. The aquifer of the Tianjin sampling site is between 8 and 16 m below ground level (bgl) with overlying permeable, but unsaturated, soils. Three parallel soil cores (0–20 m bgl) were collected and sectioned at 1 m intervals based on the model of PowerProbe drill rigs. The aquifer of the Basel sampling site is below 7.2 m bgl, and three parallel soil cores (0–10 m bgl) were collected and sectioned at 1.2 m intervals based on the model of GeoProbe drill rigs. The collected soil cores were sealed and transported to the laboratory at 4 °C as soon as possible. One part was immediately incubated to determine anammox activities, and another part was sieved through 0.5 mm for chemical component analysis; subsamples were stored at −80 °C for DNA extraction and subsequent molecular analysis. The groundwater samples for incubation media were also collected, and measured for dissolved oxygen (DO) concentration (Fig. 1I) and in situ temperatures (T, 13 ± 2 °C and 10 ± 2 °C in Tianjin and Basel aquifers, respectively), sterilized through 0.22 μm filter, and finally stored at 4 °C.
DNA Extraction, PCR, Cloning, and Sequencing Analysis
About 0.35 g of freeze-dried soil from each sample (20 in Tianjin and 8 in Basel) was used for DNA extraction using a FastDNA SPIN Kit for Soil (Bio 101, USA), following the manufacturer’s protocol. Each sample was determined reparably three times. The extracted DNA was checked on 1% agarose gel, and the concentration was determined with Nanodrop ND-1000 ultraviolet-visible spectrophotometry (NanoDrop Technologies, Wilmington, DE, USA). Polymerase chain reaction (PCR) was performed in a C1000 thermal cycler (BioRad, USA). The function/16 S rRNA genes of anammox and related microorganism used in this study was following: hydrazine synthase β-subunit (hzsB) and 16 S rRNA gene of anammox bacteria, ammonia monooxygenase α-subunit (amoA) gene of AOB & AOA, and particulate methane monooxygenase α-subunit (pmoA) gene of n-DAMO. The details of primers and corresponding reaction profiles are shown in Table S1. The PCR product was gel-purified and ligated using pGEM-T Easy Vector (Promega, USA). The resulting ligation products were used to transform Escherichia coli JM109 competent cells, following the manufacturer’s instructions. In total, 96 clones were picked for each of the PCR products from each sampling site. PCR screens for the presence of inserts were performed using T7 and SP6 vector primers, and the amplicons were analyzed with restriction endonuclease Hha I, Hae III, and Rsa I (TAKARA, Dalian, China). Restriction digestion was carried out in a total volume of 20 μL that included 5 U restriction enzymes and 4 μL PCR products, and the system was incubated for 2 h at 37 °C. Digested DNA fragments were analyzed by fragment separation on a 2% (w/v) agarose gel and visualized with a GBOX/HR-E-M (Syngene, UK). Representative clones from each digestion pattern were selected for sequencing using an ABI 3730XL automated sequencer (Applied Biosystems, USA). Searches using the Basic Local Alignment Search Tool (BLAST) from the National Center for Biotechnology Information (NCBI) against the GenBank database verified that the PCR products were most closely related to the target sequences. All the sequences and their relatives obtained from the NCBI-BLAST were aligned using the Clustal X1.83 program47. The anammox bacterial sequences that shared 97% nucleotide similarity for 16 S rRNA and 98% nucleotide similarity for the hzsB gene were grouped into the same operational taxonomic unit (OTU) by employing the furthest neighbor approach using DOTUR software48. Biodiversity indicators (Shannon and Chao 1) were also calculated with DOTUR software. Phylogenetic trees were constructed using the neighbor-joining (NJ) mode and the maximum composite likelihood mode of the MEGA 4 software package49. The sequences obtained in this study are available at the NCBI under the Accession numbers of KF905049-KF905142 (anammox hzsB gene), GU083969-GU084010 and KF896206–KF896228 (anammox 16 S rRNA gene), KP168045–KP168078, HQ202453, and KC341050 (AOA amoA gene), KC341292–KC341300 (n-DAMO pmoA gene), and KT005575–KT005589 (AOB amoA gene).
Quantitative Real-Time PCR
SYBR-Green I- based real-time qPCR assays were carried out in a volume of 20 μL containing 10 μL SYBR Premix Ex Taq (TAKARA, Dalian, China), 4 pmol of each primer, and 2 μL of ten-fold diluted DNA template. Each sample was determined reparably three times. Amplification and detection were carried out with an ABI Prism 7300 Sequence Detection System (Applied Biosystems, USA) with the primer set and thermal profiles compiled in Supplementary Table S1. The plasmid DNAs were extracted with a GeneJet Plasmid Miniprep Kit (Fermentas, Lithuania), and concentrations were determined on a Nanodrops ND-1000 UV-Vis Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) for calculations of hzsB and amoA gene copy numbers. Ten-fold serial dilutions of a known copy number of the plasmid DNA were subjected to real-time qPCR in triplicate to generate an external standard curve. Melting curves were generated after each assay to check the specificity of amplification. PCR efficiencies were 90–103% (average = 92%) for the anammox bacterial hzsB and the archaeal and bacterial amoA genes. Only the results with correlation coefficients above 0.98 were used.
In the real-time qPCR quantitative assays targeting the hzsB gene, the detection limit of environmental samples was determined by a dilution method18. With the identical PCR procedure, the lowest anammox abundance was observed with the undiluted sample of CZ29-4 at ~103 copies g−1, which was the assumed detection limit in this environmental investigation (Supplementary Fig. S2).
Measuring rates of anammox, denitrification and nitrification with 15N-tracer technique
Different 15N- amended substrates were added for the rate determinations of various N-cycling processes. The homogenized soil samples with known weight and bulk density were transferred to helium-flushed, 12-mL glass vials (Exetainer, Labco, UK) together with in situ media water at in situ soil temperature. Triplicate activity tests were performed for each soil sample. For anammox and denitrification, the resulting slurries were pre-incubated under anoxic conditions to remove residual nitrite/nitrate (NOx−) in soils and incubation media according to the established protocol17. It is emphasized that, because of the different soil sample backgrounds, the pre-incubated times to remove the residual NOx− varied in a wide range, from 45 h to several days, in order to ensure completely anoxic conditions in the soils slurries, which was important for the appropriate controls. Subsequently, 100 μL of N2-purged stock solution of each isotopic mixture was added to obtain a final concentration of about 10% of the initial content background to avoid overestimate. The isotopic mixtures were as follows: (1) 15NH4+ (15N at 99.6%, negative control), (2) 15NH4+ + 14NO3− (positive control) and (3) 15NO3− (15N at 99%, for calculations). Incubation of the slurries was stopped at 0 h, 3 h, 6 h, 12 h, 24 h by 200 μL 7 M ZnCl2 injected. The slurries amended solely with 15NH4+ showed no significant accumulation of 15N2-labeled gas (29N2 and/or 30N2) in any sample, indicating that all ambient 14NOx− had been consumed during the pre-incubations. When both 15NH4+ and 14NO3− were added, 29N2 accumulated in every soil sample with no accumulation of 30N2. This pattern was reproducible and indicated that the anammox process was detectable in the soils. Slurries amended solely with 15NO3− showed significant anammox and denitrification rates. For nitrification process, the rate was measured as the production of 15NO2− from incubations with 15NH4+ (100 μL injection) via the reduction of NO2− by sulfuric acid to N2 chemically. A 12 h or longer incubation was needed to achieve better NO2− reduction efficiency, as described by ref. 50. All samples were incubated with 15N or 14N compounds to a final concentration that corresponded to a maximum of 10% of the in situ concentrations. The possibility of overestimating the potential rate measurements was therefore minimized. Three parallel assays were conducted for each soil sample. Process rates were calculated from produced 29N2 or 30N2 as measured by gas chromatography-isotope ratio mass spectrometers (GasBench-II, Elemental Analyzer FLASH 2000 HT, MAT 253, Germany).
Analytical procedures of environmental variables
The environmental variables of soils, including NH4+, NOx−, total nitrogen (TN), total phosphorus (TP), total organic matter (TOM), pH, and moisture content, were investigated51. A mixture of 5 g of sieved fresh soil sample and 12.5 mL of ddH2O (a soil-to-water ratio of 1:2.5) was measured for pH value. Both NH4+-N and NOx−-N were extracted from 6 g of sieved soil by shaking for 1 h at room temperature with 30 mL 2 M KCl, and were then measured spectrophotometrically. The TN was determined by potassium persulfate digestion and measured spectrophotometrically. The TOM was measured using the potassium dichromate method. The DO and T in groundwater were monitored in-situ using a WTW oxi/340i oxygen probe (WTW Company, Weilheim, Germany). Triplicates were run for quality assurance (QA).
Statistical analysis
The statistical analyses were conducted using SPSS Statistics 18.0 software (Predictive Analytics Software Statistics). The correlations between various variables were computed by Spearman correlation analysis. Multiple linear regressions (stepwise regression) were used to identify the most determining variables for the dependent variables. Unless otherwise specified, the level of significance in this study was α ≤ 0.05.
Additional Information
How to cite this article: Wang, S. et al. Nitrogen loss by anaerobic ammonium oxidation in unconfined aquifers soils. Sci. Rep. 7, 40173; doi: 10.1038/srep40173 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Falkowski, P. G., Fenchel, T. & Delong, E. F. The microbial engines that drive earth’s biogeochemical cycles. Science 320, 1034–1039 (2008).
Francis, C. A., Beman, J. M. & Kuypers, M. M. New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation. ISME J. 1, 19–27 (2007).
Jetten, M. S. et al. The anaerobic oxidation of ammonium. FEMS Microbiol. Rev. 22, 421–437 (1998).
Canfield, D. E., Glazer, A. N. & Falkowski, P. G. The evolution and future of earth’s nitrogen cycle. Science 330, 192–196 (2010).
Galloway, J. N. et al. Nitrogen cycles: past, present, and future. Biogeochemistry 70, 153–226 (2004).
Burgin, A. J. & Hamilton, S. K. Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front Ecol. Environ. 5, 89–96 (2007).
Dalsgaard, T., Canfield, D. E., Petersen, J., Thamdrup, B. & Acuňa-González, J. N2 production by the anammox reaction in the anoxic water of Golfo Dulce, Costa Rica. Nature 422, 606–608 (2003).
Kuypers, M. M. M. et al. Anaerobic ammonium oxidation by anammox bacteria in Black Sea. Nature 422, 608–611 (2003).
Kartal, B. et al. Molecular mechanism of anaerobic ammonium oxidation. Nature 479, 127–130 (2011).
Arrigo, K. R. Marine microorganisms and global nutrient cycles. Nature 437, 349–355 (2005).
Schmid, M. C. et al. Anaerobic ammonium-oxidizing bacteria in marine environments: widespread occurrence but low diversity. Environ. Microbiol. 9, 1476–1484 (2007).
Kuypers, M. M. M. et al. Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation. Proc. Natl. Acad. Sci. USA 102, 6478–6483 (2005).
Lam, P. et al. Linking crenarchaeal and bacterial nitrification to anammox in the Black Sea. Proc. Natl. Acad. Sci. USA 102, 6478–6483 (2007).
Lam, P. et al. Revising the nitrogen cycle in the Peruvian oxygen minimum zone. Proc. Natl. Acad. Sci. USA 106, 4752–4757 (2009).
Nie, S. et al. Nitrogen loss by anaerobic oxidation ammonium in rice rhizosphere. ISME J. 9, 2059–67 (2015).
Wang, S., Zhu, G., Peng, Y., Jetten, M. S. M. & Yin, C. Anammox bacterial abundance, activity and contribution in riparian sediments of the Pearl River Estuary. Environ. Sci. Technol. 46, 8834–8842 (2012).
Zhu, G. et al. Hotspots of anaerobic ammonia oxidation at land-freshwater interfaces. Nature Geosci. 6, 103–107 (2013).
Zhu, G. et al. Ubiquitous anaerobic ammonium oxidation in inland waters: an overlooked nitrous oxide mitigation process. Sci. Rep. 5, doi: 10.1038/srep17306 (2015).
Humbert, S. et al. Molecular detection of anammox bacteria in terrestrial ecosystems: distribution and diversity. ISME J. 4, 450–454 (2010).
Zhu, G., Jetten, M. S. M., Kuschk, P., Ettwig, K. & Yin, C. Potential roles of anaerobic ammonia and methane oxidation in the nitrogen cycle of wetland ecosystems. Appl. Microbiol. Biotechnol. 86, 1043–1055 (2010).
Zhu, G. et al. Anaerobic ammonia oxidation in a fertilized paddy soil. ISME J. 5, 1905–1912 (2011b).
Aeschbach-Hertig, W. & Gleeson, T. Regional strategies for the accelerating global problem of groundwater depletion. Nat. Geosci. 5, 853–861 (2012).
Taylor, R. G. et al. Ground water and climate change. Nat. Clim. Chang. 3, 322–329 (2013).
Böhlke, J. K., Smith, R. L. & Miller, D. N. Ammonium transport and reaction in contaminated groundwater: Application of isotope tracers and isotope fractionation studies. Water Resour. Res. 42, 1–19 (2006).
Smits, T. H. M., Huttmann, A., Lerner, D. N. & Holliger, C. Detection and quantification of bacteria involved in aerobic and anaerobic ammonium oxidation in an ammonium-contaminated aquifer. Biorem. J. 13, 41–51 (2009).
Clark, I. et al. Origin and fate of industrial ammonium in anoxic ground water—15N evidence for anaerobic oxidation (Anammox). Ground Water Monit. R. 28, 73–82 (2008).
Smith, R. L., Böhlke, J. K., Song, B. & Tobias, C. Role of anaerobic ammonium oxidation (anammox) in nitrogen removal from a freshwater aquifer. Environ. Sci. Technol. 49, 12169–77 (2015).
Ettwig, K. F. et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 456, 543–560 (2010).
Strous, M. et al. Missing lithotroph identified as new planctomycete. Nature 400, 446–449 (1999).
Moore, T. A. et al. Prevalence of anaerobic ammonia-oxidizing bacteria in contaminated groundwater. Environ. Sci. Technol. 45, 7217–7225 (2011).
Smith, R. L. Is anaerobic ammonium oxidation (“anammox”) important for nitrogen cycling in groundwater? 125th Anniversary Annual Meeting & Expo (2013).
Jewell, T. N. M. et al. Metatranscriptomic analysis of groundwater reveals an active anammox bacterial population. AGU Fall Meeting Abstracts (2014).
Schubert, C. J. et al. Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika). Environ. Microbiol. 8, 1857–63 (2006).
Wang, S. et al. Anaerobic ammonium oxidation in traditional municipal wastewater treatment plants with low-strength ammonium loading: widespread but overlooked. Water Res. 84, 66–75 (2015).
Hu, B. L. et al. New anaerobic, ammonium-oxidizing community enriched from peat soil. Appl. Environ. Microbiol. 77, 966–71 (2011).
Hu, B. et al. Enrichment of an anammox bacterial community from a flooded paddy soil. Environ. Microbiol. Rep. 5, 483–489 (2013).
Rivett, M. O., Buss, S. R., Morgan, P., Smith, J. W. & Bemment, C. D. Nitrate attenuation in groundwater: a review of biogeochemical controlling processes. Water Res. 42, 4215–4232 (2008).
Schmidt, S. I. & Jürgen Hahn, H. What is groundwater and what does this mean to fauna? Limnologica 42, 1–6 (2012).
Zhu, G. et al. Biological removal of nitrogen from wastewater. In Reviews of environmental contamination and toxicology Springer, New York, 159–195 (2008).
Kartal, B. et al. Anammox-growth physiology, cell biology, and metabolism. Adv. Microb. Physiol. 60, 211–262 (2012).
Yan, J. et al. Mimicking the oxygen minimum zones: stimulating interaction of aerobic archaeal and anaerobic bacterial ammonia oxidizers in a laboratory-scale model system. Environ. Microbiol. 14, 3146–3158 (2012).
Woebken, D., Fuchs, B. M., Kuypers, M. M. M. & Amann, R. Potential Interactions of particle-associated anammox bacteria with bacterial and archaeal partners in the Namibian Upwelling System. Appl. Environ. Microbiol. 73, 4648–4657 (2007).
Luesken, F. A. et al. Simultaneous nitrite-dependent anaerobic methane and ammonium oxidation processes. Appl. Environ. Microbiol. 77, 6802–6807 (2011).
Shi, Y. et al. Nitrogen removal from wastewater by coupling anammox and methane-dependent denitrification in a membrane biofilm reactor. Environ. Sci. Technol. 47, 11577–83 (2013).
Martens-Habbena, W., Berube, P. M., Urakawa, H., de la Torre, J. R. & Stahl, D. A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 461, 976–979 (2009).
Gleeson, T., Wada, Y., Bierkens, M. F. P. & van Beek, L. P. H. Water balance of global aquifers revealed by groundwater footprint. Nature 488, 197–200 (2012).
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. & Higgins, D. G. The CLUSTALX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25, 4876–4882 (1997).
Schloss, P. D. & Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 71, 1501–1506 (2005).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739 (2011).
Füssel, J. et al. Nitrite oxidation in the Namibian oxygen minimum zone. ISME J. 6, 1200–1209 (2012).
Bao, S. D. Chemical Analysis for Agricultural Soil China Agriculture Press, Beijing (2000).
Acknowledgements
This research is financially supported by the National Natural Science Foundation of China (No. 41322012, 41671471, and 21277156), Strategic Priority Research Program of the Chinese Academy of Sciences (XDB15020303), and a special fund of the State Key Joint Laboratory of Environment Simulation and Pollution Control (14L01ESPC). Moreover, the author Guibing Zhu gratefully acknowledges the support of Humboldt Research Fellowship (1152633) and Youth Innovation Promotion Association of CAS. Mike Jetten is supported by ERC AG 232937 and 339880.
Author information
Authors and Affiliations
Contributions
S.W., S.H., L.Z., S.Z. and G.Z. performed research; S.W., D.R., L.Z., S.Z., and G.Z. analyzed data; S.W. and G.Z. wrote the paper and all co-authors substantially contributed to commenting and revising it. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, S., Radny, D., Huang, S. et al. Nitrogen loss by anaerobic ammonium oxidation in unconfined aquifer soils. Sci Rep 7, 40173 (2017). https://doi.org/10.1038/srep40173
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40173
This article is cited by
-
Nitrogen fixation and diazotroph diversity in groundwater systems
The ISME Journal (2023)
-
Dissimilatory nitrate reduction and functional genes in two subtropical rivers, China
Environmental Science and Pollution Research (2021)
-
Resuscitation of anammox bacteria after >10,000 years of dormancy
The ISME Journal (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
