
Abstract
Transposition mutagenesis is a powerful tool to identify the function of genes, reveal essential genes and generally to unravel the genetic basis of living organisms. However, transposon-mediated mutagenesis has only been successfully applied to a limited number of archaeal species and has never been reported in Thermococcales. Here, we report random insertion mutagenesis in the hyperthermophilic archaeon Pyrococcus furiosus. The strategy takes advantage of the natural transformability of derivatives of the P. furiosus COM1 strain and of in vitro Mariner-based transposition. A transposon bearing a genetic marker is randomly transposed in vitro in genomic DNA that is then used for natural transformation of P. furiosus. A small-scale transposition reaction routinely generates several hundred and up to two thousands transformants. Southern analysis and sequencing showed that the obtained mutants contain a single and random genomic insertion. Polyploidy has been reported in Thermococcales and P. furiosus is suspected of being polyploid. Yet, about half of the mutants obtained on the first selection are homozygous for the transposon insertion. Two rounds of isolation on selective medium were sufficient to obtain gene conversion in initially heterozygous mutants. This transposition mutagenesis strategy will greatly facilitate functional exploration of the Thermococcales genomes.
Similar content being viewed by others

Introduction
It is now well established that Archaea represent a taxonomic domain distinct from Bacteria, their prokaryotic counterpart. Recent phylogenetic analysis even suggest a Bacteria-Archaea two domain tree of life in which Eukaryotes found their origin within the Archaeal domain1,2. Even early in the discovery of Archaea, phenotypic properties distinguished them from Bacteria while suggesting similarity with eukaryotes. Indeed, Archaea were found to be insensitive to most 70S-targeted drugs such as streptomycin, but were sensitive to 80S-targeted drugs such as anisomycin3. Further studies confirmed that Archaea shared many features with eukarya, including their ribosomal proteins4. From a practical standpoint, isolated archaeal macromolecular systems, such as the ribosome, are simpler and easier to study than their eucaryotic counterparts. Thus, Archaea have allowed faster understanding of many eucaryal processes4. Far from being simplistic versions of eukaryotes, Archaea have very unique features, linked to their ability to thrive at the extremes of pH, temperature, salinity and hydrostatic pressures. These properties are biologically and biochemically intriguing and makes Archaea one of the most important resources for biotechnological and industrial applications5,6. However, this resource remains difficult to access even in the age of genomics. Archaeal genomes are predominantly annotated from the knowledge of bacterial and eukaryal processes. As a result, there is a high fraction of genes for which no biochemical function can be assigned, and for instance can exceed 60% in some families such as the Thermococcales7,8,9.
Functional exploration of archaeal organisms remains limited by the lack of proper genetic tools. This is particularly evident in extremophiles whose extreme growth conditions prevent the use of genetic tools developed in mesophilic organisms and also render commonly used selective agents ineffective10. The discovery of the first archaeal plasmids allowed a first major step in the development of tools for archaeal genetics, and to date, several replicative plasmids, expression vectors, and transformation procedures are available for the major groups of Archaea11. Yet, few methods are currently available to generate random mutagenesis and identify genes of interest in uncharacterised biological processes by forward genetic screens. To date, only four archaeal species are amenable to random mutagenesis, and all are mesophilic species. The first two systems were developed for the halophile Haloarcula hispanica12,13 and the methanogen Methanosarcina acetivorans14 by engineering a native insertion sequence or the insect transposon Mariner, respectively. More recently, Methanococcus maripaludis, another methanogen, was mutagenised very efficiently using a Tn5 derivative by transforming the cells with a mixture of the transposon and the transposase15. The last system is based on Mu in vitro transposition followed by the transfer of the mutagenised fragments into the moderate halophile Haloferax volcanii genome by chemical transformation16. Thus, to date, no random mutagenesis approach exists for thermophilic archaea.
In vitro transposition has a great potential for random mutagenesis of the most extremophilic archaea since it does not require a transposon to function in the species to be mutagenised. However, the delivery of DNA in archaea which relies on chemical transformation, electroporation, spheroplast transformation, liposomes or conjugation with Escherichia coli is often very inefficient11. As a consequence, obtaining a large library of mutants may be difficult. In some archaeal species, DNA uptake is greatly facilitated by the ability to spontaneously take up extracellular DNA, allowing integration into the genome and subsequent genetic transformation. This natural transformation occurs at low frequencies in methanogens17,18, acidophiles19 and in the hyperthermophiles Thermococcus kodakarensis20, T. barophilus21 and Pyrococcus furiosus22. In this latter species, a genetically tractable variant named COM1 shows remarkable natural competence with transformation frequencies reaching 103 transformants per μg of DNA22. COM1 is also an uracil auxotroph due to the deletion of the PF1114 gene (ΔpyrF)22,23 which permits the efficient selection of transformants on selective medium without uracil, using the wild-type PF1114 gene as a selection marker. Site-specific integration of the uracil prototrophy marker can be obtained if its sequence is synthetically flanked with the targeted sequence. This strategy has largely been exploited to generate insertion or deletion mutants of P. furiosus in a single step22,23,24. Thus, uracil prototrophy selection, combined with the efficient natural transformation of COM1 allows the possibility to obtain targeted mutants in large numbers. With the random insertion of the uracil protophy marker into genomic DNA, natural transformability of COM1 with this mutagenized DNA could generate libraries of random mutants.
In this study, we describe the development of an approach to generate a random transposon insertion mutant library for Pyrococcus furiosus. This approach combined two steps: first, in vitro transposition mutagenesis of the genomic DNA of P. furiosus using a mariner-based system to generate a library of transposons inserted at random DNA loci; and second, the transfer of the randomly mutated loci to the P. furiosus chromosome by natural transformation. The high natural transformation frequencies allowed us to generate libraries of several thousand mutants of P. furiosus. Interestingly, this procedure has the potential to be adapted to other archaeal species.
Results and Discussion
Parameters affecting transformation and selection of transformants on solid medium
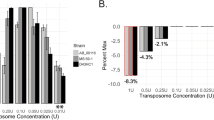
In order to maximise efficiency in construction of a random mutagenesis library for P. furiosus, we first sought to optimise conditions to increase the efficiency of the transformation experiments. Selection of transformants of P. furiosus by natural transformation requires growth on minimal solid medium22. This defined minimal medium is plagued by a relatively low plating efficiency with only 20% of cells forming colonies25. This minimal defined medium contains cellobiose and amino acids as carbon sources, and at elevated temperatures, these can react to form Maillard reaction products that are inhibitory to archaeal growth26. The formation of brown coloured Maillard reaction products is highly dependent on temperature. Indeed, there is a marked change in colour of the growth medium after incubation. The higher the temperature, the darker the colour, and the slower the growth of P. furiosus. Since P. furiosus can grow from 70 to 103 °C27, we have evaluated the plating efficiency at 98 °C compared to 80 °C, a temperature low enough to reduce Maillard reaction products but still sufficient to allow growth of P. furiosus, albeit at a slower rate (doubling time of ~2 h vs ~1 h). We also evaluated three different methods of plating: glass beads, a plastic spreader or direct spotting of liquid cultures on the plates. The plating techniques have limited impact on the plating efficiencies at 98 °C, and range from 15% when plating with a plastic spreader to 25% when plating with glass beads (Fig. 1A). These relatively low plating efficiencies are comparable to previous reports22,25,28. In contrast, we achieved plating efficiencies of up to 82% by incubating plates at 80 °C (Fig. 1A) and spreading with glass beads. For each plating method, we observed a 3 to 4-fold improvement at 80 °C compared to the high temperature incubation. This is further proof that the occurrence of the Maillard reaction, evidenced by the browning of the growth medium, is likely to be responsible for the low plating efficiencies on minimal medium at 98 °C. In subsequent experiments plating was performed using glass beads or spotting, and incubating the plates at 80 °C. We then assessed if natural transformation frequencies could be affected by abiotic or biotic parameters known to impact natural transformation in bacteria, such as pH and growth phase. We found that transformation frequencies varied by more than an order of magnitude with pH, with an optimum at pH 6.2 to 6.5 (Fig. 1B). For some naturally competent bacteria, growth phase can play a large role in natural transformation; however, transformation frequencies of P. furiosus were not dependent on growth phase (Fig. 1C). Thus, the maximum number of transformants of P. furiosus COM1 can be obtained by transforming overnight stationary-phase cultures, selecting on plates at pH 6.5, and incubating at 80 °C. All experiments presented hereafter have been performed using these conditions.

Growth and transformation conditions of P. furiosus COM1.
(A) Plating efficiencies using glass beads, plastic spreader and spotting at two temperatures of incubation (80 °C and 98 °C). Error bars indicate standart deviation from the mean of three independent experiments. (B) Transformation frequencies as a function of pH using flanking regions of 1500 bp. Transformation reactions contained 2 μg DNA per 1 ml of culture. 100 μl or 200 μl of cell culture (108 cells/ml) were respectively mixed with 200 ng or 400 ng of DNA. The transformation frequencies are calculated as the ratio of CFUs on selective medium (without uracil) divided by CFUs on non-selective medium (with uracil) per μg of DNA. Error bars represent standard deviation from the mean of eight independent experiments. (C) Transformation frequencies as a function of growth phase. Growth was monitored every two hours for 24 hours. Cells collected from a culture at time corresponding to exponential growth (4 h), mid-log (8 h), post-exponential (12 h) and stationary phase (24 h) were used for transformation in the same conditions as in panel B. Errors represent standard deviation from the mean of three technical replicates. (D) Transformation frequencies as a function of the size of the homologous flanking regions. Frequencies have been normalised to the copy number of the pyrF gene. Error bars indicate standart deviation from the mean of five independent experiments.
Genomic DNA is the most effective molecule for transformation of P. furiosus
P. furiosus strain COM1 is naturally competent for uptake of genomic, circular and linear DNA22,23. The recombination efficiency in P. furiosus COM1 allows transformation with the pyrF gene with very short homology regions, as small as 40 bp23. Longer homology regions up to 1 kb increased the number of obtained transformants by three orders of magnitude23. To generate a large number of random mutants, it was crucial to determine the type and size of DNA molecule that was most efficient in transformation experiments. This DNA molecule would subsequently become the target of in vitro random mutagenesis. To do so, we compared the efficiencies of the pyrF gene, either carried on PCR products or on genomic DNA (gDNA) to generate transformants by complementing the pyrF deletion in COM1. On PCR products, we varied the size of the homologous regions flanking the pyrF selectable marker starting from 500 bp to 3 kb. The length of these homologous regions are maximal when pyrF is carried on gDNA. As expected from the literature, the number of transformants obtained with PCR products increased with the length of homology, reaching several thousands transformants with only 200 ng of DNA mixed with 100 μl of cell suspension (~107 cells). Interestingly, the best transformation frequencies were obtained with gDNA (Fig. 1D), allowing us to generate several thousand mutants per μg of gDNA. The efficient transformation of P. furiosus cells with gDNA is further indication that DNA is indeed taken up and integrated in the genome by an active natural transformation process. Furthermore, it indicates that gDNA represents the best substrate for in vitro random mutagenesis of this species.
In vitro transposition of genomic DNA and transformation in P. furiosus.
The natural transformability of P. furiosus with genomic DNA offers the possibility to implement an in vitro random transposon mutagenesis strategy as previously described for naturally transformable bacteria29,30. In this strategy genomic DNA is subjected to in vitro transposon mutagenesis with the hyperactive form of the mariner Himar1 transposase (MarC9)31 and a suitable mini-transposon. The mutagenised gDNA is repaired and used to transform the naturally competent species (Fig. 2A). To implement this strategy in P. furiosus we constructed a transposon-containing vector (pNG-Tn-pyrF) with a selectable marker (Pgdh-pyrF) flanked by two inverted tandem repeat sequences (IRL, IRR) specific for the MarC9 Himar1 transposase (Fig. 2A and Supplementary Fig. S1). In order to ensure that transposon insertion effectively disrupts the open-reading frame, stop codons in all possible reading frames were added next to the inverted repeats. The transposon contains a wild type copy of the pyrF gene under the transcriptional control of the glutamate dehydrogenase promoter (gdh; PF1602) with a short terminator sequence (T1) from the hpyA1 gene at the 3′ end of the gene22,23 (Fig. 2A). The hyperactive MarC9 Himar1 transposase recognizes the IRL and IRR sites and transposes intervening DNA by a cut-and-paste mechanism into target DNA at TA sites32. When incubated with the transposon-containing plasmid pNG-Tn-pyrF, the transposase cuts out the transposon, leaving out the plasmid backbone and randomly transposes it into the supplied gDNA (Fig. 2B). The transposition event leaves single-stranded gaps at each transposon-chromosome junction, and these are repaired using T4 DNA polymerase and E. coli DNA ligase (Fig. 2A,B). For the purposes of this application, it was important to use target genomic DNA isolated from a uracil auxotrophic P. furiosus strain that lacked the pyrF marker utilised in the transposon. Furthermore, to enable the use of the resulting library for other downstream applications that might require a second genetic marker, we chose to use P. furiosus strain JFW002 that is also a tryptophan auxotroph, containing a deletion in tryptophan biosynthetic genes (trpAB)23. JFW002 (COM1 ΔpyrF ΔtrpAB) genomic DNA was transposed in vitro using the transposon-containing plasmid pNG-Tn-pyrF and the purified MarC9 Himar1 Mariner transposase (Fig. 2B). Transposed genomic DNA was then used to transform JFW002 cells using uracil prototrophy for selection of the pyrF marker. Compared to genomic DNA which carries pyrF at its original location, the transposon-mutagenised genomic DNA with randomly inserted pyrF gave rise to transformants with only slightly lower transformation frequency (Fig. 2C). A single transformation experiment, using 2 to 4 mL of culture and 2 to 4 μg of transposed DNA, generated from 700 to 2400 transformants (Fig. 2D). Combining four experiments we obtained a library of about 5000 mutants. Thus, P. furiosus can be efficiently and reproducibly transformed with genomic DNA that has undergone in vitro transposon-mediated mutagenesis.

In vitro transposition of mariner-based vector containing the Pgdh-pyrF cassette into genomic DNA using the MarC9 transposase.
(A) General strategy for the creation of a P. furiosus mutant library using in vitro transposition mutagenesis. The transposase MarC9 (dark green circles) recognises the IRL/IRR sites (in red) at the borders of Pgdh-pyrF cassette (carried by the pNG-Tn-pyrF plasmid) and transposes it by a cut-and-paste mechanism into genomic DNA (gDNA). Transposed gDNA is repaired using T4 DNA polymerase and E. coli DNA ligase and used to transform the uracil and trytophan auxotroph JFW02. (B) Agarose gel electrophoresis of the in vitro transposition reaction. Lane 1, transposon donor plasmid pGN-Tn-pyrF alone; lane 2, genomic DNA of strain JFW002 alone; lane 3, transposition reaction containing genomic DNA, donor plasmid pGN-Tn-pyrF and MarC9 transposase. Excision of the transposon from the transposon-donor plasmid pNG-Tn-pyrF can be evidenced by the appearance of a band at the size of the linear plasmid lacking the transposon (N, nicked plasmid; L, linear plasmid; B, backbone plasmid with transposon excised; SC, supercoiled plasmid). (C) Transformation frequencies of P. furiosus JFW002 transformed with in vitro transposed gDNA. Error bars represent standard deviation from the mean of four independent experiments. (D) Example of a transformation reaction with gDNA plated on a selective plate of minimal medium and incubated 5 days at 80 °C.
Natural transformation of P. furiosus with in vitro mutagenised genomic DNA generates random insertion mutants
In order to investigate the nature and dynamics of transposon insertions, we purified 12 mutants and confirmed the presence of the Pgdh-pyrF cassette by PCR amplification. To verify the presence and the copy number of the transposon, these mutants were analysed by Southern hybridisation using a probe specific to the transposon (Fig. 3A). A single predominant fragment was detected in all of isolates, indicating that each harbored a unique transposon insertion. Moreover, each of the 12 tested mutants presented a specific size of hybridisation fragment, suggesting random transposition into the genome of P. furiosus JFW02 (Fig. 3A). The sites of transposon insertion were identified by single primer colony PCR33. We observed that all insertion events occurred at TA sites, and their position are located throughout the genome of P. furiosus JFW02 (Fig. 3B). In all the 12 investigated mutants the transposon interrupted an open reading frame indicating the non-essential nature of the corresponding genes (Table 1 and Supplementary Fig. S2).

Identification and dynamics of transposon insertions in the genome of P. furiosus JFW002.
(A) Southern blot analysis of 12 isolated and purified mutants probed with the pyrF gene. Lanes 1–12: purified randomly selected transposon insertion mutants; pNG-Tn-pyrF: positive control; JFW002: negative control. (B) COM1 genome distribution of the transposon insertions. Gene names (PFxxxx) according to the gene nomenclature for P. furiosus DSM 3638. (C) Typical gene conversion dynamics of transposon insertions monitored by PCR (known insertion). Due to meroploidy, all genomes of the mutant cells may not contain a mutated allele (red arrow, mutated allele; green arrow, wild-type allele). Conversion may require up to three successive subcultures in liquid selective medium. (D) Conversion dynamics of a pool of mutants assessed by plating on minimal medium with or without uracil. Mutants in which gene conversion is incomplete could generate uracil-auxotrophic progeny when plated on non-selective medium (ura+). A ratio of 100% ura+ CFU and ura− CFU indicates that all genomes copies of cells contain the pyrF transposon.
Polyploidy has been suggested as a common trait of all Euryarchaeota34. The chromosome copy number in the Thermococcales family member T. kodakarensis has been determined to range from 9 to 17 copies35. Genome copy number has not been experimentally determined in P. furiosus, but being a closely related species to T. kodakarensis, we anticipated P. furiosus to also be polyploid. Thus, natural transformation of P. furiosus with randomly transposed DNA may initially generate a merodiploidy situation where one copy of a gene is inactivated, while other alleles remain wild-type. During the first few cell generations following transformation with transposed DNA, gene conversion may occur if the selection pressure is maintained, resulting in a single mutated allele in all chromosomal copies. We examined the dynamics of gene conversion following initial isolation of transposition mutants on selective medium. Using PCR analysis we found that 6 out of the 12 tested mutants already displayed only the mutated allele at the initial colony stage. For the remaining mutants two to three passages in selective liquid medium were sufficient to obtain gene conversion (Fig. 3C). Interestingly, mutants in which gene conversion is incomplete could generate uracil-auxotrophic progeny when plated on non-selective medium. We used this phenotype to assess gene conversion of a pool of mutants, which cannot be assessed by the PCR based techniques used for isolated mutants, and found that three passages in liquid selective medium were sufficient to increase the percentage of cells unable to generate uracil-auxotrophic progeny from 60% to nearly 100% (Fig. 3D). This is consistent with conversion rates observed in other archaeal species34,36,37,38.
Conclusions
We report here that the highly efficient natural competence of the hyperthermophilic archaeon P. furiosus can be exploited to easily and rapidly obtain thousands of mutants for the generation of a random mutant library. The number of mutants obtained is dependent on both the plating efficiency and parameters increasing the efficiency of natural transformation, such as incubation temperature and pH. This technique should allow the establishment of an ordered collection of insertional mutants. Combined with a phenotypic screen, efficient random mutagenesis in P. furiosus will allow the identification of novel pathways and specific functions, as has been done for decades in Bacteria and Eucarya. Alternatively, a scale-up of the protocol would allow construction of a saturating insertional mutagenesis library that could be used to define essential and non-essential genes of P. furiosus. Furthermore, the method presented here could be applied to other archaeal species, with only one restriction, e.g. the existence of an efficient transformation protocol. Natural transformation presents several advantages as it does not require chemical or physical alteration of the cells and is efficient with high molecular weight intact gDNA. However, in species that do not efficiently take up gDNA naturally, in vitro mutagenesis could also be performed on fragmented gDNA and then delivered to cells through other methods of transformation.
Materials and Methods
Strains, media and growth conditions
The strains used in this study are listed in Supplementary Table S1. Plasmids used in this study are listed in Supplementary Table S2. The strains were grown anaerobically in a defined medium with cellobiose25 as the carbon source and supplemented with 20 μM uracil for COM1 and JFW002 strains22. Medium was prepared as previously described22. Liquid cultures were inoculated with a 1 to 2% inoculum or with a single colony in 5–50 ml of medium in 50–100 ml serum bottles degassed with three cycles of vacuum and nitrogen, reduced by addition of Na2S (0.1% final concentration), and incubated at 98 °C. Solid medium was prepared by mixing as 50:50 of liquid medium at 2X concentration with 1.6% (wt/vol) Gelrite® (SERVA) solubilised by boiling in water; both solutions were maintained at 95 °C just prior to mixing. The medium was poured into glass petri dishes immediately after mixing. The plates were poured and inoculated aerobically and then put in stainless steel anaerobic jars (Schuett Biotec) with one sachet of Gaspak® EZ Anaerobe (Becton Dickinson and Company, U.S.A). These sachets generate anaerobic conditions in the jar by elimination of O2 in less than 2 hours and production of CO2 in less than 24 hours. The jars were incubated for 4 days at 98 °C or 6 days at 80 °C. To test the effect of pH on the transformation frequency, the defined medium with cellobiose at 2X concentration was prepared by mixing of all components and adjustment to the indicated pH with 1 M NaOH or 1 M HCl. Purification of intermediate strains was performed by plating 10−3 dilutions of transformant cultures onto selective plate medium (without uracil) and picking isolated colonies into selective liquid medium.
Plating efficiency
The density of P. furiosus overnight cultures was determined by counting in a Thoma counting chamber. To estimate the plating efficiency and cell viability, the 10−3, 10−4 and 10−5 dilutions of initial culture were plated using a plastic spreader or glass beads, or by spotting 10 μL of the dilutions. Plating efficiency was calculated as the ratio of the number of colonies observed on the plates to total cell counts determined by counting in a Thoma chamber.
Transformation of P. furiosus COM1
For natural transformation, aliquots of P. furiosus culture, typically grown to mid-log phase (~108 cells/ml, as determined by counting in a Thoma counting chamber) in defined liquid medium at pH 6.5 (except for experiment of Fig. 1B where various pH were tested), were mixed with DNA to obtain a final concentration of 2 μg/ml, as described previously22. Routinely, we used 100 μl or 200 μl of cell culture (108 cells/ml) mixed with 200 ng or 400 ng of DNA, respectively. The mixtures were plated on the selective solid medium without uracil using glass beads. Ten-fold serial dilutions were prepared in 1X base salts22 and spotted on the selective (using 10−1 and 10−2 dilutions) and non-selective plates (using 10−3, 10−4 and 10−5 dilutions) in order to calculate transformation frequencies. The plates were placed inverted in anaerobic jars and incubated at 80 °C for 5–6 days or at 98 °C for 3–4 days. The transformation frequency was calculated as the ratio of the number of CFUs counted on selective medium (without uracil) divided by the number of CFUs counted on non-selective medium (with uracil). DNA constructs used as substrates for natural transformation were produced by PCR amplification of the pyrF gene from wild-type gDNA using different lengths of homology regions ranging from 500 bp to 3000 bp, amplified using the following primer sets: pyrF_500F/pyrF_500R, pyrF_1000F/pyrF_1000R, GL061/GL06222, pyrF_2000F/pyrF_2000R, pyrF_3000F/pyrF_3000R (Supplementary Table S3).
Genomic DNA isolation
Cells from 2 ml of overnight P. furiosus culture were harvested and suspended in 100 μl buffer A (25% sucrose, 50 mM Tris-HCl, 40 mM EDTA, pH 7.4). Next, 250 μl 6 M guanidinium HCl–20 mM Tris, pH 8.5 was added, and the mixtures were incubated at 70 °C for 5 min. Genomic DNA was extracted with phenol-chloroform-isoamyl alcohol (25:24:1; buffered at pH 8), ethanol precipitated, and suspended in 50 μl 10 mM Tris buffer, pH 8.0. Alternatively, genomic DNA was purified from 5 ml of overnight culture using the Promega Wizard® Genomic DNA Purification Kit per the manufacturer’s instructions.
Construction of pNG-Tn-pyrF, plasmid with transposon for in vitro transposition.
The Pgdh-pyrF cassette, containing 283-bp portion of the intergenic region upstream of gdh (PF1602) joined with pyrF gene and T1 terminator from the histone gene hpyA1 (PF1722), was amplified from pJFW07023 plasmid DNA using primers Tn_pyrF_for (containing IRL-sequence and a sequence carrying stop codons in all 6 possible reading frames) and Tn_pyrF_rev (containing IRR-sequence and a sequence carrying stop codons in all 6 possible reading frames). The PCR product was purified and cloned in pJET1.2/blunt vector using CloneJET PCR Cloning Kit following the manufacturer’s instructions (Thermo Scientific, #K1231). The resulting plasmid, named, pNG-Tn-pyrF, was used for in vitro transposition. E. coli DH5α cells were transformed by chemical transformation. Plasmid DNA was isolated from liquid cultures by using QIAprep spin miniprep columns (Qiagen). The plasmid constructs were confirmed by restriction analysis and Sanger sequencing. The full sequence of pNG-Tn-pyrF is available upon request.
MarC9 transposase purification and in vitro transposition of P. furiosus genomic DNA
MarC9 transposase was purified according to a previously published method39. Briefly, an E. coli DH5α λpir strain bearing a plasmid encoding an inducible gene coding for MarC9 (hyperactive Mariner transposase) fused to the Maltose-Binding Protein (MBP) was cultivated in LB, and the overexpression of MBP-MarC9 was induced for 2 hours by addition of IPTG (isopropyl β-D-1-thiogalactopyranoside) during the exponential phase. Cells were lysed by sonication, and the lysate was applied to a column of amylose resin. MBP-MarC9 was eluted by addition of a maltose-containing buffer, aliquoted and stored at −80 °C. A typical purification yielded 100 μg of purified MBP-MarC9 at a concentration of 500 μg/ml. Transposition reactions were also performed as described, with minor modifications31,39,40. The transposition reactions were performed in a buffer containing 10% glycerol, 25 mM HEPES pH 7.9, 250 μg purified and heat-inactivated bovine serum albumin, 2 mM DTT, 100 mM NaCl and 5 mM MgCl2. The optimal concentration of MarC9 for the reaction was determined empirically by performing transposition reactions with 1 μg genomic DNA, 1 μg of pNG-Tn-pyrF plasmid, and a range of two-fold increasing transposase concentrations, followed by 2 hours of incubation at 30 °C. The reaction products were separated by gel electrophoresis and visualised with ethidium bromide staining. Activity of the transposase can be assessed by analysing conversion of the plasmid from the supercoil form into three other forms: nicked (one or both IRs cut on a single strand), linear (double-strand break in a single IR) and shortened linear (both IR cuts and totally excised transposon) (Fig. 2B). Observation of those three forms have been previously described, and longer incubation time did not modify their relative abundances. High concentrations of transposase do not improve the abundance of plasmid backbones with the transposon totally excised, but induce an apparent degradation of gDNA, probably because of a non-specific activity of the transposase on IR-like sites in the chromosome. In contrast to protocols for purification of native MarC9 transposase from inclusion bodies40, no nuclease contaminating activity was observed with purified MBP-MarC9.
Transposition of genomic DNA for natural transformation was performed on 10 μg of plasmid and 10 μg of gDNA. Buffer and transposase volumes were adjusted accordingly. After transposition, the reaction was inactivated by heating at 75 °C for 10 min, then DNA was purified by phenol-chloroforme extraction and ethanol precipitation. The transposition junctions in the DNA were repaired as previously described by filling the junctions with T4 DNA polymerase and nick-repairing with E. coli DNA ligase.
Transformation of P. furiosus with transposon-mutagenised DNA
Transposed and repaired DNA was used for natural transformation in P. furiosus strain JFW002. Overnight culture was grown to ~108 cells/ml (as determined by counting in a Thoma counting chamber) in minimal liquid medium with uracil. 2 to 4 mL of culture were cooled on ice and centrifuged at 4000 rpm for 10 min. The cell pellet was resuspended in 100–200 μl 1X base salts (1/20 of the original culture volume). The cell suspension was mixed with transposed DNA in a proportion of 1 μg DNA for 108 cells. Approximately 2 to 4 μg of DNA were used in each transformation experiment. The mixtures were spread on the selective solid medium without uracil using 100 μl of mixture per plate with glass beads.
Southern-blot analysis of transposition mutants
Approximately 1 μg of gDNA was digested with HindIII restriction enzyme (Thermo Scientific) and resolved on a 0.8% agarose gel for 1 hour at 80 V. Genomic DNA fragments were transferred from the gel to a Hybond-N + membrane (Amersham) using alkaline capillary transfer. The membrane was blocked for 1 hour at 42 °C in ECL Gold Hybridisation Buffer (Amersham) supplemented with 0.5 M of NaCl and 5% (w/v) of single strand salmon sperm DNA. Hybridisation was performed at 42 °C overnight in ECL Gold Hybridisation Buffer, hybridisation probe was labelled by ECL Direct Labelling and Detection System kit (Amersham). Washing, labelling and detection were accomplished according to the manufacturer’s directions using the ECL Direct Labelling and Detection System kit (Amersham). The probe was produced via PCR of plasmid DNA pNG-Tn-pyrF using primers pyrF-1_F and pyrF-2_R (Supplementary Table S3).
Identification of transposition insertion sites
PCR amplification to confirm the presence of transposon insertion was performed using the primers pyrF_1_F and pyrF_2_R (Supplementary Table S3). Single Primer Colony PCR was used to identify the sites of transposon insertion33. PCR was performed as follows: initial denaturation of genomic DNA of tested mutants (98 °C, 5 min), 20 high stringency cycles (98 °C, 10 sec; 50 °C, 30 sec; 72 °C, 90 sec), 30 low stringency cycles (98 °C, 10 sec; 30 °C, 30 sec; 72 °C, 90 sec) and 30 high stringency cycles (98 °C, 10 sec; 50 °C, 30 sec; 72 °C, 90 sec) followed by a final extension at 72 °C for 7 min. Primer pyrF_P1 (Supplementary Table S3) is designed in order to hybridise 118 base pairs upstream of the inverted tandem repeat (ITR). A second round of PCR is then performed using a ten-fold dilution and the above PCR reaction with primer pyrF_P2 (Supplementary Table S3) which hybridises 20 pairs downstream of pyrF_P1. This PCR reaction was send out for Sanger sequencing (GATC-biotech, Germany) using primer pyrF_seq.
Additional Information
How to cite this article: Guschinskaya, N. et al. Random mutagenesis of the hyperthermophilic archaeon Pyrococcus furiosus using in vitro mariner transposition and natural transformation. Sci. Rep. 6, 36711; doi: 10.1038/srep36711 (2016).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Raymann, K., Brochier-Armanet, C. & Gribaldo, S. The two-domain tree of life is linked to a new root for the Archaea. Proc. Natl. Acad. Sci. USA. 112, 6670–6675 (2015).
Spang, A. et al. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521, 173–179 (2015).
Hilpert, R., Winter, J., Hammes, W. & Kandler, O. The sensitivity of archaebacteria to antibiotics. Zentralblatt Für Bakteriol. Mikrobiol. Hyg. Abt Orig. C Allg. Angew. Ökol. Mikrobiol. 2, 11–20 (1981).
Wiley: Archaea: Molecular and Cellular Biology - Ricardo Cavicchioli. Available at: http://eu.wiley.com/WileyCDA/WileyTitle/productCd-1555813917.html. (Accessed: 27th September 2016).
Littlechild, J. A. Archaeal Enzymes and Applications in Industrial Biocatalysts. Archaea 2015, e147671 (2015).
Schiraldi, C., Giuliano, M. & De Rosa, M. Perspectives on biotechnological applications of archaea. Archaea 1, 75–86 (2002).
Vannier, P., Marteinsson, V. T., Fridjonsson, O. H., Oger, P. & Jebbar, M. Complete genome sequence of the hyperthermophilic, piezophilic, heterotrophic, and carboxydotrophic archaeon Thermococcus barophilus MP. J. Bacteriol. 193, 1481–1482 (2011).
Oger, P. et al. Complete Genome Sequence of the Hyperthermophilic Archaeon Thermococcus sp. Strain AM4, Capable of Organotrophic Growth and Growth at the Expense of Hydrogenogenic or Sulfidogenic Oxidation of Carbon Monoxide. J. Bacteriol. 193, 7019–7020 (2011).
Zeng, X., Jebbar, M. & Shao, Z. Complete Genome Sequence of Hyperthermophilic Piezophilic Archaeon Palaeococcus pacificus DY20341T, Isolated from Deep-Sea Hydrothermal Sediments. Genome Announc. 3, (2015).
Allers, T. & Mevarech, M. Archaeal genetics — the third way. Nat. Rev. Genet. 6, 58–73 (2005).
Farkas, J. A., Picking, J. W. & Santangelo, T. J. Genetic techniques for the archaea. Annu. Rev. Genet. 47, 539–561 (2013).
Dyall-Smith, M. L. & Doolittle, W. F. Construction of composite transposons for halophilic Archaea. Can. J. Microbiol. 40, 922–929 (1994).
Woods, W. G., Ngui, K. & Dyall-Smith, M. L. An improved transposon for the halophilic archaeon Haloarcula hispanica. J. Bacteriol. 181, 7140–7142 (1999).
Zhang, J. K., Pritchett, M. A., Lampe, D. J., Robertson, H. M. & Metcalf, W. W. In vivo transposon mutagenesis of the methanogenic archaeon Methanosarcina acetivorans C2A using a modified version of the insect mariner-family transposable element Himar1. Proc. Natl. Acad. Sci. USA. 97, 9665–9670 (2000).
Sarmiento, F., Mrázek, J. & Whitman, W. B. Genome-scale analysis of gene function in the hydrogenotrophic methanogenic archaeon Methanococcus maripaludis. Proc. Natl. Acad. Sci. USA. 110, 4726–4731 (2013).
Kiljunen, S. et al. Generation of comprehensive transposon insertion mutant library for the model archaeon, Haloferax volcanii, and its use for gene discovery. BMC Biol. 12, 103 (2014).
Bertani, G. & Baresi, L. Genetic transformation in the methanogen Methanococcus voltae PS. J. Bacteriol. 169, 2730–2738 (1987).
Worrell, V. E., Nagle, D. P. Jr., McCarthy, D. & Eisenbraun, A. Genetic transformation system in the archaebacterium Methanobacterium thermoautotrophicum Marburg. J. Bacteriol. 170, 653–656 (1988).
Berkner, S. & Lipps, G. Genetic tools for Sulfolobus spp.: vectors and first applications. Arch. Microbiol. 190, 217–230 (2008).
Sato, T., Fukui, T., Atomi, H. & Imanaka, T. Targeted gene disruption by homologous recombination in the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1. J. Bacteriol. 185, 210–220 (2003).
Thiel, A., Michoud, G., Moalic, Y., Flament, D. & Jebbar, M. Genetic manipulations of the hyperthermophilic piezophilic archaeon Thermococcus barophilus. Appl. Environ. Microbiol. 80, 2299–2306 (2014).
Lipscomb, G. L. et al. Natural competence in the hyperthermophilic archaeon Pyrococcus furiosus facilitates genetic manipulation: construction of markerless deletions of genes encoding the two cytoplasmic hydrogenases. Appl. Environ. Microbiol. 77, 2232–2238 (2011).
Farkas, J. et al. Recombinogenic properties of Pyrococcus furiosus strain COM1 enable rapid selection of targeted mutants. Appl. Environ. Microbiol. 78, 4669–4676 (2012).
Lewis, D. L. et al. A mutant (‘lab strain’) of the hyperthermophilic archaeon Pyrococcus furiosus, lacking flagella, has unusual growth physiology. Extrem. Life Extreme Cond. 19, 269–281 (2015).
Raven, N. D. & Sharp, R. J. Development of defined and minimal media for the growth of the hyperthermophilic archaeon Pyrococcus furiosus Vc1. FEMS Microbiol. Lett. 146, 135–141 (1997).
Kim, K. W. & Lee, S. B. Inhibitory effect of Maillard reaction products on growth of the aerobic marine hyperthermophilic archaeon Aeropyrum pernix. Appl. Environ. Microbiol. 69, 4325–4328 (2003).
Fiala, G. & Stetter, K. O. Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100 °C. Arch. Microbiol. 145, 56–61 (1986).
Farkas, J., Chung, D., DeBarry, M., Adams, M. W. W. & Westpheling, J. Defining Components of the Chromosomal Origin of Replication of the Hyperthermophilic Archaeon Pyrococcus furiosus Needed for Construction of a Stable Replicating Shuttle Vector. Appl. Environ. Microbiol. 77, 6343–6349 (2011).
Akerley, B. J. et al. Systematic identification of essential genes by in vitro mariner mutagenesis. Proc. Natl. Acad. Sci. 95, 8927–8932 (1998).
Pelicic, V., Morelle, S., Lampe, D. & Nassif, X. Mutagenesis of Neisseria meningitidis by in vitro transposition of Himar1 mariner. J. Bacteriol. 182, 5391–5398 (2000).
Lampe, D. J., Akerley, B. J., Rubin, E. J., Mekalanos, J. J. & Robertson, H. M. Hyperactive transposase mutants of the Himar1 mariner transposon. Proc. Natl. Acad. Sci. 96, 11428–11433 (1999).
Akerley, B. J. & Lampe, D. J. Analysis of gene function in bacterial pathogens by GAMBIT. Methods Enzymol. 358, 100–108 (2002).
Martinez, E., Cantet, F. & Bonazzi, M. Generation and Multi-phenotypic High-content Screening of Coxiella burnetii Transposon Mutants. J. Vis. Exp. JoVE doi: 10.3791/52851 (2015).
Hildenbrand, C., Stock, T., Lange, C., Rother, M. & Soppa, J. Genome Copy Numbers and Gene Conversion in Methanogenic Archaea. J. Bacteriol. 193, 734–743 (2011).
Spaans, S. K., van der Oost, J. & Kengen, S. W. M. The chromosome copy number of the hyperthermophilic archaeon Thermococcus kodakarensis KOD1. Extremophiles 19, 741–750 (2015).
Soppa, J. Polyploidy in archaea and bacteria: about desiccation resistance, giant cell size, long-term survival, enforcement by a eukaryotic host and additional aspects. J. Mol. Microbiol. Biotechnol. 24, 409–419 (2014).
Soppa, J. Ploidy and gene conversion in Archaea. Biochem. Soc. Trans. 39, 150–154 (2011).
Zerulla, K. & Soppa, J. Polyploidy in haloarchaea: advantages for growth and survival. Front. Microbiol. 5, 274 (2014).
van Opijnen, T. & Lazinski, D. W. & Camilli, A. Genome-Wide Fitness and Genetic Interactions Determined by Tn-seq, a High-Throughput Massively Parallel Sequencing Method for Microorganisms. Curr. Protoc. Mol. Biol. Ed. Frederick M. Ausubel Al 106, 7.16.1–7.16.24 (2014).
Prudhomme, M., Camilli, A. & Claverys, J. P. In The Molecular Biology of Streptococci 511–517 ( Hakenbeck, R. & Chhatwal, G. S., Norwich, UK: Horizon Scientific Press, 2007).
Acknowledgements
This work was supported in part by the Agence Nationale de la Recherche (ANR-10-BLAN-1752-01 Living deep) and in part by the CNRS ExoMod program to PO. This work was supported by a CNRS-Inserm ATIP-Avenir grant awarded to XC and by the U.S. National Science Foundation (CBET-1264052) to MWA.
Author information
Authors and Affiliations
Contributions
N.G., R.B., M.T., P.O. and X.C. designed, performed and analysed experiments. G.L.L. and M.W.W.A. provided reagents, expertise and feedback. N.G., P.O. and X.C. wrote the manuscript. N.G. and X.C. prepared the figures. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Guschinskaya, N., Brunel, R., Tourte, M. et al. Random mutagenesis of the hyperthermophilic archaeon Pyrococcus furiosus using in vitro mariner transposition and natural transformation. Sci Rep 6, 36711 (2016). https://doi.org/10.1038/srep36711
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep36711
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
