
Abstract
Tumors exhibit complex organization and contain a variety of cell populations. The realization that the regenerative properties of a tumor may be largely confined to a cell subpopulation (cancer stem cell) is driving a new era of anti-cancer research. Cancer stem cells from Glioblastoma Multiforme tumors express markers that are also expressed in non-cancerous neural stem cells, including nestin and Sox2. We previously showed that the transcription factor Hes3 is a marker of neural stem cells and that its expression is inhibited by JAK activity. Here we show that Hes3 is also expressed in cultures from glioblastoma multiforme which express neural stem cell markers, can differentiate into neurons and glia and can recapitulate the tumor of origin when transplanted into immunocompromised mice. Similar to observations in neural stem cells, JAK inhibits Hes3 expression. Hes3 RNA interference reduces the number of cultured glioblastoma cells suggesting a novel therapeutic strategy.
Similar content being viewed by others


Introduction
Glioblastoma Multiforme (GBM) is the most common primary brain tumor and is associated with very poor prognosis1,2. Tumor initiating cells (Cancer Stem Cells, “CSCs”) have been identified in GBM3,4,5,6,7 and a race is on to understand how to target them as a potential therapeutic avenue. Although the lineage connection between non-cancerous neural stem cells (NSCs) and CSCs is not established, these cell types share several properties including signaling pathways that contribute to their growth as well as the expression of common markers including nestin and Sox28.
We recently demonstrated that the transcription factor Hes39,10,11 is a marker of established neural stem cell cultures from the fetal and adult rodent central nervous system12,13,14,15,16. Factors that promote the expansion of these cells, including Notch ligands, Angiopoietin 2, insulin, as well as inhibitors of the Janus kinases (JAK) and p38MAP kinases, also increase the expression of Hes3. In the adult rodent and primate brains, Hes3 identifies a perivascular cell subpopulation that co-expresses other markers of immature cells, such as Sox2. In this report, we demonstrate that HES3 not only provides a novel identifying marker for the CSC population in GBMs, but is a key mediator of the number of these cells, thereby offering a potential therapeutic target.
Results
In biopsies from patients with GBM, Hes3 co-localizes with the putative cancer stem cell marker prominin14,17,18 (Fig. 1a,b). These results agree with the observations that magnetic immunoprecipitation of cells from the rodent brain using an antibody against prominin generate a cell fraction which is enriched in Hes3+/Sox2+ cells14. Taken together, these data suggest that Hes3 may be expressed in putative cancer stem cells in GBM and that it may mediate cell number expansion.

Hes3 is expressed in putative cancer stem cells in glioblastoma.
(a) Hes3+ cells are located adjacent to blood vessels in biopsies from GBM patients. (A blood vessel lumen is marked in white; green arrows point to Hes3+ cells; the red arrow points to nuclei of vascular cells). (b) Another example of Hes3+ cells in a GBM tumor show co-localization of Hes3 with prominin (red and green split channels are shown side-by side). White circles show the location of the nuclei, some of which appear positive for Hes3 signal (Marked by green arrows). (c) Cultured GBM cells co-express Sox2 and Ang2. (d-f) GBM cells in the presence of EGF and the absence of serum (“Self-Renewal”) express low amounts of GM1+ gangliosides, identified by labeling with a fluorophore-conjugated cholera toxin B subunit; addition of serum for 4 days increases labeling. (g) Diagram showing positive regulation on Hes3 transcription by bFGF via phosphorylation of STAT3-serine and opposition by JAK activity. Hes3 is positively regulated by signals that promote STAT3-serine phosphorylation in the absence of STAT3-tyrosine phosphorylation. In NSCs, such signals include bFGF, insulin, the Notch ligands Delta4 and Jagged1 and Angiopoietin 2. Signals that induce the JAK/STAT pathway (which leads to STAT3-tyrosine phosphorylation) do not induce Hes3 transcription. Such signals include CNTF, EGF and components of serum. [Nuclei are stained with DAPI and appear blue. Size bar: 100 micrometers].
To address the potential value of Hes3 as a target in cancer medicine, we used primary cultured cells that were isolated from GBM biopsies. These cells were maintained in vitro using standard protocols for the establishment of non-cancerous neural stem cells cultures19,20,21. These cells can be expanded with the support of the mitogen Epidermal Growth Factor (EGF) or basic Fibroblast Growth Factor (bFGF)21. Our previous work has established that the same cells used in this study can be propagated in culture over long periods of time and many passages, express several markers of NSCs, including, Sox2 and nestin, as well as prominin, can be induced to differentiate into neurons, astrocytes and oligodendrocytes, they phenocopy the tumor of origin in xenograph experiments and they respond to many growth factors that NSCs also respond to21,22.
We recently showed that cultured NSCs express Angiopoietin 2 (Ang2) as well as its receptor, Tie2 and that treatment with Ang2 increases cell number both in vitro and in vivo14,15. Here we show that GBM cells also express Ang2 together with the more established marker Sox2 (Fig. 1c). NSCs express a small number of GM1+ gangliosides on their cell surface which can be identified by binding to a fluorophor-conjugated B subunit of cholera toxin16; in contrast, their differentiated progeny expresses many binding sites. In our cultured GBM cells (in the presence EGF or bFGF), a very small percentage of cells labeled with conjugated cholera toxin (Fig. 1d–f). In contrast, when cells where treated with serum (which induces the differentiation of NSCs), the number of cholera toxin binding sites greatly increased. These results further support that the cultured GBM cells used express markers commonly associated with neural stem cells and that they possess the potential to differentiate into cells representing the main cell types of the nervous system.
Our previous work with NSCs showed that Hes3 expression is opposed by the actions of JAK. Its activity can be assessed by measuring STAT3 phosphorylation on tyrosine residue 705 (STAT3-Tyr), which is downstream of JAK23. In contrast, Hes3 expression is supported in conditions where STAT3 is phosphorylated on serine residue 727 (STAT3-Ser)12. NSCs cultured in bFGF exhibit low Hes3 mRNA levels, but these can be increased by concomitant exposure to Ciliary Neurotrophic Factor (CNTF) and JAK inhibitor (JAK I)12, a treatment which elevates STAT3-Ser and inhibits STAT3-Tyr (Suppl. Fig. 1a,b). Similar results can be obtained by low concentrations of CNTF24 that also elevate STAT3-Ser but not STAT3-Tyr phosphorylation (Suppl. Fig. 1c). The potential of JAK I to promote the undifferentiated state in NSC cultures is also demonstrated by the fact that it inhibits the expression of the glial marker GFAP in the presence of bFGF (Suppl. Fig. 1d). Low CNTF concentrations or concomitant high concentrations of CNTF and JAK I increase the number of cultured NSCs in the presence of bFGF (Suppl. Fig. 1e).
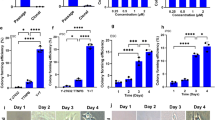
We hypothesized that GBM cells cultured in the absence of serum and the presence of mitogen would induce the expression of Hes3 and, therefore, they would resemble, in that respect, the cells of their origin in the patient. We tested the following four conditions: (a) EGF, (b) EGF + JAK I, (c) bFGF and (d) bFGF + JAK I. We found that the highest STAT3-Ser and lowest STAT3-Tyr phosphorylation state was in the FGF + JAK I condition (Fig. 2a). In these conditions, cells were able to proliferate; in fact, their number was greater than in the other conditions, after one week of treatment (Fig. 2b). We note that inclusion of the JAK inhibitor allowed cell cultures to reach a higher terminal cell density, as assessed by cell number counting following a 2 week treatment (Fig. 2c,d). Hes3 expression varied among the four conditions; the highest incidence of Hes3+ cells was in the bFGF + JAK I condition (Fig. 2e,f). Since the culture system presented here lacks serum to support cell growth, the presence of a mitogen (bFGF or EGF) is required. As a consequence, we studied the effects of treatments (e.g. JAK I) in the presence of these defined factors.

Hes3 regulates the number of GBM cells in culture.
(a) Culture conditions affect the phosphorylation states of STAT3 of GBM cells. (b) GBM cells grow efficiently in the four conditions described in the text (7-d). (c) JAK inhibition increases the terminal cell density that GBM cells reach (14-d). (d) JAK inhibition increases the terminal cell density that GBM cells reach (14-d; representative phase contrast images). (e) Culture conditions regulate the probability that GBM cells will express Hes3 (7-d). (f) Examples of Hes3 staining in human GBM cells, in the FGF+JAK I conditions (7-d; size bar: 25 micrometers. Nuclei are stained with DAPI and appear blue). (g) Hes3 knockdown in GBM cells (line “X08”) reduces the amount of Hes3 protein and cell number (5-d). Each control bar represents a different control (“scrambled”) siRNA product and each of the three “Hes3” labeled bars represents a different Hes3 siRNA product for corroboration. (h–j) Similar results are obtained using two additional GBM cell cultures (“X01” and “X04”) in the presence of EGF. (k) A cell viability assay (Promega MTS) shows reduced viability following Hes3 interference (5-days).
To address whether Hes3 is a mediator of the expansion of GBM cells and, therefore, a putative anti-cancer target, we performed transfections with Hes3 siRNA. We tested two different control (“scrambled”) siRNA products and three different Hes3 siRNA products for corroboration. We performed the experiments in both EGF and bFGF + JAK I conditions to show the sturdiness of the result and to demonstrate that the effect of Hes3 interference is not an artifact of a particular cell culture system. In all cases, Hes3 interference caused the reduction of Hes3 expression, as expected and a reduction in cell number (Fig. 2g). We confirmed the results using two additional GBM cell cultures established from different patients (Fig. 2h–j). A cell survival assay showed a reduction in viability in the Hes3 siRNA – transfected cells, suggesting that, partly, the reduction in cell number is due to a reduction in viability (Fig. 2k).
Discussion
Brain tumors contain multiple cell types, some of which may carry the lion’s share of the regenerative properties of the tumor, manifested as metastasis and recurrence. Different subpopulations of cells express different sets of markers (with plenty of overlap) allowing for the ability to distinguish among these cell types to a certain degree. These cells may also differ in terms of the signal transduction pathways they utilize in order to grow. Given the variations in extracellular signals within the tumor microenvironment, this is a logical postulate. It is, therefore, critical to study and define different signaling systems that contribute to the growth of tumors and design manipulations to counteract tumor growth.
In this manuscript we applied findings that we generated during our previous work with non-cancerous NSCs12,13,14,15,16. This work showed that NSCs utilize non-canonical signal transduction pathways to regulate their survival and numbers in vitro and in vivo. Key to these non-canonical pathways is the phosphorylation state of STAT3. When STAT3 is phosphorylated on serine 727 in the absence of detectable phosphorylation on tyrosine 705, cells survive and proliferate in a self-renewing manner. In contrast, when the tyrosine residue is also phosphorylated, NSCs differentiate with a preference towards glial fates19,25,26. STAT3-Ser phosphorylation promotes the expression of the transcription factor Hes3, which we showed to be a novel biomarker of established NSCs. In addition, Hes3 identifies new pools of cells in various areas of the adult brain which can be placed in culture where they exhibit self-renewal and multi-potentiality, generating neurons, astrocytes and oligodendrocytes.
We also showed that Hes3 is expressed in tumor tissue from glioblastoma patients and that it co-localizes with the putative CSC marker prominin. Here, we extended our studies to show that (1) primary GBM cell cultures also express Hes3 under conditions that support STAT3-Ser phosphorylation and suppress STAT3-Tyr phosphorylation and that (2) Hes3 interference reduces the numbers of cells in culture.
These early results suggest that Hes3 is a new marker of GBM cells and a mediator of cell expansion. Future studies will address the relevance of Hes3 in cultures from different GBM subtypes (e.g., as defined by The Cancer Genome Atlas). It will also be important to address the mechanisms that regulate the cell number changes reported here. For example, our results help distinguish between cell expansion rates and maximal cell density. Specifically, whereas inclusion of the JAK I in the presence of EGF did not alter cell number during the early culture phase, it did allow cells to reach higher densities. Given that JAK is implicated in the canonical Notch signaling pathway and subsequent gliogenic differentiation27, it is intriguing to speculate that non-canonical Notch signaling branches which are opposed by JAK12 may be competing with canonical pathways and that Hes3 may be a critical regulator of the balance between these two branches.
Our previous work with non-cancerous NSCs showed that activation of the STAT3-Ser/Hes3 signaling pathway promotes cell survival without altering the cell cycle duration12. Whether this applies to GBM cells, will have to be determined in future studies.
It is difficult to define a cancer stem cell population. The GBM-derived cells used in this work, however, do exhibit CSC properties, as suggested by their (1) expression of undifferentiated markers, (2) ability to differentiate into neuron and glia - like cells and (3) ability to phenocopy the tumor of origin in immunocompromised mice. These results suggest that Hes3 may be relevant as a target for the elusive CSC population of GBM and, possibly other solid tumors such as breast, colon and pancreatic cancers28,29,30,31,32. Preliminary results show that in spontaneously arising brain tumors in adult mouse null for the p53 tumor suppressor gene, Hes3+ cells are clearly seen both within the tumor mass (identified by morphology and staining for KI67), as well as outside the borders of the tumor (data not shown). Additional work will require validation in multiple in vitro and in vivo paradigms.
Beyond the possible value of Hes3 as a CSC target, these results show that the same cell (or cell line) is capable of utilizing distinct signaling pathways to ensure its growth. Glioma cells are often cultured in the presence of serum, or in the absence of serum with EGF. Here we show that the same GBM cell cultures can be cultured in EGF or FGF + JAK I in the absence of serum. EGF and serum components activate the JAK-STAT pathway which suppresses Hes3 expression, whereas bFGF activates the STAT3-Ser pathway which induces Hes3 expression. Therefore, the same cell can grow under conditions that not only are starkly different, but even opposing. This complexity at the level of signal transduction of a single cell confers growth options to the cell and may be a major contributor to the difficulties in combating cancer.
Methods
Neural stem cell culture
Neural Stem Cell cultures from the mid-gestation (E13.5) mouse embryo cortex were grown as previously described14. Tissue was dissected under a dissection microscope, triturated in 1 ml N2 medium containing bFGF with a 1 ml pipette until no tissue clamps were seen; the triturate was allowed to settle for 1 min and the top 0.9 ml was collected, diluted in N2 containing bFGF and plated. Cells were expanded in serum-free DMEM/F12 medium with N2 supplement and bFGF (20 ng/ml) for 5 days under 5% oxygen conditions and were re-plated fresh or from frozen stocks at 1,000–10,000 cells per cm2. bFGF was included throughout our experiments, unless otherwise stated. To induce differentiation, the culture medium was replaced with fresh culture medium which does not contain bFGF and the cells were incubated for 5 days. Treatments were as described in the main text. In the experiments involving the treatment of cells with CNTF and the JAK inhibitor, the inhibitor was added to the culture medium 30-min prior to the CNTF.
Glioblastoma cell culture
Three GBM cell lines (“X01”, “X04” and “X08“ lines) were established from acutely resected human tumor tissues (Glioblastoma multiforme). Resected tissue was treated using the protocol described above for mouse fetal neural stem cells. Cells were cultured under the same conditions that were used for rodent neural stem cells. Variations in their mitogenic support are noted in the text. Treatments were as described in the main text.
Pharmacological Treatments
bFGF and EGF were purchased from RnD Systems and were used at a concentration of 20 ng/ml. JAK inhibitor was purchased from Calbiochem and was used at a concentration of 200 nM.
Human biopsies of solid tissue
All human tissues in this study were obtained during surgical resections from patients with newly diagnosed or recurrent tumors. Materials in excess of pathological evaluation were used for research purposes in accordance with protocols approved by the Institutional Review Board of the National Institutes of Health. Written consent was obtained and all research tumor tissues were de-identified.
Animal handling
Animals were handled and housed according to the German Federal guidelines for the use and care of laboratory animals and the study was approved by the Landesdirektion Dresden. For research performed at the National Institutes of Health (USA), all research involving animals was conducted in accordance with NINDS ACUC (National Institute of Neurological Disorders and Stroke/Animal Care and Use Committee) guidelines and after their approval.
RNA interference
We used the Amaxa nucleofector equipment to electroporate siRNA into GBM cell cultures. Two control siRNA products were used (Santa Cruz Biotechnology, sc37007 and OriGene Tech, SR3004) and three anti-Hes3 siRNA products (Santa Cruz Biotechnology, sc88003and OriGene Tech, SR318208A and SR318208C). The day before transfection, 5 million cells were dissociated from a culture flask and transferred to T25 Falcon culture flask with suspension medium containing EGF or bFGF. Cells were then transfected with siRNA using a nucleofecting electroporator according to the manufacturer’s protocol (Amaxa Inc., Gaithersburg, MD, USA). After 24 h, the medium was replaced and cells were harvested for additional experiments. Typical transfection efficiency was at approximately 80%21,22.
Reverse transcriptase PCR
RNA was extracted from cell cultures with Trizol (Invitrogen, 15596-026) and PCR reactions were performed with the ProSTAR First-Strand RT-PCR Kit (Stratagene, 200420).
Immunohistochemistry
Under deep anesthesia, animals were perfused transcardially with a rinse of saline, followed by 4% formaldehyde fixative (pH 7.4). Brains were removed immediately, stored in the fixative solution overnight and then in 30% sucrose for 3 days. Brains were frozen-sectioned at 16 or 30 micrometers.
Immunocytochemistry
Cells were fixed onto the culture plate by replacing the culture medium with ice cold 4% formaldehyde fixative (pH 7.4), for 30 min. PFA was washed twice with PBS and the cells were immunostained using standard techniques.
Western Blot analysis
Western Blot analyses were performed using standard protocols. Briefly, Laemmli Sample Buffer from Bio-Rad was used as the harvesting buffer. Samples were boiled for 15 min and approximately 40 microgram of protein was loaded per well. Primary antibodies were used at a dilution of 1:500 and secondary antibodies at 1:4,000. Antibodies were diluted in 5% BSA in PBST. Primary antibodies were incubated overnight, with mild shaking, at 4 degrees Celsius.
Cell viability assays
Cell viability was measured by the MTS assay (Promega).
Reagents
We used the following reagents and antibodies: bFGF (233-FB), fibronectin (1030-FN), from R&D; JAK Inhibitor I (420099), from Calbiochem; Polyornithine (P-3655), insulin (I9278) from Sigma; Alexa-Fluor-conjugated secondary antibodies from Invitrogen; DAPI (D-8417) from Sigma, ECL reagents (34080) from Pierce, polyacrylamide gradient gels from Invitrogen, HRP-conjugated secondary antibodies from Jackson Immunoresearch and general chemicals from Sigma. For immunohistochemical staining and Western Blotting, we used antibodies against the following markers: Hes3 (25393), STAT3 (482), pSer727-STAT3 (8001-R), pTyr705-STAT3 (7993) from Santa Cruz; α-tubulin from Sigma, (T-6074). Hes3 siRNA was from Santa Cruz Biotechnology and OriGene Tech (please see Electroporation section).
Statistical analysis
Results shown are the mean ± S.D. Asterisks identify experimental groups that were significantly different (p-value<0.05) from control groups by the Student's t-test (Microsoft Excel), where applicable.
References
Deorah, S., Lynch, C. F., Sibenaller, Z. A. & Ryken, T. C. Trends in brain cancer incidence and survival in the United States: Surveillance, Epidemiology and End Results Program, 1973 to 2001. Neurosurg Focus 20, E1 (2006).
Stupp, R. et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol 10, 459–66 (2009).
Ignatova, T. N. et al. Human cortical glial tumors contain neural stem-like cells expressing astroglial and neuronal markers in vitro. Glia 39, 193–206 (2002).
Singh, S. K. et al. Identification of a cancer stem cell in human brain tumors. Cancer Res 63, 5821–8 (2003).
Singh, S. K. et al. Identification of human brain tumour initiating cells. Nature 432, 396–401 (2004).
Galli, R. et al. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res 64, 7011–21 (2004).
Hemmati, H. D. et al. Cancerous stem cells can arise from pediatric brain tumors. Proc Natl Acad Sci U S A 100, 15178–83 (2003).
Li, Z., Wang, H., Eyler, C. E., Hjelmeland, A. B. & Rich, J. N. Turning cancer stem cells inside out: an exploration of glioma stem cell signaling pathways. J Biol Chem 284, 16705–9 (2009).
Hirata, H., Tomita, K., Bessho, Y. & Kageyama, R. Hes1 and Hes3 regulate maintenance of the isthmic organizer and development of the mid/hindbrain. Embo J 20, 4454–66 (2001).
Lobe, C. G. Expression of the helix-loop-helix factor, Hes3, during embryo development suggests a role in early midbrain-hindbrain patterning. Mech Dev 62, 227–37 (1997).
Hatakeyama, J. et al. Hes genes regulate size, shape and histogenesis of the nervous system by control of the timing of neural stem cell differentiation. Development 131, 5539–50 (2004).
Androutsellis-Theotokis, A. et al. Notch signalling regulates stem cell numbers in vitro and in vivo. Nature 442, 823–6 (2006).
Androutsellis-Theotokis, A., Rueger, M. A., Mkhikian, H., Korb, E. & McKay, R. D. Signaling pathways controlling neural stem cells slow progressive brain disease. Cold Spring Harb Symp Quant Biol 73, 403–10 (2008).
Androutsellis-Theotokis, A. et al. Angiogenic factors stimulate growth of adult neural stem cells. PLoS One 5, e9414 (2010).
Androutsellis-Theotokis, A. et al. Targeting neural precursors in the adult brain rescues injured dopamine neurons. Proc Natl Acad Sci U S A 106, 13570–5 (2009).
Androutsellis-Theotokis, A., Walbridge, S., Park, D. M., Lonser, R. R. & McKay, R. D. Cholera toxin regulates a signaling pathway critical for the expansion of neural stem cell cultures from the fetal and adult rodent brains. PLoS One 5, e10841 (2010).
Miraglia, S. et al. A novel five-transmembrane hematopoietic stem cell antigen: isolation, characterization and molecular cloning. Blood 90, 5013–21 (1997).
Weigmann, A., Corbeil, D., Hellwig, A. & Huttner, W. B. Prominin, a novel microvilli-specific polytopic membrane protein of the apical surface of epithelial cells, is targeted to plasmalemmal protrusions of non-epithelial cells. Proc Natl Acad Sci U S A 94, 12425–30 (1997).
Johe, K. K., Hazel, T. G., Muller, T., Dugich-Djordjevic, M. M. & McKay, R. D. Single factors direct the differentiation of stem cells from the fetal and adult central nervous system. Genes Dev 10, 3129–40 (1996).
Androutsellis-Theotokis, A. et al. Generating neurons from stem cells. Methods Mol Biol 438, 31–8 (2008).
Soeda, A. et al. Hypoxia promotes expansion of the CD133-positive glioma stem cells through activation of HIF-1alpha. Oncogene 28, 3949–59 (2009).
Soeda, A. et al. Epidermal growth factor plays a crucial role in mitogenic regulation of human brain tumor stem cells. J Biol Chem 283, 10958–66 (2008).
Levy, D. E. & Darnell, J. E., Jr Stats: transcriptional control and biological impact. Nat Rev Mol Cell Biol 3, 651–62 (2002).
Ravin, R. et al. Potency and fate specification in CNS stem cell populations in vitro. Cell Stem Cell 3, 670–80 (2008).
Bonni, A. et al. Regulation of gliogenesis in the central nervous system by the JAK-STAT signaling pathway. Science 278, 477–83 (1997).
Rajan, P. & McKay, R. D. Multiple routes to astrocytic differentiation in the CNS. J Neurosci 18, 3620–9 (1998).
Kamakura, S. et al. Hes binding to STAT3 mediates crosstalk between Notch and JAK-STAT signalling. Nat Cell Biol 6, 547–54 (2004).
Al-Hajj, M., Wicha, M. S., Benito-Hernandez, A., Morrison, S. J. & Clarke, M. F. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A 100, 3983–8 (2003).
Dalerba, P. et al. Phenotypic characterization of human colorectal cancer stem cells. Proc Natl Acad Sci U S A 104, 10158–63 (2007).
O'Brien, C. A., Pollett, A., Gallinger, S. & Dick, J. E. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature 445, 106–10 (2007).
Ricci-Vitiani, L. et al. Identification and expansion of human colon-cancer-initiating cells. Nature 445, 111–5 (2007).
Li, C. et al. Identification of pancreatic cancer stem cells. Cancer Res 67, 1030–7 (2007).
Acknowledgements
The work was supported by the Department of Medicine, University of Dresden and by a grant from the Center for Regenerative Therapies Dresden, Dresden to AA-T.
Author information
Authors and Affiliations
Contributions
A.A.T. conceived the project, A.A.T., D.M.P. and S. Poser designed experiments, A.A.T., D.M.P., J.J., J.M., S.M., D.E., S.S., R.R., N.P., S. Pacioni and S. Poser performed experiments, A.A.T., D.M.P., J.J., R.D.M. and S. Poser analyzed data. A.A.T., D.M.P. and S. Poser wrote the main manuscript text. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors have submitted two patents relating to this work.
Electronic supplementary material
Supplementary Information
Supplemental Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Park, D., Jung, J., Masjkur, J. et al. Hes3 regulates cell number in cultures from glioblastoma multiforme with stem cell characteristics. Sci Rep 3, 1095 (2013). https://doi.org/10.1038/srep01095
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep01095
This article is cited by
-
Cell biology of glioblastoma multiforme: from basic science to diagnosis and treatment
Medical Oncology (2018)
-
Streptozotocin-induced β-cell damage, high fat diet, and metformin administration regulate Hes3 expression in the adult mouse brain
Scientific Reports (2018)
-
Enlightening discriminative network functional modules behind Principal Component Analysis separation in differential-omic science studies
Scientific Reports (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
