
Abstract
The functional complexity of the central nervous system (CNS) is unparalleled in living organisms. Its nested cells, circuits and networks encode memories, move bodies and generate experiences. Neural tissues can be engineered to assemble model systems that recapitulate essential features of the CNS and to investigate neurodevelopment, delineate pathophysiology, improve regeneration and accelerate drug discovery. In this Review, we discuss essential structure–function relationships of the CNS and examine materials and design considerations, including composition, scale, complexity and maturation, of cell biology-based and engineering-based CNS models. We highlight region-specific CNS models that can emulate functions of the cerebral cortex, hippocampus, spinal cord, neural-X interfaces and other regions, and investigate a range of applications for CNS models, including fundamental and clinical research. We conclude with an outlook to future possibilities of CNS models, highlighting the engineering challenges that remain to be overcome.
Key points
-
Isolating the mechanisms of central nervous system (CNS) functions will benefit human health and well-being, decrease economic burdens and inspire the design of neuromorphic computers and artificial intelligences.
-
Customizable, bioengineered tissues can recapitulate CNS structures and functions, representing simplified platforms that allow the systematic assessment of neural development and pathology in vitro.
-
CNS regions, such as the cerebral cortex, hippocampus, brainstem and spinal cord, can be modelled with organoids, spheroids, microfluidic chips and bioprinted or scaffold-based constructs that combine cells and materials.
-
Increasingly biomimetic CNS models have the potential to display higher-order functions, including cognitive abilities and conscious experience.
Similar content being viewed by others
Introduction
A functional understanding of the central nervous system (CNS) lies at the core of some of the biggest unanswered questions in science, such as the nature of consciousness1 or the boundaries of life and death2, because brains encode the unique patterns of thought, feeling and behaviour that define the individual. The socio-economic impact of our incomplete knowledge of CNS function is best illustrated by the rising annual worldwide costs associated with Alzheimer disease, which are estimated to have exceeded $1 trillion per year in 2020 and are expected to reach US$9 trillion per year by 2050 (ref. 3). Nevertheless, known principles of CNS function are inspiring the design of artificial intelligences, which have become increasingly enmeshed with day-to-day human life4. However, new techniques must be developed to achieve a comprehensive understanding of the CNS.
Animals were long used as models of human anatomy, physiology and behaviour5, and have contributed to key discoveries in CNS biology, including the characterization of the action potential6. Their ubiquity in biomedical research is a testament to their predictive validity7; however, to improve physiological relevance and translational outcomes, and to increase tractability, human-based CNS models are required. In addition, animal housing and handling costs, facility requirements, low sample sizes and ethical concerns have, in many cases, made in vitro models more appealing to understand CNS function. Although CNS tissues of animals closely reflect those of humans, they are inflexible experimental tools; that is, animal brains are constrained by morphogenetic factors that determine a highly restricted range of phenotypes. We have gained a wealth of knowledge by probing conserved nuclei and tract systems in animal models, but many fundamental questions require greater degrees of control and are, therefore, impractical to answer in vivo.
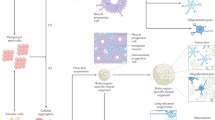
Innovations in stem cell biology and neural tissue engineering have made the bioengineering of customizable, 3D in vitro CNS models possible, enabling the study of human neural development and disease (Fig. 1). Soon after the development of the first techniques to visualize microscopic brain anatomy8, basic colloidal hydrogels were developed9 and CNS tissues were cultured for the first time with frog cells using ‘the hanging drop’ technique10, followed by chick and rodent cell culture in flasks11. The identification of multipotent stem cells12 and their presence in the brains of postnatal rats12 then enabled the design and control of bioengineered tissues using hydrogels based on cross-linked polymeric networks that would serve as tissue scaffolds and cell-infused inks13. Developments in microelectrode array (MEA) technology14,15, bioprinters16 and microfluidic devices17,18 allowed the construction and functional assessment of bioengineered CNS models with high precision. Today, neural cells can be created by reprogramming induced pluripotent stem (iPS) cells from fibroblasts19,20, enabling the creation of cerebral organoids21 with region-specific identities22 and complex functions23. Similar functional capacities can be achieved in scaffold-based 3D neural tissues24,25. In addition, the blood–brain barrier (BBB) can be modelled in microfluidic devices18 and bioprinted constructs26 (Fig. 1). Such functional in vitro models of the human CNS must recapitulate some, but not all, features of the CNS. Bioengineered tissues typically lack some native elements of CNS tissues and, thus, may not capture recondite interactions. However, they should be sufficiently biologically representative to address a specific research question (Box 1).

3D bioengineered neural tissues are becoming increasingly inexpensive, reproducible, scalable and susceptible to high-throughput investigations27. They also benefit from higher complexity compared with monolayer cultures, recapitulating neural microenvironments without the intrinsic damage associated with the preparation of organotypic slice cultures28. Importantly, bioengineered CNS models can be precisely configured, in which cell type and density, extracellular matrix (ECM) composition, orientation, geometry, mechanical properties and interfacial properties can be customized to test specific hypotheses or to enable transplantation.
Structure and function of the CNS
Modelling the CNS requires an understanding of its basic structure–function relationships at multiple scales (Fig. 2). At the macroscale, the CNS is a collection of heterogeneous tissues encased within the bones of the skull and vertebral column. Their coordinated activities enable sensation, perception, memory and voluntary movement. Each neural region’s specialized function is determined by its tissue cytoarchitecture, which is defined by cell-type composition, neural microenvironment and local circuit structures, as well as the configuration of efferent and afferent connections that link it with other regions within and outside the CNS.

Bioengineered tissue models of the central nervous system (CNS) are designed and built to mimic the structures and functions of in vivo tissues. At the macroscale, the CNS can be subdivided into gross regions, including the cerebrum, brainstem and spinal cord. The cerebrum can be further partitioned into several mesoscale sub-organs, such as the cerebral cortex, basal ganglia and hippocampus — all of which are encased within a dense field of nerve fibres (that is, white matter). At the microscale, cells form highly ordered circuits, interfaces and pathways that reflect complex functions. Cortical and hippocampal circuits display re-entrant or looped activation patterns, which enable cell synchronization and the integration of information associated with perception and memory. However, when left unchecked, re-entrant loops can precipitate seizure activity. Similarly, the neurovascular unit forms a blood–brain barrier (BBB) by tightly joining endothelial cells, pericytes, glial cells and neurons in concentric layers — a selectively permeable gateway that becomes compromised with neurodegenerative disease, cancers, infections and trauma. At finer scales, cells are immersed within complex microenvironments patterned with biophysical and biochemical cues that determine their fate and behaviour. To accurately model CNS function in vitro, the essential elements need to be identified that optimally recapitulate structures and functions of the CNS. CA, cornu ammonis.
The cerebrum comprises several mesoscale tissues, including the cerebral cortex, the hippocampus and several basal nuclei, or ganglia, which provide executive control29, memory encoding30 and conscious experience31. Deep within the centre of the cerebrum are the thalamic and hypothalamic nuclei, which are collections of cells that transmit signals from the brainstem to the cortex32 and regulate autonomic functions33, respectively. Anatomically continuous with the thalamus are the divisions of the brainstem: the midbrain, pons and medulla. Each subdivision contains nuclei with distinct sensory, motor and vital functions. Functionally coupled to the brainstem is the cerebellum that coordinates the timing and execution of fine movements as well as features of cognition34. The spinal cord, which is continuous with the brainstem and runs down the centre of the vertebral column, consists of a core of cell bodies surrounded by nerve fibres that receive sensory data from and transmit output motor sequences to the peripheral nervous system35. The CNS also contains specialized olfactory and retinal tissues, which transduce chemical and optical signals, respectively.
At the microscale, the CNS is composed of several distinct cell populations. Neurons are electrically excitable cells that are specialized for rapid cell-to-cell communication. They synthesize neurotransmitters and neuropeptides which serve as information carriers within complex networks. Oligodendrocytes interact with neurons to form an insulating myelin sheath along neuronal axons, increasing signalling rates tenfold. Ependymal cells secrete cerebrospinal fluid to clear waste, maintain brain buoyancy and mitigate mechanical impacts. In addition, astrocytes buffer extracellular molecules, and microglia perform immune functions. These and other cells of the CNS exist cooperatively within a complex microenvironment composed of a proteoglycan-rich ECM36 that is further specialized at neuronal surfaces by perineuronal nets37, facilitating the stabilization of connections between cells within neural networks. CNS cells, such as pericytes and astroglia, also form specialized vascular interfaces that define the BBB38. If cells of the CNS are damaged or if their ability to release, transport or sequester specific signals is compromised, they can precipitate psychological states of depression, anxiety and psychosis39.
Design considerations of CNS models
Bioengineered CNS models can capture only key features of native CNS tissues, and therefore several design elements should be considered to maximize their validity as experimental tools (Fig. 3).

Design elements can be selected and combined to recapitulate desired physiological states in bioengineered central nervous system (CNS) tissues. The composition, dimensions, maturity, complexity and interfaces can be customized by selecting different design elements. ECM, extracellular matrix.
Composition
Perhaps the most essential design consideration involves the selection of relevant cell types for co-culture. In addition to the many subtypes of glia and other non-neural cells, molecular analyses have revealed various neuronal subtypes in the human brain40,41; however, the true number likely ranges from several hundred to a thousand42. Primary tissue extracts from animals can provide balanced ratios of excitatory and inhibitory neuronal populations and native glial cell concentrations; however, the presence or absence of any particular cell type in primary tissue isolates is dependent upon neurodevelopment43. Therefore, cells must be harvested from embryonic, perinatal or adult animals, depending on the purpose, balancing cell viability and plasticity with region-specific phenotypes.
The goal of CNS modelling is often the investigation of human physiology and disease, and therefore iPS cells, embryonic stem (ES) cells or neural stem cells (NSCs) from human sources should be included. These can be differentiated along specific neural lineages, generating programmed neuronal populations with specific neurotransmitter and neuropeptide profiles44 as well as astrocytes, microglia and oligodendrocytes45. Patient-specific cells can also be harvested and transformed to CNS lineages to generate personalized human CNS models with the individual’s genetic material to optimally simulate their unique disease phenotype and overcome treatment resistance with high-throughput drug screening46. Cells and species can be combined in vitro, including human–chimpanzee47,48 and other hybrid systems. Indeed, evolutionary hypotheses can be addressed by inserting the genes of extinct hominids, such as Neanderthals, into human brain organoids49.
In addition to cell-type composition, materials must be carefully selected to create scaffolds (for example, hydrogels), bioinks as well as other bulk components and surfaces to simulate distinct CNS microenvironments. Importantly, the elastic modulus of adult brain ECM (0.1–1.0 kPa) differs greatly from embryonic brain ECM (110 Pa) or spinal cord ECM (90–230 kPa)50. Biophysical cues modulate cell migration, differentiation and neurotransmission, and therefore ECM properties, such as stiffness, should be considered in the CNS model51. Mammalian polymers, such as collagen or hyaluronan, can be purified from decellularized ECM extracts to generate physiologically relevant hydrogels and scaffolds52. Alternatively, synthetic polymers provide flexible, biocompatible options with highly tunable properties53.
Dimensions
Bioengineered CNS tissues can be designed sufficiently modular to support multiscale patterning, as observed in native tissues. For example, organoids can be fused to create ‘assembloids’ with circuit-like or system-like properties54, including neuromuscular junctions55. Neurospheroids, formed by microscale cell aggregations, can be daisy-chained into functional arrays, or shaped by polydimethylsiloxane (PDMS) moulds into mesoscale tissue blocks, which can, in turn, be combined to form macroscale networks of neural modules56. Bioprinting supports customizable tissue geometry with single-cell precision57, and laminar organizations characteristic of the cortex58 or neurovascular unit26. Scaffolds offer optimal control over compartmentalization of tissue types; for example, toroidal geometries provide high surface areas and centre surround-type organizations that reflect the nested nuclei of CNS regions, such as the amygdala, hippocampus and hypothalamus. In addition, microfluidic conduits or etched surfaces can be engineered to guide neurites or vasculature, and to polarize tissue structure59.
Maturity
Cell maturity is an important element in CNS modelling, because plasticity, regenerative potential, genetic expression, metabolic activity and disease processes are dependent upon development. Models derived from human stem cells and non-human embryonic primary tissues rich in progenitors may be ideal tools to investigate neurodevelopmental disorders (for example, autism, schizophrenia)60; however, such models may not be representative of adult phenotypes, which are less plastic, more specialized and have less regenerative potential compared with embryonic tissue-based models. The ageing methylome — the total distribution of cytosine-bound methyl groups in the genome — and its epigenetic interactions with genotypic differences of sex and ethnicity should also be considered61. Cells with high stemness may extend cell viability for long-term culture, but are less relevant as models of CNS injury or degeneration62. Tissue maturity can also be tuned independent of cell sources by changing the mechanical properties of scaffolds; here, material stiffness can mimic ECM alterations associated with normal ageing63 and age-related disease64. For example, the expression of a truncated form of lamin A protein (progerin) induces ageing and disease phenotypes in a genetically susceptible iPS cell-based model of Parkinson disease65. In addition, pH66, electrical conductivity67, viscoelasticity68 and physiological concentrations of iron and other metals69 may be customized to achieve phenotypes that reflect different states of brain maturity.
Complexity
The CNS is the most functionally complex organ in the human body. Complexity describes the interaction of a set of elements to generate non-linear, synergistic or emergent functions that are greater than the sum of their parts. As elements are removed from a system to evaluate their roles, fewer interactions are possible and complexity may decrease. Therefore, modelling the CNS should strike a balance between functional relevance and utility (Box 1). Bioengineered models of the CNS are often deliberately designed to be less complex than their natural templates because simple models are more tractable than in vivo equivalents, offering greater control over each element and the number of potential interactions (for example, neural, endothelial, glial, ECM)70. From an experimental perspective, fewer possible interactions increase the likelihood of identifying causal factors that contribute to development, regeneration, injury or disease. However, the elimination of key elements can contribute to the suppression of emergent or synergistic functions and, therefore, to spurious conclusions. Thus, complexity must be considered at scale, depending whether the research question may be better addressed by low-complexity models that simulate simple circuits71 or high-complexity alternatives that simulate networks and systems72,73.
Interfaces
The brain and spinal cord interface with multiple tissues to orchestrate bodily functions. Cranial nerves and ascending tract systems from the spinal cord relay sensory data to the brain, informing cognitive functions. Other cranial nerves and descending tracts conduct information from the brain to lower-level effectors that drive muscle activation to enable ambulation and speech. Without sensory inputs and motor outputs, functional models of the CNS are closed-loop systems that are intrinsically resistant to assessment of higher-order functions. Similarly, the integrity of the CNS is dependent upon the perfusion of nutrients and oxygen across neurovascular interfaces at single-cell resolution. Moreover, systems-level inputs from the cerebral ventricles, the gut and other viscera are important.
Modelling the CNS
Modelling approaches
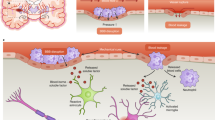
Functional CNS models can be designed by engineering-based or cell biology-based approaches (Fig. 4). Cell biology-based models, which include organoids, spheroids and assembloids, are generated by cell-autonomous processes, resulting in the self-organization of tissues into complex architectures consistent with the development of organisms28. Despite their unparalleled recapitulation of native CNS structure–function, cell biology-based models lack the intrinsic tunability of engineering-based models that combine cells with materials to direct tissue patterning. Here, tissue organization can be engineered by bottom-up approaches, that is, organizing modular tissues into complex structures, or top-down approaches, in which cells are seeded within pre-assembled 3D microenvironments that provide cues for self-organization74.

Engineering-based models combine materials and cell sources to generate biomimetic central nervous system (CNS) tissues. Synthetic materials (for example, polycaprolactone (PCL), poly(3,4-ethylenedixoythiophene) (PEDOT)), biomaterials (such as chitosan, collagen, silk) or combinations thereof are used to generate highly customizable and tractable hydrogels, 3D bioprintable inks, scaffolds and organ-on-a-chip systems. However, engineered models require judicious selection and micromanagement of design elements to avoid generating physiologically aberrant tissues. Cell biology-based models offer greater cytoarchitectonic complexity and ideal 3D microenvironments at the cost of some customizability. Self-assembling organoids, assembloids and spheroids, derived from pluripotent stem cells (PSCs) and primary tissues, do not require support materials, and display conserved neurodevelopmental programmes consistent with in vivo tissues.
Organoids
Neural organoids are formed by cellular self-organization with inputs from soluble factors and other microenvironmental cues. To generate organoids, embryoid bodies composed of aggregated pluripotent stem cells (PSCs) are typically embedded in Matrigel (solubilized native matrix rich in ECM components, such as laminin, collagen and embedded factors), in which cell sorting and tissue patterning progress towards a 3D morphology that is 0.5–4.0 mm wide with embryonic CNS features, ideally suited for studies of neurodevelopment and evolution75. Non-directed protocols, which lack factors to guide tissue patterning, yield whole-brain phenotypes termed cerebral organoids, which are highly variable in cell-type composition and spatial organization28. Dual SMAD inhibition can induce neural ectodermal fate commitment22. Alternatively, in directed protocols, additional morphogens are introduced to generate region-specific neural organoids22 by guiding tissue growth and patterning.
Organoids have been developed with cortical, hippocampal, diencephalic, mesencephalic, ventricular and cerebellar phenotypes22; however, tissue organization and cell type ratios remain incomplete. In addition, the integration of 3D vasculature to support stable, long-term viability of organoids remains limited, often suffering from poor perfusion of nutrients and oxygen, which restricts organoid volume and can result in necrotic cores76. Moreover, organoids are not particularly modular structures and, similar to organisms, are limited by conserved developmental programmes.
Spheroids
Neurospheroids consist of aggregated cells from primary tissues, immortalized cell lines or stem cells. In the absence of adherent surfaces, or when forced together by centrifugation or gravity, cells adhere to each other77, and therefore spheroid generation does not require scaffolding or other external support. Spheroids establish their own 3D microenvironments by secreting ECM78; however, their internal structures are less reflective of in vivo cytoarchitecture compared with organoids. Although spheroids are typically less than 1 mm in diameter, their cell densities (105–106 cells mm−3) are similar to in vivo CNS tissues56. Therefore, despite their uniformity and simplicity, spheroids represent excellent models of neurodegenerative disease, brain tumours and other neuropathologies. To increase their scale and complexity, spheroids can be fused to form cuboid, mesoscale neural building blocks with cell-specific identities56. These, in turn, can be connected to generate macroscopic ensembles that display functional network properties. However, without microfluidic-assisted perfusion or other supports, spheroids lack vascularization, preventing continuous, long-term culture79 and limiting their utility for studies of maturation, senescence or long-term exposure to drugs and environmental hazards.
3D-bioprinted models
Bioprinted CNS models combine materials and cells to form 3D tissue constructs with single-cell spatial resolution. In direct bioprinting, cells are infused with hydrogels to create printable bioinks that can be extruded as continuous filaments, ejected as discrete droplets or sculpted by ultra-precise, laser-based printing techniques to form complex 3D tissue architectures26. Printable materials include alginate, collagen, chitosan, gelatin, poly(ethylene glycol) diacrylate (PEGDA) and silk fibroin, which can be partially cross-linked or doped with catalysts prior to printing for rapid gelation80. Printing needles can be designed to enable simultaneous deposition of cells, gels and cross-linkers using co-axial nozzle configurations81, thus allowing high spatial resolution and fabrication of hollow tube structures. Cells from primary tissues, immortalized lines, stem cell donors or combinations thereof can be incorporated in bioinks to customize the properties of the model system, including viability, plasticity or regenerative competence. However, 3D bioprinting is currently limited by material constraints (such as viscosity), shear forces, lower cell densities relative to native tissues and cell-perturbing gelation methods involving chemical and light-based catalysts.
Microfluidic models
Microfluidics-based CNS-on-a-chip models distil the most essential features of a physiological process from multivariate neural systems. Although less complex than other models, microfluidic-based CNS models allow the incorporation of biophysical and chemical signalling gradients to mimic microenvironmental cues, interstitial fluid flow and single-cell resolution neurite guidance82. Microfluidic PDMS platforms can be fabricated by soft lithography, using a template to emboss and print identical replicate chips for maximum reproducibility and high-throughput modelling83. These chips are optically clear, and thus CNS-on-a-chip models enable real-time monitoring of cell migration and network formation84. Moreover, specialized tissue–tissue interfaces can be designed in multichannel chips to model the BBB17 and the gut–brain axis85. Although versatile tools for drug discovery, microfluidic-based models are not ideal implantables, which may be addressed by integrating materials with high biocompatibility, biodegradability and wireless controllers86. In addition, they often lack 3D cytoarchitecture, they contain only few cell types and PDMS can absorb drugs and proteins87.
Materials
Engineering-based 3D modelling techniques combine cells with suitable materials to pattern tissues into reproducible 3D constructs. To guide their initial assembly and support long-term function, materials for CNS modelling52 should reflect the physical, chemical and mechanical properties of native tissues and their unique 3D microenvironments. In addition to providing structural stability, materials can facilitate oxygen and nutrient transport, waste outflow and ECM deposition. Materials can be modified by altering local surface topology or, in bulk, by integrating cell signalling peptides to increase biocompatibility.
Matrix scaffolds and hydrogels
The in vivo conditions of the CNS can be closely mimicked by decellularized CNS tissues, including the meninges. These ECM scaffolds can be re-seeded with different cell populations88, serving as signal-rich microenvironments for neural precursor cells and promoting viability, adherence and differentiation. Hydrogels composed of collagen, hyaluronic acid, silk fibroin, alginate or chitosan28,36 also closely approximate natural ECM scaffolds36, and their cross-linked polymeric network structures can be infused with biological factors to improve cell growth and proliferation, or to guide tissue organization. Alternatively, synthetic polymers, such as polyvinylidene fluoride, have piezoelectric properties that can guide cellular signalling and regeneration89. Similarly, poly(3,4-ethylenedixoythiophene) (PEDOT) layers can be integrated with hydrogels to enhance electrical conductivity90. Depending on the material, hydrogels can be made biocompatible, enabling cellular grafting to repair injured brain tissues following trauma or stroke91. Cell-free scaffolds, channels and hydrogels may also be fabricated prior to cell seeding by indirect bioprinting, extending the selection of materials and techniques to mimic 3D neural microenvironments26. However, such scaffolds suffer from batch-to-batch variability, size and density restrictions, and scaling issues, which can limit high-throughput applications92.
Biomaterials
Cells of the CNS are embedded within and connected by a specialized ECM that maintains tissue stability, regulates diffusion of molecules, controls local biomechanical signalling, participates in synaptogenesis, guides neurite outgrowth, supports cell migration and inhibits tissue remodelling. The main components of brain ECM include fibrous glycoproteins, such as collagen (type IV), laminin, fibronectin and tenascins, as well as several proteoglycans and hyaluronic acid36. Therefore, biomaterials extracted and purified from decellularized ECM, such as Matrigel, are suitable options for CNS modelling93. Matrigel represents a common substrate for CNS bioengineering; however, its contents are variable between batches, incompletely defined and sourced from cancerous tissues with oncogenic ECM microenvironments. Nevertheless, natural biopolymers are biocompatible and promote cell–material interactions, such as adhesion, growth, differentiation and network formation; however, rapid degradation, variable mechanical properties and cytotoxic degradation products may limit their use in CNS models.
Collagen is abundant in all connective tissues of the body, including in the CNS. Collagen type IV is most common in the brain; however, collagen type I, which can be inexpensively and readily derived from rat tail extracts, has tunable cross-linking properties that can be exploited to tailor mechanical strength, porosity and other physiologically relevant factors93. To promote cell–material interactions, collagen is often combined with other synthetic and natural polymers, such as hyaluronan. Although hyaluronan does not favour cell adhesion, combined with collagen in hydrogels, its high porosity and viscoelastic properties enhance survival and CNS regeneration, and promote neurite outgrowth, proliferation of neural precursors and differentiation94. Collagen and hyaluronan can be extracted from the ECM of vertebrates, showing low antigenicity with minimal inflammatory response when used as grafting materials, and, similar to gelatin, agarose or alginate, represent common bioink building blocks for 3D-printed CNS models.
Chitosan is the deacetylated form of chitin, one of the most abundant natural polysaccharides. Chitin has traditionally been sourced from crustacean shells, but is also available from beetles, algae, bacteria and yeast95. Chitosan can be formed into sponges, gels, bioinks and scaffolds. In hydrogel form, chitosan supports cell adhesion, survival, neurite outgrowth and CNS regeneration, with anti-fungal and anti-bacterial properties96. Similarly, silk fibroin — derived from the cocoons of Bombyx mori — is a biocompatible, highly tunable biomaterial that can be formed into hydrogels with a stiffness similar to that of CNS tissues (<100 kPa)50 and a controllable rate of biodegradation. Porous scaffolds composed of silk fibroin microfibres can be combined with collagen type I to support CNS models with synchronous network activity and compartmentalization of cell bodies and neurites25, as well as long-term in vitro structure–function stability to study chronic neurodegenerative disease97. Silk can also be combined with electrically conductive materials to integrate microelectronics98.
Synthetic materials
Synthetic materials address many limitations of natural biomaterials, offering high mechanical stability and control over scaffold topology, fibre alignment and biodegradability53. Although typically less biocompatible, synthetic materials can be coated with biomaterials or infused with neurotrophic factors to enhance cell–scaffold interactions. In addition to glass, ceramics and metals, synthetic polymers such as poly(ethylene glycol) (PEG), polycaprolactone (PCL), poly-l-lactic acid (PLLA) and PEDOT represent non-biologically derived options for neural tissue engineering99. Of note, the hydrophilicity of synthetic biopolymers can be tuned100 to avoid monocyte adhesion to hydrophobic material surfaces, and subsequent immune rejection of grafts.
PEG is non-immunogenic and offers tunable mechanical properties that mimic soft tissues, such as the brain101. For example, PEGDA hydrogels promote neurite extension to regenerate CNS tissues102. PCL is an elastic, slow-degrading synthetic polymer that is typically generated by electrospinning103. Combined with natural polymers, it is biocompatible and promotes cell adhesion; however, PCL can have cytotoxic effects if combined with organic solvents, limiting some applications, including experiments involving the dilution of water-insoluble drugs, which are common in neuropsychiatric research104. PLLA scaffolds made of nanoscale and microscale fibres mimic the ECM, displaying a high surface-to-volume ratio as well as high porosity and variable pore sizes, promoting cell migration, differentiation and neurite outgrowth105. PLLA can be biofunctionalized with natural biomaterials, which form covalent bonds with the ester linkages along its polymer backbone.
Synthetic polymers can not only mimic the neural microenvironment but also provide additional functions, such as electroconductivity. For example, PEDOT can be used to electrically record and stimulate bioengineered neural tissues, promoting differentiation of stem cells and guiding axonal outgrowth by galvanotaxis93. Combining PEDOT with polystyrene sulfonate (PSS) or biomaterials, such as chitosan and silk, allows the creation of biofunctionalized, electroconductive 3D printable hydrogels and scaffolds for multiple applications, including integrated MEAs for real-time electrical monitoring106. Carbon nanotubes, which can also be integrated with 3D printing, have similar electrical properties to PEDOT, guiding neurite outgrowth in brain and spinal cord regeneration93.
PDMS is a low-cost silicon elastomer amenable to high-throughput manufacturing. PDMS can serve as a mould to shape bioprinted and neurospheroidal assemblies, enabling fine control of features. In addition, PDMS is commonly used as a substrate for microfluidic CNS model systems83. However, the hydrophobic properties of unmodified PDMS can be problematic for tissue engineering owing to issues of cell adhesion, which can be addressed by collagen coating or topographical patterning107.
Region-specific CNS models
Whole-brain CNS modelling was initially achieved in 2013 with an organoid system21, and since then innovations in 3D microenvironment biomimicry have allowed the engineering of increasingly vascularized108 and functionally competent23,109 CNS tissues. However, to isolate and interrogate the diverse functions of neural tissues, region-specific model systems are being explored, circuits and pathways reconstructed, multiple techniques synergistically integrated and multiplexed functional read-outs developed (Table 1).
Cerebrum
Cerebral cortex
The cerebral cortex is a thin (1–4 mm), multilayered tissue (3–6 layers) that envelops the superficial folds of the cerebrum. It is divided into at least 50 subregions with distinct functional correlates, from vision and hearing to moral judgement110. Many functional models have been explored for the cerebral cortex, in part because it is the primary neural correlate of conscious experience111 but also because it is particularly vulnerable to degeneration from ageing and disease. However, to increase physiological relevance and enable clinical translation, the most essential elements of cortical tissue structure–function relationships need to be captured in bioengineered models.
Cortical organoid models have long suffered from persistent embryonic-like phenotypes and poor vascularization112. Postnatal phenotypes can be achieved after approximately 9 months in culture, closely mirroring in vivo development and confirming the conservation of intrinsic neurodevelopmental programs in vitro113. Similar periods of maturation were observed in silk scaffold-based cortical models based on neurons and glia derived from iPS cells114, which, similar to organoids, can be cultured continuously for 2 years or longer97. Mature, long-term cultures are key to investigating mechanisms of neurodegenerative diseases and other protracted or chronic illnesses.
Microfluidic devices provide platforms for neurovascular interfacing18 and enable vascularization in cortical organoids. Alternatively, vascularization can be achieved through co-culture with human umbilical vein endothelial cells115, or by transplanting organoids into immunodeficient rodents112. Transplanted into the brains of animals, cortical organoids116 and scaffold-based constructs117 form graft-to-host projections and integrate in native tissues. For example, an organoid model overexpressing the early endothelial progenitor marker ETV2 transcription factor can form perfused vascular-like structures with tight junctions, characteristic of the BBB118. Owing to their tunable porosity, hydrogel-based and scaffold-based cortical models can balance important 3D microenvironmental features such as the perfusion of oxygen and nutrients to facilitate nervous system repair119. Furthermore, neurovascular interfacing can be achieved using cortico-vascular spheroid hybrids120 or 3D-bioprinted neurovascular tissues with laminated cytoarchitecture58 and tunable BBB-relevant cell subtypes such as pericytes and vascular smooth muscle cells26.
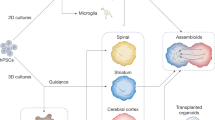
The functional competence of cerebral cortex models can be assessed by analysing electrical dynamics. For example, electric oscillations in cortical organoids driven by mature glutamatergic and GABAergic (GABA (γ-aminobutyric acid)-producing) neurons resemble human electroencephalography in preterm neonates23. These cortical organoids contain high glial cell concentrations and show an age-dependent decrease in progenitors. Electric responses consistent with learning can also be evoked by patterned stimulation of a silk fibroin scaffold-based cortical model embedded in a collagen hydrogel121. However, the functional potential of bioengineered CNS models is often underestimated and mainly based on electrophysiological techniques that were initially designed to assess 2D cultures. With the emergence of 3D MEAs14 and other highly integrated monitoring platforms, it is now possible to assess electrical, optical, chemical and thermal parameters simultaneously. These techniques are being applied to assess the functional correlates of assembloids, which can recapitulate features of the human brain connectome including cortico-thalamic circuits54. Indeed, CNS models may soon be sufficiently complex to express higher-order functions, including cognition in vitro122. For example, the term ‘consciousnessoid’ implies that neural correlates of consciousness may be detectable in vitro123.
Cortical modelling has long been limited by the absence of gyrification — the process by which the cerebrum acquires its folded morphology. To address this limitation, 4D bioprinting can be applied to combine cells with ‘smart’ materials that can be activated by near-infrared light or other triggers to recreate temporal changes in the cerebral structure consistent with neurodevelopment in vivo124. Similarly, microfluidic chips can be integrated with neurospheroids and organoids to assess the impact of cerebral fluid flow on 3D cortical folding dynamics in vitro125. Cortical folding in developing organisms is regulated by the stiffness of the overlaying skull and other mechanical factors126, and therefore gyrification may be tuned by constraining tissue expansion with materials.
Hippocampus
The hippocampal bodies, which comprise interlocking c-shaped tissue structures, are located deep within the temporal lobes. Their cells receive direct inputs from the neighbouring entorhinal cortex, where pace-making stellate cells deliver a steady input of theta (~7 Hz) oscillations that couple with and phase-modulate gamma (~40 Hz) oscillations. Together, these signal patterns are crucial for memory encoding and retrieval127. Hippocampal bodies degenerate in patients with Alzheimer disease and other dementias, leading to impaired theta–gamma coupling and memory deficits128. Bilateral resection of hippocampal bodies results in dense anterograde amnesia129.
Few models of the hippocampus have been bioengineered thus far, despite its importance as a major correlate of dementias and neurodegenerative pathologies, such as Alzheimer disease. Biomaterial-based hydrogels130,131,132, bioprinted constructs133, multichannel scaffolds134 and microfluidic devices135 seeded with primary hippocampal cultures allow the investigation of axon guidance, ECM-dependent viability and pathologies such as traumatic brain injury. Nanofabricated graphene oxide-functionalized scaffolds promote hippocampal cell differentiation136, and microfluidic devices enable the creation of cortico-hippocampal circuits-on-a-chip137.
Hippocampal organoids were first generated by adapting cerebral organoid protocols138. These self-organized tissues contain pyramidal neurons and granule cell types that define the cornu ammonis (CA) fields and dentate gyrus, which are essential contributors to the hippocampal circuit — a complex structure which has yet to be fully recapitulated in vitro owing to inadequate controls over cell composition and tissue polarization. Interestingly, hippocampal organoids also display interfacial features characteristic of the choroid plexuses, which generate cerebral spinal fluid within the cerebral ventricles. Such hippocampal organoids have been used to assess the neuroinvasive potential of SARS-CoV-2, as a model of CNS-related COVID-19 infection139. In addition, iPS cell-derived hippocampal spheroids can serve as tools for patient-tailored assessment and treatment of Alzheimer disease140.
Basal ganglia
The basal ganglia include the putamen, caudate nuclei, nucleus accumbens and other regions that are functionally tied to involuntary movement, decision-making, learning and addiction141. Dysfunctions of neurons that transmit dopamine, glutamate and GABA within and between the basal nuclei underlie the signs and symptoms of Parkinson disease and other disorders142.
The putamen and caudate nucleus, which together form the striatum, represent the most commonly modelled regions of the basal ganglia. Human striatal organoids express medium spiny neurons with GABAergic markers and postnatal electrical phenotypes143. They also contain glutamatergic neurons, astrocytes and oligodendrocytes. Fused with cortical organoids, these striatal organoids form cortico-striatal assembloids with functional neural circuits which become aberrant if cell sources from patients with neurodevelopmental disorders are used.
The cortico-striatal circuit can also be recapitulated in microfluidic devices, as a minimal model of striatal atrophy and network hypersynchronization in Huntington disease144. Similarly, a human nigro-striatal pathway-on-a-chip can be established on a compartmentalized microfluidic platform145. In addition, models of the cortico-striatal and nigro-striatal pathways can be bioprinted using bioinks infused with cells that are reprogrammed to express specific phenotypes in response to cocktails of transcription factors133.
Thalamus
CNS modelling trends towards the development of systems and pathways, and therefore models of the thalamus may enable more complex bioengineering owing to its role as a relay point, gating centre and modulator of sensory information. Thalamic neurons can be generated from mouse ES cells exposed to bone morphogenetic protein 7 (BMP7), yielding phenotypically mature cells that can innervate organotypic cultures and brains in vivo, when implanted subcortically146. Thalamic organoids can then be generated from human ES cells fused with cortical organoids to recapitulate cortico-thalamic circuits147. Remarkably, retinal organoids coupled to cortico-thalamic organoids can model visual pathway development148. Although bioprinted and scaffold-based models of the thalamus have not yet been reported, brain-on-a-chip models have been developed that contain cortical and thalamic co-cultures149. In such microfluidic devices with integrated MEAs, the activity of cortical ensembles can be monitored, which is differentially modulated by thalamic and hippocampal inputs150.
Hypothalamus
Hypothalamic nuclei regulate autonomic functions, such as satiety, sexual behaviour, aggression, metabolism and circadian rhythms. They also interface with vasculature, endocrine organs and the cerebral ventricles to detect changes in temperature, soluble chemicals and pressure, among other signals. Mouse ES cell-derived neuroectodermal cells can be driven towards neural progenitors with hypothalamic phenotypes by removal of factors that promote growth and tissue patterning151. Similarly, hypothalamic neurons with ventricular-like interfacial features can be derived from human ES and iPS cells; these neurons express oxytocin, vasopressin, corticotropin-releasing hormone (CRH) and thyrotropin-releasing hormone, among others152. Directed differentiation protocols are also available to generate specific neuropeptidergic cell subtypes153 and nucleus-specific hypothalamic organoids154,155,156.
Organoids expressing molecular markers characteristic of the arcuate nucleus can be generated with iPS cells from healthy donors and patients with Prader–Willi syndrome154, displaying transcriptomic dysfunctions consistent with the disorder in vivo. In addition, 3D pituitary models157,158 may pave the way for neuroendocrine hybrids159. Human iPS cells can also be differentiated into functional hypothalamic–pituitary units that respond to hypoglycaemic conditions by secreting adrenocorticotropic hormone (ACTH), thus recapitulating the CRH–ACTH stress pathway160. Alternatively, brain-on-a-chip neuroendocrine systems are being explored161.
Brainstem
CNS models of the medulla, pons and the midbrain remain limited; however, owing to the clinical relevance of the substantia nigra and the ventral tegmental area as key anatomical correlates of neuropsychiatric disorders, such as Parkinson disease and schizophrenia, brainstem organoids have been developed162,163. These midbrain-like organoids express neuromelanin pigmentation and dopaminergic profiles consistent with substantia nigra structure and function. Midbrain organoids can recapitulate pathophysiological characteristics of Parkinson disease, including network dysfunction and α-synuclein aggregation164,165.
Heterogeneous brainstem organoids can be created from human iPS cells with a strong midbrain-like phenotype and dopaminergic cells166. Brainstem organoids express specific cholinergic (choline acetyltransferase (ChaT)) and noradrenergic markers (dopamine β-hydroxylase) as well as hindbrain-specific genes (zinc finger of the cerebellum 1 (ZIC1), ZIC4), and therefore likely contain pontine and medullary cells. Moreover, hindbrain organoids can be derived from human iPS cells and serotonergic cells with genetic profiles congruent with genes expressed in the raphe nuclei of the medulla167. Engineering-based models of the brainstem have not yet been developed.
Cerebellum
The cerebellum, similar to the cerebrum, comprises several embedded nuclei surrounded by dense tract systems and a thin, three-layered outer shell of cortical tissue with high gyrification; however, its unique microcircuitry greatly differs from that of the cerebral cortex. Engineering-based models of the cerebellum have not yet been developed, but bioprinting strategies, explored for the generation of laminar morphology and gyrification in the cerebral cortex, may also be adapted for cerebellar modelling.
Cerebellar organoids with self-organized and polarized 3D microanatomy can be generated using human ES cells with populations of functional Purkinje cells; however, their morphologies remain phenotypically embryonic with neural tube-like features168. Alternatively, spinning bioreactors can be integrated to generate cerebellar organoids with mature cytoarchitectures169. In addition, spheroids and organoids can be created by exposing human iPS cells to morphogens, including retinoic acid, Wingless, Int-1 signals (WNT) and sonic hedgehog (SHH), which promote cerebellar differentiation. These cell biology-based cerebellum models express markers of multiple cortical layers and show normal electrophysiological signatures of Purkinje cell function170.
Spinal cord
Modelling the spinal cord is particularly interesting for the development of regenerative therapies for spinal cord dissection, degeneration, atrophy and other pathologies. The spinal cord has a rather simple internal cytoarchitecture and circuitry, and thus both cell biology-based and engineering-based models of the spinal cord could be developed.
Spinal cord tissues can be 3D-printed using a fibrin-based bioink infused with differentiated neural progenitor cells derived from human iPS cells, ultimately expressing lower motor neuron markers indicative of neural cell fates consistent with the ventral spinal cord171. Similarly, chitosan, hyaluronan and Matrigel can provide fast-gelling bioinks to construct implantable, NSC-laden, 3D-printed scaffolds to promote axonal regeneration and decrease scarring after an experimental spinal cord injury in living rodent models172. A scaffold–organoid hybrid method can be bioprinted using a gelatin-based, enzymatically cross-linked hydrogel bioink infused with boundary cap neural crest stem cells to construct 3D spinal cord tissue173. Importantly, bioprinting allows the fabrication of cell type-specific tissues174 with single-cell precision and the infusion of printed scaffolds with growth factors and signalling molecules58, thus potentially enabling the construction of 3D spinal circuits that underlie the basic reflexes involving compartmentalized motor neurons, interneurons and sensory neurons.
Lower motor neuron-containing neuromuscular junction circuits can be designed on microfluidic devices175,176, for example, using optogenetically excitable motor neurons differentiated from mouse ES cells within a 3D collagen–Matrigel hydrogel. Such systems allow the study of degenerative disorders, such as amyotrophic lateral sclerosis (ALS) and spinal muscular atrophy177. In particular, compartmentalized 3D ALS-on-a-chip models are promising tools for clinical research178,179. More complex, highly parallelized spinal circuits can also be generated in microfluidic devices by guiding the axons of multiple motor neuron spheroids towards myofibre targets180.
In addition, cerebral organoid protocols can be adapted to generate spinal cord organoids181 with motor neurons, interneurons, spinal astrocytes and embedded morphogen gradients, consistent with the rostro-caudal axes of the ventral horns of spinal cords in vivo. Interestingly, by modulating concentrations of BMP4 and SHH, dorsal and ventral-intermediate spinal cord organoids can be generated with distinct sensory and motor neuron populations, respectively182. Thus, sensory-motor circuitry could be realized by fusing dorsal and ventral segments183. Such modular tissues can be combined to form complex, neuromuscular assembloids to study neural tube defects184.
Retinal tissues
The retina is a multilayered neural tissue situated deep within the eye. The retina contains various neuronal subtypes, photoreceptive cells and ganglion cells, whose axons form the optic nerves that initiate signalling within the visual pathway. The first cell biology-based model of the retina was generated by aggregation of dissociated cells from mouse and chick tissues into internally laminated retinospheroids185. Additional tissue patterning, including pigmentation and centre-surround polarization, was later achieved in retinal organoids derived from ES cells by stepwise differentiation protocols with transient morphogen exposures186. Retinal organoids can also be further functionalized with photoreceptors that actively respond to light.
The migratory behaviour of human and non-human retinal lineage cells can be explored in microfluidic devices that contain chemical gradients, tissue geometry and ECM substrates187,188. Moreover, bioprinted hydrogels based on hyaluronan can recapitulate biophysical features of the retinal microenvironment, promoting differentiation of photoreceptors from retinal progenitors189. Similarly, photoreceptors can be precisely printed as layers on top of retinal pigment epithelium190. To create tissue-guiding moulds for retinal research, synthetic scaffolds can be 3D-printed with hexagonal arrays of pores using two-photon polymerization; the scaffolds can then be seeded with retinal progenitor cells, which differentiate into retinal neurons191.
Olfactory tissues
Olfactory epithelia, bulbs and nerves transduce chemical energy and transmit signals to distinct nuclei and cortical regions, where smell is ultimately perceived. Although implicated in several psychiatric and neurological disorders, the olfactory epithelium has received little modelling attention thus far, compared with other CNS tissues. Olfactory neurospheroids can serve as models of schizophrenia192, autism spectrum disorder (ASD)193 and Parkinson disease194. In addition, bioprinted scaffold-based models195 and organoid models of the olfactory epithelium are being explored196,197.
Neural-X interfaces
Muscular
Neuromuscular interfaces underlie all behaviours, including speech, facial expression and basic spinal reflexes. Unlike in living organisms, disembodied 2D or 3D neural tissues are unable to physically interact with their environments. Therefore, investigations of behavioural phenomena characteristic of neurological disorders have historically been limited to the study of animals. Alternatively, neuromuscular assembloids can provide in vitro platforms for the investigation of corticospinal function, featuring definable upper (cortical) and lower (spinal) motor organoids as well as excitable musculature55. In these 3D assembloids of the pyramidal tracts, key features of diseases of the neuromuscular junction, such as myasthenia gravis, can be modelled198. Similarly, cortico-striatal models can recapitulate the extrapyramidal system to study ASD, obsessive-compulsive disorder and Parkinson disease141.
A neuromuscular interface-on-a-chip can be engineered using a microfluidic device and spheroids made from cells from patients with ALS to model axonal regression, motor neuron death and muscle atrophy in vitro179. Similarly, a developmental model of the neuromuscular junction can be generated by combining human skeletal muscle cells and ES cell-derived motor neuron clusters in a hydrogel, followed by seeding in PDMS moulds, which results in self-organized 3D tissues that express cholinergic neurotransmission199. Such engineering-based models200 may serve as high-throughput tools for drug screening and personalized medicine to combat neuromuscular disease.
Sensory
Perceptual modalities, such as vision, hearing and somatosensation, allow organisms to avoid threats and pursue positive fitness outcomes. These modalities rely on the transmission of sensory information from the peripheral nervous system to the CNS. However, the role of sensory input as a determinant of cultured neural network function is often overlooked. Disembodied neurons display physiologically aberrant electrical activity in vitro201, which can be rescued by applying patterned stimulations201,202.
Several 3D neural–sensory interfaces have been developed. For example, forebrain organoids can be engineered by assembling optic vesicles as primordial eye fields with lens-like cells, corneal tissue, retinal progenitor cells and pigment epithelia203. These optic vesicle-containing brain organoids are functional and sensitive to different intensities of photostimulation. In addition, organoid models of the inner ear have been developed that contain functional hair cells204,205. A CNS–peripheral nervous system model, generated with specialized dorsal horn sensory neurons responsive to µ-opioid receptor activators and inhibitors, can be applied to study ascending spinal sensory pathways206. Similarly, neuro-mesodermal assembloids and 3D-bioprinted somatosensory constructs respond to capsaicin and menthol, recapitulating the receptive features of several pathways governing pain, temperature and taste207,208.
Robotic
Neural models can also be functionally coupled to machines. For example, neural networks cultured on MEAs can serve as neural–machine interfaces to control mobile robots209, mechanical limbs201,210, virtual aircrafts in flight simulators211 and virtual animal avatars210. Robots and other non-neural effectors can be equipped with dynamic sensors to provide patterned sensory feedback on cells and tissues contingent upon action212. The resulting sensory-motor feedback loops may underlie basic mechanisms of cognition213,214. These hybrid robots, or ‘hybrots’, can be based on cell monolayers or 3D neural tissues. For example, thick (0.5 mm) organotypic slice cultures can be coupled to robots using MEAs215. 3D CNS models and organotypic slice cultures are similar in scale and equally compatible with available electrical and optical sensors, and therefore hybrots may be controllable by bioengineered neural networks122,216,217 to improve our operational understanding of learning and inspire the design of neuromorphic artificial intelligences122.
Outlook
Bioengineered models of the CNS are powerful tools for basic and applied research. However, many engineering challenges remain to maximize the translational potential of CNS models. Despite the development of promising vascularization strategies, oxygenation and nutrient transport issues remain crucial bottlenecks in cell biology-based models. The seamless integration of functional vessels will likely require micromanagement of tissue patterning to mimic highly parallel and autonomous processes which are typically expressed during embryonic development and later suppressed as organisms mature. Therefore, there is a need to develop engineering approaches that activate intrinsic developmental programmes to achieve integrated tissue phenotypes. Until new perfusion strategies are developed, existing hybrid systems that combine 3D tissues with multi-compartment microfluidic devices and other vascular interfaces can support spheroid and organoid research towards increasingly personalized brain models to assist with the diagnosis and treatment of patients with neuropsychiatric diseases. The brain is among the most vascularized organs in the body and, therefore, vascular interfaces are key to the maturation of CNS models.
Bioprinting strategies, customizable scaffolds, complex microfluidic channels and other engineering-based techniques offer high spatial and temporal control over bioengineered CNS tissues. However, these approaches suffer from printing shear stress, a lack of appropriate bioinks, low cell densities, imprecise tissue architectures and low microenvironmental relevance. In particular, ECM composition and fine structure, including the integration of proteoglycans, perineuronal nets and biophysical cues, remain undervalued. Drawing inspiration from organization principles found in nature, such as molecular self-assembly, disorder to order processes and diffusion reactions, complex material designs could be implemented, ensuring a balance of physiological relevance and tractability for high-throughput applications72. However, model systems remain specialized tools, and multiple approaches are needed to overcome their intrinsic limitations. For example, the physiological relevance of organoids can be combined with the controlled perfusion of microfluidic devices into a superior, hybrid model system.
Engineered CNS tissues are often assembled by self-organization of stem cells, which co-opts endogenous embryonic-like morphogenetic programmes. However, mature neural circuits with postnatal phenotypes218 would better recapitulate the physiology of later stages of ontogeny, including puberty and senescence, with age-matched regenerative competence, and thus substantially improve the translational potential of bioengineered CNS tissues. Indeed, drug development and repurposing, injury repair and even the reversal or suppression of normal ageing processes are dependent upon progress in this area219.
Finally, higher degrees of functional complexity in CNS models, and coupling with sensory and motor interfaces, would allow the in vitro assessment of reflex arcs, associative learning and other rudimentary sensory-motor phenomena. Thus, CNS models may serve as platforms to test hypotheses related to embodied and minimal cognition, elucidating the mechanisms and evolutionary origins of intelligence and other higher-order brain functions. Building brain models with cells and other biological building blocks derived from diverse species may enable comparative analyses of cognitive function in vitro. Bioengineered tissues may also facilitate the design of neuromorphic computers and bioinspired artificial intelligence. Of note, CNS functions also include a capacity for sentience as well as the experiences of pain and suffering, which raises ethical and legal concerns related to embodied or minimally cognitive neural systems123,220. Therefore, ethical and legal frameworks should be developed for bioengineered CNS tissues; however, the full impact of higher functional competence in CNS models may not be fully appreciated until they are built and measured.
Citation diversity statement
The authors acknowledge that papers authored by scholars from minoritized groups are systematically under-cited. Here, we have made every attempt to reference relevant papers in a manner that is equitable in terms of racial, ethnic, gender and geographical representation.
References
Koch, C. What is consciousness. Nature 557, S8–S12 (2018).
Lewis, A. & Greer, D. Current controversies in brain death determination. Nat. Rev. Neurol. 13, 505–509 (2017).
Jia, J. et al. The cost of Alzheimer’s disease in China and re‐estimation of costs worldwide. Alzheimers Dement. 14, 483–491 (2018).
Hassabis, D., Kumaran, D., Summerfield, C. & Botvinick, M. Neuroscience-inspired artificial intelligence. Neuron 95, 245–258 (2017).
Ericsson, A. C., Crim, M. J. & Franklin, C. L. A brief history of animal modeling. Mo. Med. 110, 201 (2013).
Hodgkin, A. L. & Katz, B. The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. 108, 37 (1949).
Hartung, T. Thoughts on limitations of animal models. Parkinsonism Relat. Disord. 14, S81–S83 (2008).
Golgi, C. Sulla sostanza grigia del cervello [Italian]. Gazz. Med. Ital. 33, 244–246 (1873).
Bemmelen, J. M. V. Der Hydrogel und das kristallinische Hydrat des Kupferoxydes [German]. Z. Anorg, Chem. 5, 466 (1894).
Harrison, R. G. The outgrowth of the nerve fiber as a mode of protoplasmic movement. J. Exp. Zool. 9, 787–846 (1910).
Millet, L. J. & Gillette, M. U. Over a century of neuron culture: from the hanging drop to microfluidic devices. Yale J. Biol. Med. 85, 501 (2012).
Till, J. E. & McCulloch, E. A. A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat. Res. 14, 213–222 (1961).
Wichterle, O. & Lim, D. Hydrophilic gels for biological use. Nature 185, 117–118 (1960).
Choi, J. S., Lee, H. J., Rajaraman, S. & Kim, D. H. Recent advances in three-dimensional microelectrode array technologies for in vitro and in vivo cardiac and neuronal interfaces. Biosens. Bioelectron. 171, 112687 (2021). This review discusses state-of-the-art 3D MEA technologies that are enabling deep-tissue functional recordings to assess network properties of bioengineered CNS models.
Pine, J. Recording action potentials from cultured neurons with extracellular microcircuit electrodes. J. Neurosci. Methods 2, 19–31 (1980).
Ma, W. et al. CNS stem and progenitor cell differentiation into functional neuronal circuits in three-dimensional collagen gels. Exp. Neurol. 190, 276–288 (2004).
Terry, S. C., Jerman, J. H. & Angell, J. B. A gas chromatographic air analyzer fabricated on a silicon wafer. IEEE Trans. Electron. Devices 26, 1880–1886 (1979).
Oddo, A. et al. Advances in microfluidic blood–brain barrier (BBB) models. Trends Biotechnol. 37, 1295–1314 (2019).
Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676 (2006).
Yu, J. et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 318, 1917–1920 (2007).
Lancaster, M. A. et al. Cerebral organoids model human brain development and microcephaly. Nature 501, 373–379 (2013). This paper reports whole-brain cerebral organoid development, providing a foundation for current innovations in region-specific organoid models and assembloids.
Vieira de Sa, R., Canizares Luna, M. & Pasterkamp, R. J. Advances in central nervous system organoids: a focus on organoid-based models for motor neuron disease. Tissue Eng. C Methods 27, 213–224 (2021).
Trujillo, C. A. et al. Complex oscillatory waves emerging from cortical organoids model early human brain network development. Cell Stem Cell 25, 558–569 (2019). This article demonstrates that organoids display electrical activity with complex oscillations characteristic of the developing brains of neonates, thus confirming a functional recapitulation of in vivo human brain dynamics, in vitro.
Dingle, Y. T. L. et al. Functional characterization of three-dimensional cortical cultures for in vitro modeling of brain networks. iScience 23, 101434 (2020).
Tang-Schomer, M. D. et al. Bioengineered functional brain-like cortical tissue. Proc. Natl Acad. Sci. USA 111, 13811–13816 (2014). This paper presents the first functional, multi-compartmental, scaffold-based cortical CNS model comprising primary cortical neurons from rodents embedded in a porous silk fibroin scaffold and collagen hydrogel.
Potjewyd, G., Moxon, S., Wang, T., Domingos, M. & Hooper, N. M. Tissue engineering 3D neurovascular units: a biomaterials and bioprinting perspective. Trends Biotechnol. 36, 457–472 (2018).
Lovett, M. L., Nieland, T. J., Dingle, Y. T. L. & Kaplan, D. L. Innovations in 3D tissue models of human brain physiology and diseases. Adv. Funct. Mater. 30, 1909146 (2020).
Zhuang, P., Sun, A. X., An, J., Chua, C. K. & Chew, S. Y. 3D neural tissue models: from spheroids to bioprinting. Biomaterials 154, 113–133 (2018).
Baddeley, A. D. & Della Sala, S. Working memory and executive control. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 351, 1397–1404 (1996).
Dolan, R. J. & Fletcher, P. C. Dissociating prefrontal and hippocampal function in episodic memory encoding. Nature 388, 582–585 (1997).
Cotterill, R. M. Cooperation of the basal ganglia, cerebellum, sensory cerebrum and hippocampus: possible implications for cognition, consciousness, intelligence and creativity. Prog. Neurobiol. 64, 1–33 (2001).
Sherman, S. M. Thalamus plays a central role in ongoing cortical functioning. Nat. Neurosci. 19, 533–541 (2016).
Seoane-Collazo, P. et al. Hypothalamic-autonomic control of energy homeostasis. Endocrine 50, 276–291 (2015).
Buckner, R. L. The cerebellum and cognitive function: 25 years of insight from anatomy and neuroimaging. Neuron 80, 807–815 (2013).
Craven, J. Major ascending and descending tracts in the spinal cord. Anaesth. Intensive Care Med. 12, 26–27 (2011).
Rauti, R., Renous, N. & Maoz, B. M. Mimicking the brain extracellular matrix in vitro: a review of current methodologies and challenges. Isr. J. Chem. 60, 1141–1151 (2020).
Fawcett, J. W., Oohashi, T. & Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 20, 451–465 (2019).
Daneman, R. & Prat, A. The blood–brain barrier. Cold Spring Harb. Perspect. Biol. 7, a020412 (2015).
Sarter, M., Bruno, J. P. & Parikh, V. Abnormal neurotransmitter release underlying behavioral and cognitive disorders: toward concepts of dynamic and function-specific dysregulation. Neuropsychopharmacology 32, 1452–1461 (2007).
Lake, B. B. et al. Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science 352, 1586–1590 (2016).
Luo, C. et al. Single-cell methylomes identify neuronal subtypes and regulatory elements in mammalian cortex. Science 357, 600–604 (2017).
Masland, R. H. Neuronal cell types. Curr. Biol. 14, R497–R500 (2004).
Rahman, T. et al. Effect of immune activation during early gestation or late gestation on inhibitory markers in adult male rats. Sci. Rep. 10, 1–12 (2020).
Agboola, O. S., Hu, X., Shan, Z., Wu, Y. & Lei, L. Brain organoid: a 3D technology for investigating cellular composition and interactions in human neurological development and disease models in vitro. Stem Cell Res. Ther. 12, 1–16 (2021).
Sharma, A., Sances, S., Workman, M. J. & Svendsen, C. N. Multi-lineage human iPSC-derived platforms for disease modeling and drug discovery. Cell Stem Cell 26, 309–329 (2020).
Alić, I. et al. Patient-specific Alzheimer-like pathology in trisomy 21 cerebral organoids reveals BACE2 as a gene dose-sensitive AD suppressor in human brain. Mol. Psychiatry 26, 5766–5788 (2021).
Agoglia, R. M. et al. Primate cell fusion disentangles gene regulatory divergence in neurodevelopment. Nature 592, 421–427 (2021).
Gokhman, D. et al. Human–chimpanzee fused cells reveal cis-regulatory divergence underlying skeletal evolution. Nat. Genet. 53, 467–476 (2021).
Trujillo, C. A. et al. Reintroduction of the archaic variant of NOVA1 in cortical organoids alters neurodevelopment. Science 371, eaax2537 (2021). This article reports the creation of brain organoids with archaic gene variants found in non-human hominins (Neanderthals and Denisovans), enabling unprecedented comparative analyses of traits separating humans and extinct relatives in 3D CNS tissues.
Hopkins, A. M. et al. Silk hydrogels as soft substrates for neural tissue engineering. Adv. Funct. Mater. 23, 5140–5149 (2013).
Kim, D. H., Wong, P. K., Park, J., Levchenko, A. & Sun, Y. Microengineered platforms for cell mechanobiology. Annu. Rev. Biomed. Eng. 11, 203–233 (2009).
Doblado, L. R., Martínez-Ramos, C. & Pradas, M. M. Biomaterials for neural tissue engineering. Front. Nanotechnol. 3, 643507 (2021). This review provides a comprehensive and up-to-date discussion of biomaterials and synthetic materials for neural tissue engineering.
Ai, J. et al. Polymeric scaffolds in neural tissue engineering: a review. Arch. Neurosci. 1, 15–20 (2014).
Miura, Y. et al. Engineering brain assembloids to interrogate human neural circuits. Nat. Protoc. 17, 15–35 (2022). This paper presents methods to generate 3D CNS assembloid circuits with region-specific phenotypes that can be interrogated using optogenetic and MEA technologies.
Andersen et al. Generation of functional human 3D cortico-motor assembloids. Cell 183, 1913–1929 (2020).
Kato‐Negishi, M., Morimoto, Y., Onoe, H. & Takeuchi, S. Millimeter‐sized neural building blocks for 3D heterogeneous neural network assembly. Adv. Healthc. Mater. 2, 1564–1570 (2013).
Zhang, J. et al. Single cell bioprinting with ultrashort laser pulses. Adv. Funct. Mater. 31, 2100066 (2021).
Cadena, M. et al. 3D bioprinting of neural tissues. Adv. Healthc. Mater. 10, 2001600 (2021).
Sundararaghavan, H. G., Masand, S. N. & Shreiber, D. I. Microfluidic generation of haptotactic gradients through 3D collagen gels for enhanced neurite growth. J. Neurotrauma 28, 2377–2387 (2011).
Adams, J. W., Cugola, F. R. & Muotri, A. R. Brain organoids as tools for modeling human neurodevelopmental disorders. Physiology 34, 365–375 (2019).
Seale, K., Horvath, S., Teschendorff, A., Eynon, N. & Voisin, S. Making sense of the ageing methylome. Nat. Rev. Genet. 23, 585–605 (2022).
Jgamadze, D. et al. Modeling traumatic brain injury with human brain organoids. Curr. Opin. Biomed. Eng. 14, 52–58 (2020).
Takamura, T. et al. Influence of age on global and regional brain stiffness in young and middle‐aged adults. J. Magn. Reson. Imaging 51, 727–733 (2020).
Abramczyk, H. & Imiela, A. The biochemical, nanomechanical and chemometric signatures of brain cancer. Spectrochim. Acta A Mol. Biomol. Spectrosc. 188, 8–19 (2018).
Miller, J. D. et al. Human iPSC-based modeling of late-onset disease via progerin-induced aging. Cell Stem Cell 13, 691–705 (2013).
Decker, Y. et al. Decreased pH in the aging brain and Alzheimer’s disease. Neurobiol. Aging 101, 40–49 (2021).
Indahlastari, A. et al. Modeling transcranial electrical stimulation in the aging brain. Brain Stimul. 13, 664–674 (2020).
Sack, I. et al. The impact of aging and gender on brain viscoelasticity. Neuroimage 46, 652–657 (2009).
Zhou, W. et al. Dysfunction of the glymphatic system might be related to iron deposition in the normal aging brain. Front. Aging Neurosci. 12, 559603 (2020).
Gao, P. & Ganguli, S. On simplicity and complexity in the brave new world of large-scale neuroscience. Curr. Opin. Neurobiol. 32, 148–155 (2015).
Wu, Y. et al. A neuronal wiring platform through microridges for rationally engineered neural circuits. APL Bioeng. 4, 046106 (2020).
Azevedo, H. S. & Mata, A. Embracing complexity in biomaterials design. Biomater. Biosyst. 6, 100043 (2022).
Marton, R. M. & Pașca, S. P. Organoid and assembloid technologies for investigating cellular crosstalk in human brain development and disease. Trends Cell Biol. 30, 133–143 (2020).
Rey, F. et al. Advances in tissue engineering and innovative fabrication techniques for 3-D-structures: translational applications in neurodegenerative diseases. Cells 9, 1636 (2020).
Mostajo-Radji, M. A., Schmitz, M. T., Montoya, S. T. & Pollen, A. A. Reverse engineering human brain evolution using organoid models. Brain Res. 1729, 146582 (2020).
Andrews, M. G. & Kriegstein, A. R. Challenges of organoid research. Annu. Rev. Neurosci. 45, 23–39 (2022).
Lee, D., Pathak, S. & Jeong, J. H. Design and manufacture of 3D cell culture plate for mass production of cell-spheroids. Sci. Rep. 9, 1–8 (2019).
Jeong, M. et al. Generating inner ear organoids containing putative cochlear hair cells from human pluripotent stem cells. Cell Death Dis. 9, 1–13 (2018).
Shin, N. et al. Vascularization of iNSC spheroid in a 3D spheroid‐on‐a‐chip platform enhances neural maturation. Biotechnol. Bioeng. 119, 566–574 (2022).
Gopinathan, J. & Noh, I. Recent trends in bioinks for 3D printing. Biomater. Res. 22, 1–15 (2018).
Richard, C., Neild, A. & Cadarso, V. J. The emerging role of microfluidics in multi-material 3D bioprinting. Lab Chip 20, 2044–2056 (2020).
Kim, S., Kim, H. J. & Jeon, N. L. Biological applications of microfluidic gradient devices. Integr. Biol. 2, 584–603 (2010).
Karimi, M. et al. Microfluidic systems for stem cell-based neural tissue engineering. Lab Chip 16, 2551–2571 (2016).
Au, A. K., Bhattacharjee, N., Horowitz, L. F., Chang, T. C. & Folch, A. 3D-printed microfluidic automation. Lab Chip 15, 1934–1941 (2015).
Raimondi, I. et al. Organ-on-a-chip in vitro models of the brain and the blood–brain barrier and their value to study the microbiota–gut–brain axis in neurodegeneration. Front. Bioeng. Biotechnol. 7, 435 (2020).
Singh, M. et al. 3D printed conformal microfluidics for isolation and profiling of biomarkers from whole organs. Lab Chip 17, 2561–2571 (2017).
Toepke, M. W. & Beebe, D. J. PDMS absorption of small molecules and consequences in microfluidic applications. Lab Chip 6, 1484–1486 (2006).
Vishwakarma, S. K., Lakkireddy, C., Bardia, A., Paspala, S. A. B. & Khan, A. A. Engineering bio-mimetic humanized neurological constructs using acellularized scaffolds of cryopreserved meningeal tissues. Mater. Sci. Eng. C. 102, 34–44 (2019).
Zaszczyńska, A. et al. Influence of process–material conditions on the structure and biological properties of electrospun polyvinylidene fluoride fibers. Bull. Pol. Acad. Sci. Tech. Sci. 68, 627–633 (2020).
Wang, P. et al. 3D bioprinting of hydrogels for retina cell culturing. Bioprinting 12, e00029 (2018).
Gopalakrishnan, A., Shankarappa, S. A. & Rajanikant, G. K. Hydrogel scaffolds: towards restitution of ischemic stroke-injured brain. Transl. Stroke Res. 10, 1–18 (2019).
Marchini, A. & Gelain, F. Synthetic scaffolds for 3D cell cultures and organoids: applications in regenerative medicine. Crit. Rev. Biotechnol. 42, 468–486 (2022).
Boni, R., Ali, A., Shavandi, A. & Clarkson, A. N. Current and novel polymeric biomaterials for neural tissue engineering. J. Biomed. Sci. 25, 1–21 (2018).
Schizas, N. et al. Hyaluronic acid-based hydrogel enhances neuronal survival in spinal cord slice cultures from postnatal mice. J. Biomater. Appl. 28, 825–836 (2014).
Zargar, V., Asghari, M. & Dashti, A. A review on chitin and chitosan polymers: structure, chemistry, solubility, derivatives, and applications. ChemBioEng Rev. 2, 204–226 (2015).
Ojeda-Hernández, D. D., Canales-Aguirre, A. A., Matias-Guiu, J., Gomez-Pinedo, U. & Mateos-Díaz, J. C. Potential of chitosan and its derivatives for biomedical applications in the central nervous system. Front. Bioeng. Biotechnol. 8, 389 (2020).
Rouleau, N. et al. A long‐living bioengineered neural tissue platform to study neurodegeneration. Macromol. Biosci. 20, 2000004 (2020).
Kook, G. et al. Wafer-scale multilayer fabrication for silk fibroin-based microelectronics. ACS Appl. Mater. Interfaces 11, 115–124 (2018).
Kargozar, S., Singh, R. K., Kim, H. W. & Baino, F. “Hard” ceramics for “soft” tissue engineering: paradox or opportunity? Acta Biomater. 115, 1–28 (2020).
Yang, F., Murugan, R., Wang, S. & Ramakrishna, S. Electrospinning of nano/micro scale poly(L-lactic acid) aligned fibers and their potential in neural tissue engineering. Biomaterials 26, 2603–2610 (2005).
George, J., Hsu, C. C., Nguyen, L. T. B., Ye, H. & Cui, Z. Neural tissue engineering with structured hydrogels in CNS models and therapies. Biotechnol. Adv. 42, 107370 (2020).
Chapla, R., Alhaj Abed, M. & West, J. Modulating functionalized poly(ethylene glycol) diacrylate hydrogel mechanical properties through competitive crosslinking mechanics for soft tissue applications. Polymers 12, 3000 (2020).
Xie, J., MacEwan, M. R., Schwartz, A. G. & Xia, Y. Electrospun nanofibers for neural tissue engineering. Nanoscale 2, 35–44 (2010).
Yu, T. T. & Shoichet, M. S. Guided cell adhesion and outgrowth in peptide-modified channels for neural tissue engineering. Biomaterials 26, 1507–1514 (2005).
Hu, X. et al. Biodegradable poly(lactic acid-co-trimethylene carbonate)/chitosan microsphere scaffold with shape-memory effect for bone tissue engineering. Colloids Surf. B Biointerfaces 195, 111218 (2020).
Lee, S. J. et al. Advances in 3D bioprinting for neural tissue engineering. Adv. Biosyst. 2, 1700213 (2018).
Sharma, D. et al. Polydopamine and collagen coated micro-grated polydimethylsiloxane for human mesenchymal stem cell culture. Bioact. Mater. 4, 142–150 (2019).
Pham, M. T. et al. Generation of human vascularized brain organoids. Neuroreport 29, 588 (2018).
Cho, A. N. et al. Microfluidic device with brain extracellular matrix promotes structural and functional maturation of human brain organoids. Nat. Commun. 12, 1–23 (2021).
Zilles, K. & Amunts, K. Centenary of Brodmann’s map — conception and fate. Nat. Rev. Neurosci. 11, 139–145 (2010).
Boly, M. et al. Are the neural correlates of consciousness in the front or in the back of the cerebral cortex? Clinical and neuroimaging evidence. J. Neurosci. 37, 9603–9613 (2017).
Matsui, T. K., Tsuru, Y., Hasegawa, K. & Kuwako, K. I. Vascularization of human brain organoids. Stem Cell 39, 1017–1024 (2021).
Gordon, A. et al. Long-term maturation of human cortical organoids matches key early postnatal transitions. Nat. Neurosci. 24, 331–342 (2021).
Cantley, W. L. et al. Functional and sustainable 3D human neural network models from pluripotent stem cells. ACS Biomater. Sci. Eng. 4, 4278–4288 (2018).
Shi, Y. et al. Vascularized human cortical organoids (vOrganoids) model cortical development in vivo. PLoS Biol. 18, e3000705 (2020). This paper presents vascularized human cortical organoids with mesh-like, embedded vessels comprising co-cultured human umbilical endothelial cells, which accelerates tissue maturation relative to non-vascularized tissues.
Dong, X. et al. Human cerebral organoids establish subcortical projections in the mouse brain after transplantation. Mol. Psychiatry 26, 2964–2976 (2021).
Liaudanskaya, V. et al. Engineering advanced neural tissue constructs to mitigate acute cerebral inflammation after brain transplantation in rats. Biomaterials 192, 510–522 (2019).
Cakir, B. et al. Engineering of human brain organoids with a functional vascular-like system. Nat. Methods 16, 1169–1175 (2019).
Struzyna, L. A. et al. Rebuilding brain circuitry with living micro-tissue engineered neural networks. Tissue Eng. A 21, 2744–2756 (2015).
Song, L. et al. Assembly of human stem cell-derived cortical spheroids and vascular spheroids to model 3-D brain-like tissues. Sci. Rep. 9, 1–16 (2019).
Rouleau, N., Cairns, D. M., Rusk, W., Levin, M. & Kaplan, D. L. Learning and synaptic plasticity in 3D bioengineered neural tissues. Neurosci. Lett. 750, 135799 (2021). This article reports the first demonstration of a learning paradigm adapted to an engineering-based 3D CNS model, where bioengineered neural tissues display electrophysiological and biomolecular signatures of habituation and synaptic plasticity.
Rouleau, N., Murugan, N. J. & Kaplan, D. L. Toward studying cognition in a dish. Trends Cognit. Sci. 25, 294–304 (2021).
Lavazza, A. ‘Consciousnessoids’: clues and insights from human cerebral organoids for the study of consciousness. Neurosci. Conscious. 7, 1–11 (2021).
Cui, H. et al. A novel near-infrared light responsive 4D printed nanoarchitecture with dynamically and remotely controllable transformation. Nano Res. 12, 1381–1388 (2019).
Karzbrun, E., Kshirsagar, A., Cohen, S. R., Hanna, J. H. & Reiner, O. Human brain organoids on a chip reveal the physics of folding. Nat. Phys. 14, 515–522 (2018).
Budday, S., Steinmann, P. & Kuhl, E. The role of mechanics during brain development. J. Mech. Phys. Solids 72, 75–92 (2014).
Colgin, L. L. Theta–gamma coupling in the entorhinal–hippocampal system. Curr. Opin. Neurobiol. 31, 45–50 (2015).
Nakazono, T. et al. Impaired in vivo gamma oscillations in the medial entorhinal cortex of knock-in Alzheimer model. Front. Syst. Neurosci. 11, 48 (2017).
Scoville, W. B. & Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 20, 11 (1957).
Xu, T. et al. Electrophysiological characterization of embryonic hippocampal neurons cultured in a 3D collagen hydrogel. Biomaterials 30, 4377–4383 (2009).
Wei, Y. T. et al. Hyaluronic acid hydrogel modified with nogo-66 receptor antibody and poly(L-lysine) enhancement of adherence and survival of primary hippocampal neurons. J. Bioact. Compat. Polym. 24, 205–219 (2009).
Zhou, Z., Yu, P., Geller, H. M. & Ober, C. K. The role of hydrogels with tethered acetylcholine functionality on the adhesion and viability of hippocampal neurons and glial cells. Biomaterials 33, 2473–2481 (2012).
Qiu, B. et al. Bioprinting neural systems to model central nervous system diseases. Adv. Funct. Mater. 30, 1910250 (2020).
Zhang, Q. et al. Preparation of uniaxial multichannel silk fibroin scaffolds for guiding primary neurons. Acta Biomater. 8, 2628–2638 (2012).
Bhattacharjee, N. & Folch, A. Large-scale microfluidic gradient arrays reveal axon guidance behaviors in hippocampal neurons. Microsyst. Nanoeng. 3, 1–14 (2017).
Kang, M. S. et al. Graphene oxide-functionalized nanofibre composite matrices to enhance differentiation of hippocampal neuronal cells. Mater. Adv. 1, 3496–3506 (2020).
Lenoir, S. et al. Recreating mouse cortico-hippocampal neuronal circuit in microfluidic devices to study BDNF axonal transport upon glucocorticoid treatment. STAR Protoc. 2, 100382 (2021).
Sakaguchi, H. et al. Generation of functional hippocampal neurons from self-organizing human embryonic stem cell-derived dorsomedial telencephalic tissue. Nat. Commun. 6, 1–11 (2015).
Jacob, F. et al. Human pluripotent stem cell-derived neural cells and brain organoids reveal SARS-CoV-2 neurotropism predominates in choroid plexus epithelium. Cell Stem Cell 27, 937–950 (2020).
Pomeshchik, Y. et al. Human iPSC-derived hippocampal spheroids: an innovative tool for stratifying Alzheimer disease patient-specific cellular phenotypes and developing therapies. Stem Cell Rep. 15, 256–273 (2020).
Obeso, J. A., Rodriguez-Oroz, M. C., Stamelou, M., Bhatia, K. P. & Burn, D. J. The expanding universe of disorders of the basal ganglia. Lancet 384, 523–531 (2014).
Schapira, A. H. Neurobiology and treatment of Parkinson’s disease. Trends Pharmacol. Sci. 30, 41–47 (2009).
Miura, Y. et al. Generation of human striatal organoids and cortico-striatal assembloids from human pluripotent stem cells. Nat. Biotechnol. 38, 1421–1430 (2020).
Virlogeux, A. et al. Reconstituting corticostriatal network on-a-chip reveals the contribution of the presynaptic compartment to Huntington’s disease. Cell Rep. 22, 110–122 (2018).
Kajtez, J. et al. 3D‐Printed soft lithography for complex compartmentalized microfluidic neural devices. Adv. Sci. 7, 2001150 (2020).
Shiraishi, A., Muguruma, K. & Sasai, Y. Generation of thalamic neurons from mouse embryonic stem cells. Development 144, 1211–1220 (2017).
Xiang, Y. et al. hESC-derived thalamic organoids form reciprocal projections when fused with cortical organoids. Cell Stem Cell 24, 487–497 (2019).
Fligor, C. M. et al. Extension of retinofugal projections in an assembled model of human pluripotent stem cell-derived organoids. Stem Cell Rep. 16, 2228–2241 (2021).
Kanagasabapathi, T. T. et al. Functional connectivity and dynamics of cortical–thalamic networks co-cultured in a dual compartment device. J. Neural Eng. 9, 036010 (2012).
Brofiga, M., Pisano, M., Callegari, F. & Massobrio, P. Exploring the contribution of thalamic and hippocampal input on cortical dynamics in a brain-on-a-chip model. IEEE Trans. Med. Robot. Bionics 3, 315–327 (2021).
Wataya, T. et al. Minimization of exogenous signals in ES cell culture induces rostral hypothalamic differentiation. Proc. Natl Acad. Sci. USA 105, 11796–11801 (2008).
Merkle, F. T. et al. Generation of neuropeptidergic hypothalamic neurons from human pluripotent stem cells. Development 142, 633–643 (2015).
Ogawa, K. et al. Vasopressin-secreting neurons derived from human embryonic stem cells through specific induction of dorsal hypothalamic progenitors. Sci. Rep. 8, 1–10 (2018).
Huang, W. K. et al. Generation of hypothalamic arcuate organoids from human induced pluripotent stem cells. Cell Stem Cell 28, 1657–1670 (2021).
Ozaki, H., Suga, H. & Arima, H. Hypothalamic–pituitary organoid generation through the recapitulation of organogenesis. Dev. Growth Differ. 63, 154–165 (2021).
Qian, X. et al. Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nat. Protoc. 13, 565–580 (2018).
Cox, B. et al. Organoids from pituitary as a novel research model toward pituitary stem cell exploration. J. Endocrinol. 240, 287–308 (2019).
Zhou, Y. et al. Pituitary lineage differentiation from human induced pluripotent stem cells in 2D and 3D cultures. Stem Cell Dev. 31, 239–249 (2022).
Gokyurek, M., Yilmaz, K. B. & Yilgor Huri, P. 3D bioprinting for the endocrine glands. Emergent Mater. 3, 441–452 (2020).
Kasai, T. et al. Hypothalamic contribution to pituitary functions is recapitulated in vitro using 3D-cultured human iPS cells. Cell Rep. 30, 18–24 (2020).
Schwerdtfeger, L. A. & Tobet, S. A. From organotypic culture to body‐on‐a‐chip: a neuroendocrine perspective. J. Neuroendocrinol. 31, e12650 (2019).
Jo, J. et al. Midbrain-like organoids from human pluripotent stem cells contain functional dopaminergic and neuromelanin-producing neurons. Cell Stem Cell 19, 248–257 (2016).
Qian, X. et al. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell 165, 1238–1254 (2016).
Jo, J. et al. Lewy body-like inclusions in human midbrain organoids carrying glucocerebrosidase and α‐synuclein mutations. Ann. Neurol. 90, 490–505 (2021).
Mohamed, N. V. et al. Microfabricated disk technology: rapid scale up in midbrain organoid generation. Methods 203, 465–477 (2022).
Eura, N. et al. Brainstem organoids from human pluripotent stem cells. Front. Neurosci. 14, 538 (2020).
Valiulahi, P. et al. Generation of caudal-type serotonin neurons and hindbrain-fate organoids from hPSCs. Stem Cell Rep. 16, 1938–1952 (2021).
Muguruma, K., Nishiyama, A., Kawakami, H., Hashimoto, K. & Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Rep. 10, 537–550 (2015).
Silva, T. P. et al. Scalable generation of mature cerebellar organoids from human pluripotent stem cells and characterization by immunostaining. JoVE 160, e61143 (2020).
Hua, T. et al. Cerebellar differentiation from human stem cells through retinoid, Wnt, and sonic hedgehog pathways. Tissue Eng. A 27, 881–893 (2021).
De la Vega, L. A. et al. 3D bioprinting human induced pluripotent stem cell-derived neural tissues using a novel lab-on-a-printer technology. Appl. Sci. 8, 2414 (2018).
Liu, X. et al. 3D bioprinted neural tissue constructs for spinal cord injury repair. Biomaterials 272, 120771 (2021). This article demonstrates a major development in 3D bioprinting with NSCs to generate anatomically complex cell–scaffold constructs for spinal cord repair, demonstrating high viability, axonal regeneration and reduced glial scarring.
Han, Y. et al. Towards 3D bioprinted spinal cord organoids. Int. J. Mol. Sci. 23, 5788 (2022).
Joung, D. et al. 3D printed stem‐cell derived neural progenitors generate spinal cord scaffolds. Adv. Funct. Mater. 28, 1801850 (2018).
Southam, K. A., King, A. E., Blizzard, C. A., McCormack, G. H. & Dickson, T. C. Microfluidic primary culture model of the lower motor neuron–neuromuscular junction circuit. J. Neurosci. Methods 218, 164–169 (2013).
Taylor, A. M. et al. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2, 599–605 (2005).
Uzel, S. G. et al. Microfluidic device for the formation of optically excitable, three-dimensional, compartmentalized motor units. Sci. Adv. 2, e1501429 (2016).
Nijssen, J., Aguila, J., Hoogstraaten, R., Kee, N. & Hedlund, E. Axon-seq decodes the motor axon transcriptome and its modulation in response to ALS. Stem Cell Rep. 11, 1565–1578 (2018).
Osaki, T., Uzel, S. G. & Kamm, R. D. On-chip 3D neuromuscular model for drug screening and precision medicine in neuromuscular disease. Nat. Protoc. 15, 421–449 (2020).
Machado, C. B. et al. In vitro modeling of nerve–muscle connectivity in a compartmentalized tissue culture device. Adv. Biosyst. 3, 1800307 (2019).
Hor, J. H. et al. Cell cycle inhibitors protect motor neurons in an organoid model of spinal muscular atrophy. Cell Death Dis. 9, 1100 (2018).
Ogura, T., Sakaguchi, H., Miyamoto, S. & Takahashi, J. Three-dimensional induction of dorsal, intermediate and ventral spinal cord tissues from human pluripotent stem cells. Development 145, dev162214 (2018).
Khong, Z. J., Hor, J. H. & Ng, S. Y. Spinal cord organoids add an extra dimension to traditional motor neuron cultures. Neural Regen. Res. 14, 1515–1516 (2019).
Lee, J. H. et al. Production of human spinal-cord organoids recapitulating neural-tube morphogenesis. Nat. Biomed. Eng. 6, 435–448 (2022).
Layer, P. G., Weikert, T. & Willbold, E. Chicken retinospheroids as developmental and pharmacological in vitro models: acetylcholinesterase is regulated by its own and by butyrylcholinesterase activity. Cell Tissue Res. 268, 409–418 (1992).
Kuwahara, A. et al. Generation of a ciliary margin-like stem cell niche from self-organizing human retinal tissue. Nat. Commun. 6, 1–15 (2015).
Mishra, S., Thakur, A., Redenti, S. & Vazquez, M. A model microfluidics-based system for the human and mouse retina. Biomed. Microdevices 17, 1–10 (2015).
Pena, C. D., Zhang, S., Majeska, R., Venkatesh, T. & Vazquez, M. Invertebrate retinal progenitors as regenerative models in a microfluidic system. Cells 8, 1301 (2019).
Wang, L. et al. A human three-dimensional neural-perivascular ‘assembloid’ promotes astrocytic development and enables modeling of SARS-CoV-2 neuropathology. Nat. Med. 27, 1600–1606 (2021).
Masaeli, E. et al. Tissue engineering of retina through high resolution 3-dimensional inkjet bioprinting. Biofabrication 12, 025006 (2020).
Worthington, K. S. et al. Two-photon polymerization for production of human iPSC-derived retinal cell grafts. Acta Biomater. 55, 385–395 (2017).
Naujock, M. et al. Neuronal differentiation of induced pluripotent stem cells from schizophrenia patients in 2D and in 3D cultures reveals increased expression of the Kv4.2 subunit DPP6 that contributes to decreased neuronal activity. Stem Cell Dev. 29, 1577–1587 (2020).
Yang, G. & Shcheglovitov, A. Probing disrupted neurodevelopment in autism using human stem cell‐derived neurons and organoids: an outlook into future diagnostics and drug development. Dev. Dyn. 249, 6–33 (2020).
Matigian, N. et al. Disease-specific, neurosphere-derived cells as models for brain disorders. Dis. Model. Mech. 3, 785–798 (2010).
Othon, C. M., Wu, X., Anders, J. J. & Ringeisen, B. R. Single-cell printing to form three-dimensional lines of olfactory ensheathing cells. Biomed. Mater. 3, 034101 (2008).
Dai, Q. et al. Notch signaling regulates Lgr5+ olfactory epithelium progenitor/stem cell turnover and mediates recovery of lesioned olfactory epithelium in mouse model. Stem Cell 36, 1259–1272 (2018).
Li, X. et al. Age-dependent activation and neuronal differentiation of Lgr5+ basal cells in injured olfactory epithelium via Notch signaling pathway. Front. Aging Neurosci. 12, 602688 (2020).
Martins, J. M. F. et al. Self-organizing 3D human trunk neuromuscular organoids. Cell Stem Cell 26, 172–186 (2020).
Bakooshli, M. A. et al. A 3D culture model of innervated human skeletal muscle enables studies of the adult neuromuscular junction. eLife 8, e44530 (2019).
Rimington, R. P., Fleming, J. W., Capel, A. J., Wheeler, P. C. & Lewis, M. P. Bioengineered model of the human motor unit with physiologically functional neuromuscular junctions. Sci. Rep. 11, 1–15 (2021).
Potter, A. M. et al. in Proc. Brain Inspired Cognitive Systems 1–5 (Univ. of Stirling, 2004).
Madhavan, R., Wagenaar, D. A. & Potter, S. M. Multisite stimulation quiets bursts and enhances plasticity in cultured networks. Soc. Neurosci. Annu. Meet. 1, 45–56 (2003).
Gabriel, E. et al. Human brain organoids assemble functionally integrated bilateral optic vesicles. Cell Stem Cell 28, 1740–1757 (2021).
Jeong, G. S. et al. Networked neural spheroid by neuro-bundle mimicking nervous system created by topology effect. Mol. Brain 8, 1–12 (2015).
Koehler, K. R. et al. Generation of inner ear organoids containing functional hair cells from human pluripotent stem cells. Nat. Biotechnol. 35, 583–589 (2017).
Olmsted, Z. T. & Paluh, J. L. Co-development of central and peripheral neurons with trunk mesendoderm in human elongating multi-lineage organized gastruloids. Nat. Commun. 12, 1–19 (2021).
Hirano, M. et al. 3D bioprinted human iPSC-derived somatosensory constructs with functional and highly purified sensory neuron networks. Biofabrication 13, 035046 (2021).
Rockel, A. F., Wagner, N., Ergün, S. & Wörsdörfer, P. Neuro-mesodermal assembloids recapitulate aspects of peripheral nervous system development in vitro. Preprint at bioRxiv https://doi.org/10.1101/2022.06.30.498240 (2022).
Novellino, A. et al. Connecting neurons to a mobile robot: an in vitro bidirectional neural interface. Comput. Intell. Neurosci. 2007, 12725 (2007).
Bakkum, D. J., Chao, Z. C. & Potter, S. M. Spatio-temporal electrical stimuli shape behavior of an embodied cortical network in a goal-directed learning task. J. Neural Eng. 5, 310 (2008).
DeMarse, T. B. & Dockendorf, K. P. Adaptive flight control with living neuronal networks on microelectrode arrays. Proc. 2005 IEEE Int. Jt. Conf. Neural Netw. 3, 1548–1551 (2005).
Tessadori, J., Bisio, M., Martinoia, S. & Chiappalone, M. Modular neuronal assemblies embodied in a closed-loop environment: toward future integration of brains and machines. Front. Neural Circuits 6, 99 (2012).
Linson, A., Clark, A., Ramamoorthy, S. & Friston, K. The active inference approach to ecological perception: general information dynamics for natural and artificial embodied cognition. Front. Robot. AI 5, 21 (2018).
Potter, S. M., Wagenaar, D. A. & DeMarse, T. B. in Advances in Network Electrophysiology (eds Taketani, M. & Baudry M.) 215–242 (Springer, 2006).
Killian, N. J., Vernekar, V. N., Potter, S. M. & Vukasinovic, J. A device for long-term perfusion, imaging, and electrical interfacing of brain tissue in vitro. Front. Neurosci. 10, 135 (2016).
Abramson, C. I. & Levin, M. Behaviorist approaches to investigating memory and learning: a primer for synthetic biology and bioengineering. Commun. Integr. Biol. 14, 230–247 (2021).
Furusawa, K., Teramae, R., Ohashi, H. & Shimizu, M. Development of living “bio-robots” for autonomous actuations. J. Robot. Mechatron. 34, 279–284 (2022).
Kelley, K. W. & Pașca, S. P. Human brain organogenesis: toward a cellular understanding of development and disease. Cell. 185, 42–61 (2021).
Tan, H. Y., Cho, H. & Lee, L. P. Human mini-brain models. Nat. Biomed. Eng. 5, 11–25 (2021).
Krausová, A., & Hazan, H. Robots with biological brains: autonomy and liability of a semi-artificial life form. The Lawyer Quarterly 7, 162–172 (5 September 2017).
Park, Y. et al. Three-dimensional, multifunctional neural interfaces for cortical spheroids and engineered assembloids. Sci. Adv. 7, eabf9153 (2021).
Cairns, D. M. et al. A 3D human brain-like tissue model of herpes-induced Alzheimer’s disease. Sci. Adv. 6, eaay8828 (2020).
Pereira, J. D. et al. Human sensorimotor organoids derived from healthy and amyotrophic lateral sclerosis stem cells form neuromuscular junctions. Nat. Commun. 12, 1–17 (2021).
Liu, C. et al. Mitochondrial HSF1 triggers mitochondrial dysfunction and neurodegeneration in Huntington’s disease. EMBO Mol. Med. 14, e15851 (2022).
Abdelrahman, S. A. et al. Parkinson’s disease model composed of 3D bioprinted dopaminergic neurons within a biomimetic peptide scaffold. Biofabrication. 14, 044103 (2022).
Smits, L. M. et al. Modeling Parkinson’s disease in midbrain-like organoids. NPJ Parkinsons Dis. 5, 1–8 (2019).
de Jong, J. O. et al. Cortical overgrowth in a preclinical forebrain organoid model of CNTNAP2-associated autism spectrum disorder. Nat. Commun. 12, 1–14 (2021).
Srikanth, P. et al. Shared effects of DISC1 disruption and elevated WNT signaling in human cerebral organoids. Transl. Psychiatry 8, 1–14 (2018).
Sarkar, A. et al. Efficient generation of CA3 neurons from human pluripotent stem cells enables modeling of hippocampal connectivity in vitro. Cell Stem Cell 22, 684–697 (2018).
Mellios, N. et al. MeCP2-regulated miRNAs control early human neurogenesis through differential effects on ERK and AKT signaling. Mol. Psychiatry 23, 1051–1065 (2018).
Blair, J. D., Hockemeyer, D. & Bateup, H. S. Genetically engineered human cortical spheroid models of tuberous sclerosis. Nat. Med. 24, 1568–1578 (2018).
Ko, E., Poon, M. L. S., Park, E., Cho, Y. & Shin, J. H. Engineering 3D cortical spheroids for an in vitro ischemic stroke model. ACS Biomater. Sci. Eng. 7, 3845–3860 (2021).
Wang, S. N. et al. Cerebral organoids repair ischemic stroke brain injury. Transl. Stroke Res. 11, 983–1000 (2020).
Soon, K. et al. A human model of arteriovenous malformation (AVM)-on-a-chip reproduces key disease hallmarks and enables drug testing in perfused human vessel networks. Biomaterials 288, 121729 (2022).
Liaudanskaya, V. et al. Modeling controlled cortical impact injury in 3D brain‐like tissue cultures. Adv. Healthc. Mater. 9, 2000122 (2020).
Li, Y. et al. A precise, controllable in vitro model for diffuse axonal injury through uniaxial stretch injury. Front. Neurosci. 13, 1063 (2019).
Weightman, A. P., Pickard, M. R., Yang, Y. & Chari, D. M. An in vitro spinal cord injury model to screen neuroregenerative materials. Biomaterials 35, 3756–3765 (2014).
Wang, X. et al. 3D bioprinted glioma microenvironment for glioma vascularization. J. Biomed. Mater. Res. A 109, 915–925 (2021).
Altman, J. & Das, G. D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 124, 319–335 (1965).
Min, J. K., Mosadegh, B., Dunham, S. & Al’Aref, S. J. (eds) 3D Printing Applications in Cardiovascular Medicine (Academic, 2018).
Faulkner-Jones, A. et al. Bioprinting of human pluripotent stem cells and their directed differentiation into hepatocyte-like cells for the generation of mini-livers in 3D. Biofabrication 7, 044102 (2015).
Brodmann, K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues [German] (J. A. Barth, 1909).
Martin, G. R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl Acad. Sci. USA 78, 7634–7638 (1981).
Thomson, J. A. et al. Embryonic stem cell lines derived from human blastocysts. Science 282, 1145–1147 (1998).
Svendsen, C. N., Caldwell, M. A. & Ostenfeld, T. Human neural stem cells: isolation, expansion and transplantation. Brain Pathol. 9, 499–513 (1999).
Vierbuchen, T. et al. Direct conversion of fibroblasts to functional neurons by defined factors. Nature 463, 1035–1041 (2010).
Huxley, T. H. An Introduction to the Classification of Animals 6–59 (John Churchill & Sons, 1869).
Acknowledgements
The authors acknowledge support from the National Sciences and Engineering Research Council of Canada (NSERC), Discovery Grants RGPIN-2022-04162 (to N.R.) and RGPIN-2021-03783 (to N.J.M.) as well as the National Institutes of Health (NIH) grants P41EB027062 and R01AG061838 (to D.L.K.). They also thank scientific illustrator L. Hugdahl for her contributions.
Author information
Authors and Affiliations
Contributions
N.R., N.J.M. and D.L.K. contributed equally.
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflicts of competing interest, financial or otherwise.
Peer review
Peer review information
Nature Reviews Bioengineering thanks Anna Herland, Steven Sloan, Melissa Cadena and the other, anonymous, reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Rouleau, N., Murugan, N.J. & Kaplan, D.L. Functional bioengineered models of the central nervous system. Nat Rev Bioeng 1, 252–270 (2023). https://doi.org/10.1038/s44222-023-00027-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s44222-023-00027-7
