
Abstract
Coastal habitats are amongst the most dynamic on Earth, due to their simultaneous exposure to terrestrial, oceanic and atmospheric processes. Coastal taxa are therefore often ecologically specialised and adapted to withstand frequent shifts in sea level, wave exposure, temperature or salinity. This specialisation often resulting in significant cryptic diversity. Previous molecular studies have suggested that genetic differentiation in non-marine coastal organisms may be influenced by oceanic currents and fronts, but the extent to which such processes affect dispersal and evolution of such taxa remains unclear. Here we explore whether population genetic structure in two supralittoral rockpool beetle species (genus Ochthebius) can be predicted from the general circulation pattern of the marine currents and associated oceanic fronts. We simulated dispersal using a Lagrangian particle tracking model and compared this with population genetic structure inferred from COI (mitochondrial) and wingless (nuclear) genes applying linear models and Mantel tests. We show that a biophysical model based on oceanic currents and fronts in the western Mediterranean Sea is a much better predictor of observed population genetic structure than isolation by distance in both species. Our results show that oceanic processes, besides shaping contemporary population connectivity in fully marine organisms, also exert a meaningful influence on terrestrially-derived coastal taxa such as supralittoral rockpool beetles — the first time this mode of dispersal has been demonstrated in an insect.
Similar content being viewed by others
Introduction
Coastlines are ecotones between land and ocean and are amongst the most dynamic environments on Earth. Within these areas, supralittoral rockpools are geographically restricted, fragmented and exposed to processes of physical disturbance operating across a broad variety of temporal and spatial scales. They suffer constant habitat modifications driven by several terrestrial, oceanic and atmospheric processes, and are highly vulnerable to large-scale physical disturbances1. The biological assemblages found in these areas are dominated by strongly adapted species that can withstand frequent shifts in sea level, temperature and salinity2,3. As a consequence, coastal taxa are often ecologically specialised, frequently restricted to specific habitats. These taxa may harbour significant cryptic diversity4,5,6,7 due to ecomorphological constraints and the isolation of populations by major physical coastal disturbances such as uplifts, tsunamis, hurricanes, glaciations and sea-level fluctuations1. Whilst between the tidelines, coastal biota is dominated by marine-derived species, the composition of supralittoral communities is very different. Such habitats are typically species-poor8, but support a suite of specialised, ecologically restricted taxa, many of which are terrestrial or freshwater in origin rather than marine6. Compared to intertidal biotas, the inhabitants of the supralittoral are much less well studied, and the extent to which their population biology and evolution resemble terrestrial or marine organisms remains unclear.
Although major disturbances play an important role in isolating populations, other mechanisms can also drive genetic differentiation. Habitat discontinuities, local adaptation, behaviour and dispersal capabilities are key to explaining the genetic variability between and within populations9,10. In the sea, it is widely recognised that oceanic fronts and currents represent sharp discontinuities of physical and biochemical variables11 and constitute barriers to faunal dispersal, often limiting gene flow between marine populations12. These oceanic features can explain the genetic structure of benthic marine littoral taxa, particularly those which are sedentary or with low vagility, in which population units are distributed in habitat patches whose connectivity is mediated by species’ life history traits13,14.
Despite comprehensive evidence for the influence of oceanic fronts and currents on the genetic structure of marine animals, their importance for organisms that are not strictly marine remains unclear, including the insects which dominate supralittoral rockpools in the terrestrial-marine ecotone. Beetles of the genus Ochthebius Leach, 1815 are the predominant macroinvertebrates inhabiting coastal supralittoral rockpools in many parts of the world, being particularly diverse in the western Palaearctic15,16. Although most adult Ochthebius can fly, their active dispersal capabilities may be limited by their small body size (c. 1–2 mm long) and erratic flight17. For this reason, similar to many small insects, they are likely to be subjected to passive dispersal by air currents or birds over short distances and by winds or even marine currents over medium to large spatial scale18,19. As previously proposed7, taxa such as Ochthebius may be affected by the action of storms and cyclones, which can promote gene flow among populations. These taxa lack a marine dispersal stage in their life cycle, but large waves or spring tides can remove them from supralittoral rockpools. Under these conditions, more resistant life-history stages such as eggs, able to survive at marine salinity conditions20,21, could be dispersed through coastal currents by floating or drifting on algal mats and plants/macroalgal remains, where they frequently oviposit. Marine currents may therefore be critical in shaping the diversity, connectivity and demography of inhabitants of supralittoral rockpools such as Ochthebius beetles, as reported in other arthropods such as Collembola22 and mites23.
On the western Mediterranean coast, the phylogeography of supralittoral Ochthebius has recently been studied7 using a combination of mitochondrial and nuclear genes. Villastrigo et al.7 proposed that both historical and contemporary marine barriers (Messinian Salinity Crisis and the Ibiza Channel, respectively) may have driven genetic isolation between extant populations. In particular, strong genetic discontinuities were observed in populations of two species (Ochthebius quadricollis Mulsant, 1844 and Ochthebius subinteger Mulsant & Rey, 1861) located on either side of the Ibiza Channel, which also acts as an impermeable geographical barrier for another (Ochthebius lejolisii Mulsant & Rey, 1861). Another genetic discontinuity was also found between western and eastern coastal areas of the Alboran Sea, where the interface between two anticyclonic gyres may operate as a barrier24. In a similar manner, Ochthebius populations from coastal rockpools in South Africa have apparently diverged in association with the Benguela and Agulhas marine currents19, like some fully marine members of the southern African rocky shore and estuarine communities25. Evidence suggests that properties of the ocean may exert a significant influence on the population biology and evolution of such taxa.
To investigate this further, and understand whether supralittoral rockpool inhabitants are primarily dispersed by oceanic currents, integrated approaches must include not only genetic data, but also biological and physical dispersal modelling (hereafter biophysical models). Such approaches have become common tools for the study of connectivity in both marine and terrestrial organisms26 due to the advantages they offer for estimating population connectivity over a range of spatial and temporal scales, and for multiple species at relatively low cost14. To date, they have been widely used in combination with genetic data to explore such questions in fully marine taxa27,28, but not the terrestrial-derived inhabitants of the coastal zone. In the Mediterranean Sea, they have also been used to assess levels of connectivity between protected and unprotected populations, evaluate dispersal potential from effective marine reserves, and hindcast the position of fish spawning areas29,30,31. Whilst biophysical models have proven to be relevant tools in the study of marine connectivity, they do need to be supported by other, complementary, techniques32. Combining oceanographic models with population genetic information32 is key to understanding how oceanographic currents and fronts may influence the extent of genetic variation within and among coastal populations. For example, strong links between genetic structure and biophysical model prediction would indicate that a significant part of the species’ phylogeographical structure results from ocean dynamics33.
This study is the first to combine population genetics and biophysical modelling to explore the possible role of oceanic currents and fronts in affecting the contemporary genetic structure of non-marine coastal organisms, focussing on supralittoral rockpool beetles in the Western Mediterranean Basin. Using this combined approach, we investigate whether the genetic structure of the studied species is best explained by a biophysical model or isolation by distance. We simulate potential population connectivity via marine propagules using a Lagrangian approach, and test whether this, or a model of isolation by distance best fits phylogeographic patterns in coastal Ochthebius.
Results
Simulated population connectivity
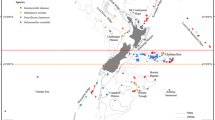
To investigate the potential dispersal of Ochthebius by oceanic currents along the western Mediterranean coast, propagule drifting was simulated for 21 rocky shore localities (Fig. 1) using Lagrangian tools. Of 3,528,000 released propagules, the mean connection probability was 1.88% (i.e., only 66.221 propagules), of which more than half were returned to the same locality by surface currents (Fig. S1) and approximately 40% successfully arriving at other sites. Locations in Catalonia (#1 to #3: Cala Sant Francesc, Vilanova and Sant Carles de la Ràpita) and Morocco (#21: Ghansou) did not have any connection to other locations. Additional localities without an effective connectivity (bellow 0.1% of their total propagules dispersed to other localities) were in the Gymnesian Islands (i.e., Mallorca and Menorca, localities #5 to #7). Two isolated clusters were detected across remaining sites separated by the Almeria-Oran Front: populations in the Alboran Sea (#18 to #20: Cala Rijana, Velilla and Nerja) and the remaining populations (#4 and #8 to #17). The most frequent connections were detected between adjacent populations (Fig. 2), with some localities behaving as source populations (e.g., Ibiza or Denia, locations #8 and #9, respectively), others as sinks (e.g., Cala Reona - location #14, with inputs from most localities except those from the Alboran Sea and Catalonia).

Orange circles indicate localities in which Ochthebius quadricollis and Ochthebius lejolisii were found in sympatry whilst blue diamonds represent localities exclusively for Ochthebius quadricollis. Arrows indicate the main direction of the surface oceanic currents. Main marine geographic barriers are as follows: Gibraltar Strait (GS), Almeria-Oran Front (AOF), Ibiza Channel (IC) and Balearic Front (BF).

It does not show connectivity below 0.1% of the total liberated propagules reaching another location, ignoring autoconnection. Each ribbon represents the connectivity between two populations: flow direction is indicated from source population (ribbon touching the scale) to sink population (blank space between ribbon and the scale).
Most localities showed propagule dispersal patterns from north to south, with very limited dispersal even to geographically close localities to their north. For example, propagules liberated on Ibiza disperse to the south, distant populations, such as Cala Reona, more than 200 km away, but displaying very low connectivity with a locality situated north of the Ibiza Channel. Populations located in the northern Ibiza Channel display negligible connectivity with the Balearic Islands, in most cases lower than 0.05% of realised propagules (scenario maximum of 75 propagules reaching Menorca [location #6], out of the 168.000 total propagules liberated in Vilanova i la Geltrú [location #2]). These results demonstrate the impact of the Ibiza Channel and the Balearic Front on dispersal from localities on the northern coast of the western Mediterranean.
Southern populations located close to the eastern boundary of the Almeria-Oran Front (locations #16 and #17) disperse principally from west to east, converging both west to east and north to south dispersal patterns in the southeastern Iberian Peninsula (Cala Reona - location #14).
Isolation by distance and Mantel correlations
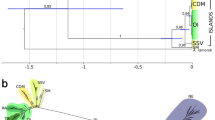
A significant positive relationship between geographical distance and genetic differentiation was seen only for COI in O. quadricollis (Fig. 3). There was no evidence that isolation by distance had a strong influence on genetic structure for either genes in O. lejolisii or for wingless in O. quadricollis (Fig. 3).

Ochthebius quadricollis (A, B) and Ochthebius lejolisii (B, C) for COI (A, C) and wingless (B, D) genes.
Mantel tests revealed that the biophysical model better explained genetic differentiation between populations than geographical distance regardless COI or wingless gene in O. quadricollis (Fig. 4 and Table S1). Mantel R statistics show a consistently higher correlation between the number of propagules that reach a locality (in the biophysical model) and genetic structure than with geographical distance, including 95% confidence intervals. Weaker evidence was found in O. lejolisii, however, where a significant correlation between biophysical model output and population differentiation was only seen in COI, with a similar Mantel R statistic confidence interval (Fig. 4 and Table S1).

Statistical significance (assessed by two-tailed p value after 9999 permutations) is indicated by asterisks.
Discussion
This study explores, for the first time, whether marine currents and their associated fronts influence the genetic structure of non-marine coastal taxa by affecting population dispersal. We show that a biophysical simulation of inter-population connectivity explains the genetic structure of supratidal Ochthebius beetles better than the classical isolation-by-distance model, suggesting that surface water currents may play an important role in Ochthebius’ dispersal and connectivity/isolation. Whilst some have suggested that such marine movement may occur in coastal beetles34, ours is the first study to integrate population genetics with biophysical modelling to show that at least some coastal insects may primarily disperse in a manner similar to that seen in truly marine organisms. The requirement of a dry surface that allow unfolding and drying of wings during metamorphosis is clearly very rare in the ocean, and indeed being a winged insects has been suggested to be one of the primary reasons why insects are largely absent from marine systems35. Our results strongly suggest that although most insects, including aquatic taxa, disperse mostly through active flight18, rockpool beetles move between distant localities mainly in a passive manner, via ocean currents, like the majority of fully marine inhabitants of the coastal zone.
Previous studies of passive dispersal via oceanic currents have focused on long-distance dispersal by floating or rafting in brooding intertidal invertebrates lacking a planktonic/pelagic dispersal stage36,37, or on relatively rare events allowing terrestrial taxa to cross oceans on, for example, drifting debris38,39,40,41. Whilst often proposing plausible dispersal scenarios, such studies have typically been based on limited data and relied on single methodologies, our study being the first to combine population genetics with biophysical modelling. Clearly a wider application of our approach to a range of biogeographical and evolutionary scenarios may prove illuminating.
The alternative hypothesis, isolation-by-distance explaining the current genetic structure of coastal Ochthebius, was rejected in almost all cases due to the lack of significant relationships between genetic and geographical distance, suggesting that additional factors played a more important role in the dispersal and isolation of Ochthebius populations at the spatial scale studied here. Our results suggested significant isolation-by-distance only for COI in O. quadricollis, something which was not mirrored by wingless in this species. Despite their widespread use, mitochondrial DNA data have serious limitations for detecting isolation-by-distance, being highly susceptible to type I error (i.e., false positives) when combining distinct regional populations and not very suitable for examining fine-scale geographical patterns on their own42. In our study, connectivity via oceanic currents better explains present genetic structure than geographical distance in O. quadricollis based on Mantel tests. Interestingly, the Mantel test showed no correlation between the biophysical model and the genetic structure of the wingless gene of O. lejolisii, whereas geographical distance was apparently correlated. This may reflect the different effects that oceanic currents have on the two species. As stated above, these two beetles are broadly sympatric, but their microhabitat preferences are slightly different: O. lejolisii is usually found in pools further from the sea16, potentially meaning that oceanic currents have a lesser impact on their population connectivity than in O. quadricollis. These interspecific differences in habitat preference, together with differences in physiological tolerance and life cycles20,43 may explain the apparently different effects of oceanic currents on the two species. However, O. lejolisii also has a more limited distribution on the western Mediterranean coast, reaching but not crossing the Ibiza Channel. Since the effect of the Ibiza Channel is therefore not tested in our models for this species (although the effect can be seen by its absence north of the Ibiza Channel), it may be that connectivity through oceanic currents is less apparent in our statistical tests.
Of course, it is possible that other dispersal mechanisms also contribute, but likely to a lesser extent, to the genetic structure observed in these Ochthebius. Anemochory may contribute to the dispersal of adult Ochthebius, although such coastal wind mediate dispersal would be expected to act at smaller spatial scales than ocean currents. Similarly, active flight, or flight assisted by winds likely plays a limited role in the long-distance dispersal of Ochthebius. As observed by other authors44, in hypersaline inland Ochthebius, genetic flow amongst populations is restricted by orographic factors, even over short spatial scales in the same watershed, suggesting limited capacity for long-distance dispersal by flight. Coastal Ochthebius may also disperse via the transport of eggs by endo- and epizoochory with seabirds that frequently occur in rockpools. No cases of Ochthebius transport through avifauna have been documented, although microcrustaceans are routinely found in bird plumage45,46,47, suggesting that such mechanisms may facilitate movement of rockpool invertebrates.
Our Lagrangian model strongly suggests that the Ibiza Channel and Balearic Front act as significant dispersal barriers in Ochthebius, something previously suggested based on molecular data alone7. Our results confirm the importance of oceanographic discontinuities as barriers to connectivity and gene flow along the Mediterranean coast9,12,48, even for non-marine taxa. We also found that populations of Ochthebius in the Alboran Sea are completely isolated from other Mediterranean populations, both in Spain and Morocco. Whereas Villastrigo et al.7 inferred the influence of a historical barrier within the Alboran Sea49, simulated population connection in the biophysical model suggests that the Almeria-Oran Front and associated processes act as a significant contemporary barrier to dispersal in rockpool Ochthebius. The Atlantic jet flowing through the Gibraltar Strait forms two anticyclonic gyres in the Alboran Sea50, which may facilitate dispersal of Ochthebius propagules from the western to the eastern Alboran basins, and prevent connectivity in the opposite direction24. In addition, the Atlantic jet hinders transport between Moroccan and Iberian coasts in the Alboran Sea.
In short, our study suggests that oceanic currents may play a significant role in the dispersal of non-marine coastal taxa, despite their lack of a specific planktonic/pelagic dispersal stage51. By combining biophysical dispersal modelling and population genetics we show that rockpool beetles apparently disperse in a similar manner to fully marine organisms, the first time this has been demonstrated for any non-marine insect. Further work, using a wider range of molecular markers (e.g., microsatellites or SNPs), and including additional taxa, will allow us to determine whether this applies to other areas and other coastal terrestrially-derived inhabitants.
Methods
Study species
Our study focused on two widespread representative species of rockpool specialist Ochthebius lineages: O. (Ochthebius) quadricollis and O. (Cobalius) lejolisii, which are partially sympatric in the Spanish Mediterranean coast (Fig. 1)16. O. quadricollis has a wide geographical distribution in the central and western Mediterranean plus the Atlantic coast of Morocco and the Iberian Peninsula7,16. O. lejolisii is distributed principally on the Atlantic coast from the British Isles to Morocco, reaching the southeastern Iberian Mediterranean coast16. The study area (see below) includes two additional coastal Ochthebius that were not considered in this study: Ochthebius subinteger more common in the central Mediterranean and with a low representation in the Iberian Peninsula16, and Ochthebius evae, a poorly known flightless species that likely occupy a different microhabitat52.
Although species living in this fluctuating and stressful environment are expected to have similar adaptations and life history traits, some studies21,43 have found differences in physiological tolerances related to ancestral and recent environmental variation in their habitats, being adult and larval stages of O. quadricollis more tolerant to heat than O. lejolisii, but the latter is more resistant to salt and desiccation52. These different traits are likely the result of their microhabitat preference, more distant from the sea in the case of O. lejolisii, and nearer to the coastline, with a higher incidence of sea spray and splash, in O. quadricollis, for which longer hydroperiod is required53. Both species are multivoltine with overlapping cohorts to cope with intense and fluctuating environmental stressors (e.g., salinity, temperature, dissolved oxygen or desiccation) and highly variable water availability across the year20. In addition, egg hatching is more successful in O. quadricollis than O. lejolisii, which, together with its shorter overall life cycle, leads to a greater demographic success of O. quadricollis20,21. Furthermore, Ochthebius adults, larvae and eggs are able to survive in seawater, eggs are even resistant to extreme desiccation for several days20,21.
Study area and field sampling
We focused on the western Mediterranean Sea, a complex marine system influenced by the inflow of lighter Atlantic water through the Gibraltar Strait that generates discontinuities when interacting with higher-density Mediterranean water (Fig. 1)11. In this area, there are several geographical barriers to the dispersal of marine organisms, including the Almeria-Oran Front, which has been suggested to play a key role in the genetic differentiation of species distributed both in the Atlantic Ocean and in the Mediterranean Sea48,54. Additional discontinuities that have been reported as strong barriers to gene exchange are the Balearic Front and the Ibiza Channel12,55.
We selected 20 rocky shore localities along the Mediterranean coast of the Iberian Peninsula and one in Morocco – from Cala Sant Francesc to Nerja plus Ghasou, representing all coastal features, including capes and gulfs. These localities were chosen non-randomly with the aim of equidistant localities but following the sites presented in Villastrigo et al.7 (Fig. 1). Individuals were collected from random pools following a gradient of distance from the sea in an attempt to collect both species at the localities where they are present.
Molecular data
We used the molecular dataset from Villastrigo et al.7 which contains samples of both target species along the Mediterranean coast of the Iberian Peninsula, excluding samples from contiguous regions (e.g., France or the Atlantic coast of the Iberian Peninsula and Morocco). This dataset contains molecular data for three randomly selected specimens per locality for two molecular markers: 1) the 3′-end of the mitochondrial COI gene (829 bp) and 2) a fragment of the nuclear gene wingless (466 bp). Sample information including geographical data and accession numbers for DNA sequences is available in Table S2 and Fig. 1.
To reduce the molecular uncertainty caused by DNA ambiguities, we inferred haplotype sequences using the PHASE software56 as implemented in DNAsp 657, considering 1,000 iterations, a thinning parameter of 5 and discarding the first 100 iterations as burn-in. We then calculated the fixation index (Fst) as a measure of genetic differentiation between pairs of localities using DNAsp 657. Linearization of Fst values was done following the regression of Fst/(1-Fst) by Rousset58 in the R environment59 to estimate the isolation-by-distance effect.
Biophysical model
To investigate the potential dispersal of Ochthebius through oceanic currents in western Mediterranean and therefore population connectivity, propagule drifting was simulated using the Lagrangian software Ichthyop v3.360. This creates an individual-based, three-dimensional particle-tracking model that integrates the input time series of salinity, temperature and current velocity fields generated by the ocean model to advect and influence the virtual propagules. In this study, the term “propagule” is used to refer to resistant Ochthebius eggs that can be transported by oceanic currents without affecting their survival (the low floating capability of adults and larvae and the aerial respiration of adults makes them less likely to disperse via oceanic currents).
A multi-year simulation of the Western Mediterranean OPerational modelling system (WMOP)61,62, developed by the Balearic Islands Coastal Observing and Forecasting System63 was used to simulate propagule trajectories and velocity. WMOP is based on the regional configuration of the Regional Oceanic Modeling System ROMS model64 implemented over the western Mediterranean Sea, which is a free-surface, split-explicit model solving primitive hydrostatic equations using terrain-following curvilinear vertical coordinates, with a horizontal resolution of ca. 2 km and 32 sigma-levels in the vertical dimension. It covers an area from the Gibraltar Strait to the Sardinia Channel (6°W to 9.2°E and 35°N to 44.5°N). The model is forced by high-resolution atmospheric forcing (5 km, 3 h) from the High-Resolution Limited Area Model (HIRLAM) simulations developed by the Spanish Meteorological Agency. The simulation used in this study is a free-run hindcast simulation of WMOP, spanning the period 2009-2015. Initial and boundary conditions were provided by the CMEMS Mediterranean model reanalysis65. Further details, evaluation and validation of this high-resolution WMOP simulation were presented by Mourre et al.62 and Aguiar et al.66. In particular, the simulation was shown to accurately represent the main circulation features (including the Atlantic Jet, Alboran gyres and the Algerian, Northern and Balearic Currents), as well as the spatial variability of surface eddy kinetic energy.
The dispersal model considered the release of 1000 particles representing virtual Ochthebius propagules from each study site. For this purpose, the release zone is defined by the geographical coordinates of a 50 metres radius polygon centred 100 m offshore and extending from 0 to 1 m depth. Particle releases were scheduled every 15 days from January to December throughout the study period (2009-2015) to cover several consecutive reproductive seasons and record the annual and interannual variability in marine currents. In total, 3,528,000 propagules were released (1000 propagules × 7 years × 12 months × 2 releases per month × 21 localities). After release, the location of each propagule was tracked for 5 days considering the at-sea duration of viable propagules, based on the expected hatching time20. This model was chosen to produce the most realistic but conservative dispersal and connectivity scenario, whilst keeping the computational cost to a manageable level.
Ichthyop output files gathered all possible propagule locations at the end of the marine dispersal period, which were analysed using geographic information systems. Twenty-one rockpool recruitment areas were then considered, from Cala Sant Francesc in the north (Catalonia, Spain) to Ghansou (Morocco) in the south. Virtual propagules were considered to have reached a recruitment area when they reached a minimum buffer distance of approximately 10 km radius from the epicentre of any other studied rockpool locality. This buffer size was considered to cover a reasonable water area surrounding the site, but to avoid overlapping the buffer zones of closely spaced sites. Raw values of pairwise potential connectivity were calculated and visualised using Circos67. The spread of propagules throughout our biophysical model does not take into account their survival during the dispersal event (e.g., predation by other organisms) and/or their establishment on new rocky shores. For this purpose, we considered two localities to be positively connected when the number of propagules reaching a locality was equal to or greater than 0.1% of the initial number of propagules dispersed from the source locality.
Data analyses
The relationship between genetic differentiation and geographical distance between studied localities was explored using isolation by distance (IBD) analysis. A linear model (lm function of the R-package stats59) was used to fit linearised Fst values against the natural logarithm of geographical distance between populations. Linear geographical distances between pairs of locations were computed using the vector analysis tool in QGIS 3.16.13.
To assess whether the simulated oceanic connection or IBD best fits observed genetic patterns, Euclidean distance matrices for the following variables were generated using the dist function of the R-package stats59: Fst values, geographical distance and the number of simulated propagules dispersed between each pair of localities (raw data available in Table S3). Due to the lack of genetic data for multiple specimens, we discarded six localities (numbers 2, 4, 6, 7, 8 and 21) in subsequent analyses. Mantel tests were calculated for the Fst distance matrix against the distance matrices of the other variables using the Spearman correlation method under the mantel function of the R-package ecodist68 with 9,999 permutations. Bootstrapped confidence intervals (95%) for the Mantel test were calculated using 9999 iterations.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The data is fully available in the Supplementary material.
References
Parvizi, E., Fraser, C. I. & Waters, J. M. Genetic impacts of physical disturbance processes in coastal marine ecosystems. J. Biogeogr. 49, 1877–1890 (2022).
Thomsen, M. S. et al. Local extinction of bull kelp (Durvillaea spp.) due to a marine heatwave. Front. Mar. Sci. 6, 84 (2019).
Thorner, J., Kumar, L. & Smith, S. D. A. Impacts of climate-change-driven sea level rise on intertidal rocky reef habitats will be variable and site specific. PLoS ONE 9, e86130 (2014).
Greenan, T. M., Griffiths, C. L. & Santamaria, C. A. Molecular approaches uncover cryptic diversity in intertidal Ligia isopods (Crustacea, Isopoda, Ligiidae) across the southern Africa coastline. PeerJ 6, e4658 (2018).
Pfingstl, T., Lienhard, A., Baumann, J. & Koblmüller, S. A taxonomist’s nightmare - Cryptic diversity in Caribbean intertidal arthropods (Arachnida, Acari, Oribatida). Mol. Phylogenet. Evol. 163, 107240 (2021).
Sabatelli, S., Ruspantini, P., Cardoli, P. & Audisio, P. Underestimated diversity: Cryptic species and phylogenetic relationships in the subgenus Cobalius (Coleoptera: Hydraenidae) from marine rockpools. Mol. Phylogenet. Evol. 163, 107243 (2021).
Villastrigo, A. et al. Cryptic lineages, cryptic barriers: historical seascapes and oceanic fronts drive genetic diversity in supralittoral rockpool beetles (Coleoptera: Hydraenidae). Zool. J. Linn. Soc. 196, 740–756 (2022).
Margalef, R. Sobre la ecología de las larvas del mosquito Aëdes mariae. Publ. Inst. Biol. Apl. 6, 83–101 (1949).
Schunter, C. et al. Matching genetics with oceanography: directional gene flow in a Mediterranean fish species. Mol. Ecol. 20, 5167–5181 (2011).
Pascual, M., Rives, B., Schunter, C. & MaCpherson, E. Impact of life history traits on gene flow: a multispecies systematic review across oceanographic barriers in the Mediterranean Sea. PLoS ONE 12, 1–20 (2017).
Millot, C. Circulation in the Mediterranean Sea: evidences, debates and unanswered questions. scimar 69, 5–21 (2005).
Galarza, J. A. et al. The influence of oceanographic fronts and early-life-history traits on connectivity among littoral fish species. Proc. Natl. Acad. Sci. USA. 106, 1473–1478 (2009).
Cowen, R. K., Lwiza, K. M., Sponaugle, S., Paris, C. B. & Olson, D. B. Connectivity of marine populations: open or closed? Science 287, 857–859 (2000).
Cowen, R. K. & Sponaugle, S. Larval dispersal and marine population connectivity. Ann. Rev. Mar. Sci. 1, 443–466 (2009).
Sabatelli, S. et al. Molecular ecology and phylogenetics of the water beetle genus Ochthebius revealed multiple independent shifts to marine rockpools lifestyle. Zool. Scr. 45, 175–186 (2016).
Villastrigo, A., Hernando, C. & Millán, A. The Ochthebius (Coleoptera, Hydraenidae) from western Palaearctic supratidal rockpools. Supl. del Bol. Asoc. Esp. Entomol. 4, 100–108 (2022).
Urbanelli, S. Genetic divergence and reproductive isolation in the Ochthebius (Calobius) complex (Coleoptera: Hydraenidae). Heredity 88, 333–341 (2002).
Bilton, D. T., Freeland, J. R. & Okamura, B. Dispersal in freshwater invertebrates. Annu. Rev. Ecol. Syst. 32, 159–181 (2001).
Bilton, D. T. Differentiation of South African coastal rock pool Ochthebius is associated with major ocean currents (Coleoptera: Hydraenidae). Acta Entomol. Mus. Natl. Pragae 61, 253–260 (2021).
Velasco, J., Mirón-Gatón, J. M., García-Meseguer, A. J. & Botella-Cruz, M. Life cycle differences between two coexisting species of supratidal rockpools: Ochthebius quadricollis Mulsant, 1844 and Ochthebius lejolisii Mulsant and Rey, 1861 (coleoptera, Hydraenidae). Supl. Bol. Asoc. Esp. Entomol. 4, 131–136 (2022).
Mirón-Gatón, J. M., Botella-Cruz, M., García-Meseguer, J. A., Millán, A. & Velasco, J. Discordant pattern between realised and fundamental saline niches in two supralittoral Ochthebius species (Coleoptera: Hydraenidae). Ecol. Entomol. 48, 284–294 (2023)
McGaughran, A., Stevens, M. I. & Holland, B. R. Biogeography of circum-Antarctic springtails. Mol. Phylogenet. Evol. 57, 48–58 (2010).
Pfingstl, T. Resistance to fresh and salt water in intertidal mites (Acari: Oribatida): implications for ecology and hydrochorous dispersal. Exp. Appl. Acarol. 61, 87–96 (2013).
Sánchez-Garrido, J. C. & Nadal, I. The Alboran Sea circulation and its biological response: a review. Front. Mar. Sci. 9, 933390 (2022).
von der Heyden, S. Why do we need to integrate population genetics into South African marine protected area planning? Afr. J. Mar. Sci. 31, 263–269 (2009).
Lett, C., Barrier, N. & Bahlali, M. Converging approaches for modeling the dispersal of propagules in air and sea. Ecol. Modell. 415, 108858 (2020).
Monzón-Argüello, C. et al. Evidence from genetic and Lagrangian drifter data for transatlantic transport of small juvenile green turtles. J. Biogeogr. 37, 1752–1766 (2010).
Freire, A. S. et al. Does the transport of larvae throughout the south Atlantic support the genetic and morphometric diversity of the Sally Lightfoot Crabs Grapsus grapsus (Linnaeus, 1758) and Grapsus adscensionis (Osbeck, 1765) (Decapoda: Grapsidae) among the oceanic islands? J. Mar. Syst. 223, 103614 (2021).
Andrello, M. et al. Low connectivity between Mediterranean marine protected areas: a biophysical modeling approach for the dusky grouper Epinephelus marginatus. PLoS ONE 8, e68564 (2013).
Calò, A. et al. A review of methods to assess connectivity and dispersal between fish populations in the Mediterranean Sea. Adv. Oceanogr. Limnol. 4, 150–175 (2013).
Legrand, T., Di Franco, A., Ser-Giacomi, E., Caló, A. & Rossi, V. A multidisciplinary analytical framework to delineate spawning areas and quantify larval dispersal in coastal fish. Mar. Environ. Res. 151, 104761 (2019).
Bode, M. et al. Successful validation of a larval dispersal model using genetic parentage data. PLoS Biol. 17, e3000380 (2019).
Jahnke, M. & Jonsson, P. R. Biophysical models of dispersal contribute to seascape genetic analyses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 377, 20210024 (2022).
Nakahama, N. et al. Possible dispersal of the coastal and subterranean carabid beetle Thalassoduvalius masidai (Coleoptera) by ocean currents. Biol. J. Linn. Soc. 135, 265–276 (2022).
Ruxton, G. D. & Humphries, S. Can ecological and evolutionary arguments solve the riddle of the missing marine insects? Mar. Ecol. 29, 72–75 (2008).
Highsmith, R. C. Floating and algal rafting as potential dispersal mechanisms in brooding invertebrates. Mar. Ecol. Prog. Ser. 25, 169–179 (1985).
Reid, D. G. Systematics and Evolution of Littorina. vol. 164 463 (The Ray Society, 1996).
Houle, A. Floating islands: a mode of long-distance dispersal for small and medium- sized terrestrial vertebrates. Divers. Distrib. 4, 201–216 (1998).
de Queiroz, A. The resurrection of oceanic dispersal in historical biogeography. Trends Ecol. Evol. 20, 68–73 (2005).
Yeh, H.-Y. et al. Rafting on floating fruit is effective for oceanic dispersal of flightless weevils. J. Exp. Biol. 221, jeb190488 (2018).
Lindo, Z. Transoceanic dispersal of terrestrial species by debris rafting. Ecography 43, 1364–1372 (2020).
Teske, P. R. et al. Mitochondrial DNA is unsuitable to test for isolation by distance. Sci. Rep. 8, 8448 (2018).
Mirón-Gatón, J. M., Botella-Cruz, M., García-Meseguer, A. J., Millán, A. & Velasco, J. Thermal tolerance differs between co-occurring congeneric beetle species in marine supratidal rockpools. Mar. Ecol. Prog. Ser. 681, 185–196 (2022).
Abellán, P. et al. Conservation genetics in hypersaline inland waters: Mitochondrial diversity and phylogeography of an endangered Iberian beetle (Coleoptera: Hydraenidae). Conserv. Genet. 8, 79–88 (2007).
Incagnone, G., Marrone, F., Barone, R., Robba, L. & Naselli-Flores, L. How do freshwater organisms cross the “dry ocean”? A review on passive dispersal and colonization processes with a special focus on temporary ponds. Hydrobiologia 750, 103–123 (2015).
Powlik, J. J. Habitat characters of Tigriopus californicus (Copepoda: Harpacticoida), with notes on the dispersal of supralittoral fauna. J. Mar. Biol. Assoc. UK 79, 85–92 (1999).
Swanson, G. A. Dissemination of Amphipods by Waterfowl. J. Wildl. Manage. 48, 988–991 (1984).
García-Merchán, V. H. et al. Phylogeographic patterns of decapod crustaceans at the Atlantic–Mediterranean transition. Mol. Phylogenet. Evol. 62, 664–672 (2012).
Booth-Rea, G., R. Ranero, C. & Grevemeyer, I. The Alboran volcanic-arc modulated the Messinian faunal exchange and salinity crisis. Sci. Rep. 8, 1–14 (2018).
Millot, C. Circulation in the Western Mediterranean Sea. J. Mar. Syst. 20, 423–442 (1999).
Kyle, C. J. & Boulding, E. G. Comparative population genetic structure of marine gastropods (Littorina spp.) with and without pelagic larval dispersal. Mar. Biol. 137, 835–845 (2000).
Villastrigo, A., Hernando, C., Millán, A. & Ribera, I. The neglected diversity of the Ochthebius fauna from Eastern Atlantic and Central and Western Mediterranean coastal rockpools (Coleoptera, Hydraenidae). Org. Divers. Evol. 20, 785–801 (2020).
García-Meseguer, A. J. et al. Fine-scale niche differences allow the co-existence of congeneric aquatic beetles in supratidal rockpools. Hydrobiologia. https://doi.org/10.1007/s10750-023-05333-0 (2023).
Patarnello, T., Volckaert, F. A. M. J. & Castilho, R. Pillars of Hercules: is the Atlantic-Mediterranean transition a phylogeographical break? Mol. Ecol. 16, 4426–4444 (2007).
Torrado, H. et al. Impact of individual early life traits in larval dispersal: A multispecies approach using backtracking models. Prog. Oceanogr. 192, 102518 (2021).
Stephens, M., Smith, N. J. & Donnelly, P. A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Genet. 68, 978–989 (2001).
Rozas, J. et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol 34, 3299–3302 (2017).
Rousset, F. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genetics 145, 1219–1228 (1997).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Preprint at https://www.R-project.org/ (2022).
Lett, C. et al. A Lagrangian tool for modelling ichthyoplankton dynamics. Environ. Model. Softw. 23, 1210–1214 (2008).
Juza, M. et al. SOCIB operational ocean forecasting system and multi-platform validation in the Western Mediterranean Sea. J. Operational Oceanogr. 9, s155–s166 (2016).
Mourre, B. et al. Assessment of high-resolution regional ocean prediction systems using multi-platform observations: Illustrations in the western Mediterranean Sea. in New Frotiers in Operation Oceanography (eds. Chassignet, E. P., Pascual, A., Tintoré, J. & Verron, J.) 663–694 (2018).
Tintoré, J. et al. SOCIB: the balearic islands coastal ocean observing and forecasting system responding to science, technology and society needs. Mar. Technol. Soc. J. 47, 101–117 (2013).
Shchepetkin, A. F. & McWilliams, J. C. The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Model 9, 347–404 (2005).
Simoncelli, S. et al. Mediterranean Sea Physical Reanalysis (CMEMS MED-Physics) (Version 1) [Data set]. Copernicus Monitoring Environment Marine Service (CMEMS). Preprint at https://doi.org/10.25423/MEDSEA_REANALYSIS_PHYS_006_004 (2019).
Aguiar, E. et al. Multi-platform model assessment in the Western Mediterranean Sea: impact of downscaling on the surface circulation and mesoscale activity. Ocean Dyn. 70, 273–288 (2020).
Krzywinski, M. et al. Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645 (2009).
Goslee, S. C. & Urban, D. L. The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 22, 1–19 (2007).
Acknowledgements
This work was partly funded by the projects CGL2017-84157-P funded by MCIN/AEI/10.13039/501100011033 and by ‘ERDF A way of making Europe’ to J.V.; A.V. was funded by the Alexander von Humboldt Foundation through a Humboldt Research Fellowship and by the Carl Friedrich von Siemens Foundation. A.J.G.M. was supported by a predoctoral grant (FPI programme) and J.M.M.G. by a predoctoral grant (FPU programme). We are indebted to Ignacio Ribera for his undoubted contribution to the study of coastal rockpool fauna in the western Palaearctic, which was crucial to our current understanding and drove our curiosity into these usually neglected species.
Author information
Authors and Affiliations
Contributions
A.V.: methodology, data analyses, interpretation, writing - original draft, writing - review and editing. V.O.-S.: methodology, data analyses, interpretation, writing - original draft, writing - review and editing. A.J.G.-M.: interpretation, writing - review and editing. J.M.M.-G.: interpretation, writing - review and editing. B.M.: writing - review and editing. A.M.: conceptualisation, interpretation, writing - original draft and writing - review and editing. J.V.: conceptualisation, interpretation, writing - original draft, writing - review and editing, project administration and funding acquisition. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Villastrigo, A., Orenes-Salazar, V., García-Meseguer, A.J. et al. Oceanic currents maintain the genetic structure of non-marine coastal taxa in the western Mediterranean Sea. npj biodivers 2, 25 (2023). https://doi.org/10.1038/s44185-023-00028-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s44185-023-00028-0
