
Abstract
Gonadotropin-releasing hormone (GnRH) has a role in hypothalamic control of aging, but the underlying patterns and relationship with downstream reproductive hormones are still unclear. Here we report that hypothalamic GnRH pulse frequency and irregularity increase before GnRH pulse amplitude slowly decreases during aging. GnRH is inhibited by nuclear factor (NF)-κB, and GnRH pulses were controlled by oscillations in the transcriptional activity of NF-κB. Exposure to testosterone under pro-inflammatory conditions stimulated both NF-κB oscillations and GnRH pulses. While castration of middle-aged mice induced short-term anti-aging effects, preventing elevation of luteinizing hormone (LH) levels after castration led to long-term anti-aging effects and lifespan extension, indicating that high-frequency GnRH pulses and high-magnitude LH levels coordinately mediate aging. Reprogramming the endogenous GnRH pulses of middle-aged male mice via an optogenetic approach revealed that increasing GnRH pulses frequency causes LH excess and aging acceleration, while lowering the frequency of and stabilizing GnRH pulses can slow down aging. In conclusion, GnRH pulses are important for aging in male mice.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
Data availability
Source data are provided with this paper. There is no restriction on data availability for this manuscript.
Change history
20 October 2021
A Correction to this paper has been published: https://doi.org/10.1038/s43587-021-00137-0
References
Zhang, G. et al. Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nature 497, 211–216 (2013).
Cai, D. & Khor, S. “Hypothalamic microinflammation” paradigm in aging and metabolic diseases. Cell Metab. 30, 19–35 (2019).
Zhang, Y. L. et al. Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature 548, 52–57 (2017).
Hsin, H. & Kenyon, C. Signals from the reproductive system regulate the lifespan of C. elegans. Nature 399, 362–366 (1999).
Hamilton, J. B. & Mestler, G. E. Mortality and survival: comparison of eunuchs with intact men and women in a mentally retarded population. J. Gerontol. 24, 395–411 (1969).
Wilson, J. D. & Roehrborn, C. Long-term consequences of castration in men: lessons from the Skoptzy and the eunuchs of the Chinese and Ottoman courts. J. Clin. Endocrinol. Metab. 84, 4324–4331 (1999).
Min, K. J., Lee, C. K. & Park, H. N. The lifespan of Korean eunuchs. Curr. Biol. 22, R792–R793 (2012).
Veldhuis, J. D. et al. The aging male hypothalamic–pituitary–gonadal axis: pulsatility and feedback. Mol. Cell. Endocrinol. 299, 14–22 (2009).
Veldhuis, J. D. Aging and hormones of the hypothalamo–pituitary axis: gonadotropic axis in men and somatotropic axes in men and women. Ageing Res. Rev. 7, 189–208 (2008).
Tenover, J. S., Matsumoto, A. M., Plymate, S. R. & Bremner, W. J. The effects of aging in normal men on bioavailable testosterone and luteinizing hormone secretion: response to clomiphene citrate. J. Clin. Endocrinol. Metab. 65, 1118–1126 (1987).
Rosario, E. R., Carroll, J. C. & Pike, C. J. Evaluation of the effects of testosterone and luteinizing hormone on regulation of β-amyloid in male 3×Tg-AD mice. Brain Res. 1466, 137–145 (2012).
Hyde, Z. et al. Higher luteinizing hormone is associated with poor memory recall: the Health in Men Study. J. Alzheimers Dis. 19, 943–951 (2010).
Verdile, G. et al. Luteinizing hormone levels are positively correlated with plasma amyloid-β protein levels in elderly men. J. Alzheimers Dis. 14, 201–208 (2008).
Greenhill, C. Pituitary function: pulsatile GnRH therapy in CCPHD. Nat. Rev. Endocrinol. 13, 315 (2017).
Pincus, S. M., Veldhuis, J. D., Mulligan, T., Iranmanesh, A. & Evans, W. S. Effects of age on the irregularity of LH and FSH serum concentrations in women and men. Am. J. Physiol. 273, E989–E995 (1997).
Keenan, D. M. & Veldhuis, J. D. Disruption of the hypothalamic luteinizing hormone pulsing mechanism in aging men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 281, R1917–R1924 (2001).
Mulligan, T., Iranmanesh, A., Kerzner, R., Demers, L. W. & Veldhuis, J. D. Two-week pulsatile gonadotropin releasing hormone infusion unmasks dual (hypothalamic and Leydig cell) defects in the healthy aging male gonadotropic axis. Eur. J. Endocrinol. 141, 257–266 (1999).
Veldhuis, J. D., Urban, R. J., Lizarralde, G., Johnson, M. L. & Iranmanesh, A. Attenuation of luteinizing hormone secretory burst amplitude as a proximate basis for the hypoandrogenism of healthy aging in men. J. Clin. Endocrinol. Metab. 75, 707–713 (1992).
Choe, H. K. et al. Synchronous activation of gonadotropin-releasing hormone gene transcription and secretion by pulsatile kisspeptin stimulation. Proc. Natl Acad. Sci. USA 110, 5677–5682 (2013).
Adamson, A. et al. Signal transduction controls heterogeneous NF-κB dynamics and target gene expression through cytokine-specific refractory states. Nat. Commun. https://doi.org/10.1038/ncomms12057 (2016).
Ashall, L. et al. Pulsatile stimulation determines timing and specificity of NF-κB-dependent transcription. Science 324, 242–246 (2009).
Nelson, D. E. et al. Oscillations in NF-κB signaling control the dynamics of gene expression. Science 306, 704–708 (2004).
Tilbrook, A. J. & Clarke, I. J. Negative feedback regulation of the secretion and actions of gonadotropin-releasing hormone in males. Biol. Reprod. 64, 735–742 (2001).
Kelly, D. M. & Jones, T. H. Testosterone: a metabolic hormone in health and disease. J. Endocrinol. 217, R25–R45 (2013).
Holmes, S., Singh, M., Su, C. & Cunningham, R. L. Effects of oxidative stress and testosterone on pro-inflammatory signaling in a female rat dopaminergic neuronal cell line. Endocrinology 157, 2824–2835 (2016).
Bolshakova, A., Magnusson, K. E., Pinaev, G. & Petukhova, O. EGF-induced dynamics of NF-κB and F-actin in A431 cells spread on fibronectin. Histochem. Cell Biol. 1—44, 223–235 (2015).
Campos, P. & Herbison, A. E. Optogenetic activation of GnRH neurons reveals minimal requirements for pulsatile luteinizing hormone secretion. Proc. Natl Acad. Sci. USA 111, 18387–18392 (2014).
Yu, B., Tang, Y. & Cai, D. Brain is an endocrine organ through secretion and nuclear transfer of parathymosin. Life Sci. Alliance https://doi.org/10.26508/lsa.202000917 (2020).
Tang, Y., Zuniga-Hertz, J. P., Han, C., Yu, B. & Cai, D. Multifaceted secretion of htNSC-derived hypothalamic islets induces survival and antidiabetic effect via peripheral implantation in mice. eLife https://doi.org/10.7554/eLife.52580 (2020).
Li, J. X., Tang, Y. Z. & Cai, D. S. IKKβ/NF-κB disrupts adult hypothalamic neural stem cells to mediate a neurodegenerative mechanism of dietary obesity and pre-diabetes. Nat. Cell Biol. 14, 999–1012 (2012).
Meredith, J. M., Turek, F. W. & Levine, J. E. Effects of gonadotropin-releasing hormone pulse frequency modulation on the reproductive axis of photoinhibited male Siberian hamsters. Biol. Reprod. 59, 813–819 (1998).
Cai, D. et al. IKKβ/NF-κB activation causes severe muscle wasting in mice. Cell 119, 285–298 (2004).
Oakley, A. E., Clifton, D. K. & Steiner, R. A. Kisspeptin signaling in the brain. Endocr. Rev. 30, 713–743 (2009).
Okamura, H. et al. Kisspeptin and GnRH pulse generation. Adv. Exp. Med. Biol. 784, 297–323 (2013).
Herbison, A. E. The gonadotropin-releasing hormone pulse generator. Endocrinology 159, 3723–3736 (2018).
Simonneaux, V. A Kiss to drive rhythms in reproduction. Eur. J. Neurosci. 51, 509–530 (2020).
Titolo, D., Cai, F. & Belsham, D. D. Coordinate regulation of neuropeptide Y and agouti-related peptide gene expression by estrogen depends on the ratio of estrogen receptor (ER)α to ERβ in clonal hypothalamic neurons. Mol. Endocrinol. 20, 2080–2092 (2006).
Klenke, U., Constantin, S. & Wray, S. Neuropeptide Y directly inhibits neuronal activity in a subpopulation of gonadotropin-releasing hormone-1 neurons via Y1 receptors. Endocrinology 151, 2736–2746 (2010).
Verma, S., Kirigiti, M. A., Millar, R. P., Grove, K. L. & Smith, M. S. Endogenous kisspeptin tone is a critical excitatory component of spontaneous GnRH activity and the GnRH response to NPY and CART. Neuroendocrinology 99, 190–203 (2014).
Roa, J. & Herbison, A. E. Direct regulation of GnRH neuron excitability by arcuate nucleus POMC and NPY neuron neuropeptides in female mice. Endocrinology 153, 5587–5599 (2012).
Kim, K. & Choe, H. K. Role of hypothalamus in aging and its underlying cellular mechanisms. Mech. Ageing Dev. 177, 74–79 (2019).
Kermath, B. A., Riha, P. D., Woller, M. J., Wolfe, A. & Gore, A. C. Hypothalamic molecular changes underlying natural reproductive senescence in the female rat. Endocrinology 155, 3597–3609 (2014).
Zhang, X. et al. Hypothalamic IKKβ/NF-κB and ER stress link overnutrition to energy imbalance and obesity. Cell 135, 61–73 (2008).
Au, A. et al. Estrogens, inflammation and cognition. Front. Neuroendocrinol. 40, 87–100 (2016).
Zhang, Y., Reichel, J. M., Han, C., Zuniga-Hertz, J. P. & Cai, D. Astrocytic process plasticity and IKKβ/NF-κB in central control of blood glucose, blood pressure, and body weight. Cell Metab. 25, 1091–1102 (2017).
Sieben, C. J. et al. BubR1 allelic effects drive phenotypic heterogeneity in mosaic-variegated aneuploidy progeria syndrome. J. Clin. Invest. 130, 171–188 (2020).
Johnson, M. L. et al. AutoDecon: a robust numerical method for the quantification of pulsatile events. Methods Enzymol. 454, 367–404 (2009).
Pincus, S. M. & Goldberger, A. L. Physiological time-series analysis: what does regularity quantify? Am. J. Physiol. 266, H1643–H1656 (1994).
Fix, C., Jordan, C., Cano, P. & Walker, W. H. Testosterone activates mitogen-activated protein kinase and the cAMP response element binding protein transcription factor in Sertoli cells. Proc. Natl Acad. Sci. USA 101, 10919–10924 (2004).
Hatef, A. & Unniappan, S. Gonadotropin-releasing hormone, kisspeptin, and gonadal steroids directly modulate nucleobindin-2/nesfatin-1 in murine hypothalamic gonadotropin-releasing hormone neurons and gonadotropes. Biol. Reprod. 96, 635–651 (2017).
Acknowledgements
We thank the Cai laboratory members for technical support, A. Wolfe (Johns Hopkins University) for the Gnrh-Cre mouse model, P. Mellon (University of California, San Diego) for GT1-7 and LβT4 cells, and the IVIS facility at Einstein for in vivo imaging assistance. This study was supported directly or indirectly through Einstein resources and NIH grants R01 AG031774, DK121435 and HL147477 (all to D.C.).
Author information
Authors and Affiliations
Contributions
Z.W. co-designed and performed experiments, including GnRH pulse imaging, optogenetic models and physiological assays, castration model follow-up, cell culture models, immunostaining, histology and biochemistry, performed data analysis, contributed to interpretation of the data and prepared the figures for the paper. W.W. co-designed and performed experiments, including castration, LH knockdown, physiology and biochemistry experiments, and performed data analysis and interpretation. M.K. contributed to the castration model, GnRH knockdown and physiology studies. D.C. conceived the idea and hypothesis, conceptualized the project, designed the whole study and specific aims, constructed the experimental framework, supervised the study, led data analysis and interpretation, and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Aging thanks Cláudia Cavadas and Han Kyoung Choe for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Pulses of blood FSH concentrations in male mice of different ages.
Blood FSH concentrations were measured during 180 mins in young (3 months), early middle-aged (11–12 months) and late middle-aged (16–17 months) mice. a: Representative blood FSH concentrations. b–d: Quantitative analysis of pulse frequency (b), peak concentration (c), and AUC value (d) of blood FSH concentrations. Comparisons of all groups were performed with one-way ANOVA but without statistical significance, n = 5 mice per group (b–d). Bar graphs reflect mean ± s.e.m.
Extended Data Fig. 2 A schematic model of oscillated GnRH mRNA production and NF-κB signaling.
This schematic diagram through drawing illustrative curves (which were not experimental data) is to generalize the concept for the relationship among oscillations of NF-κB activation and the production of IκBα protein, IκBα mRNA and GnRH mRNA in GnRH cells. Based on this conceptual model, it can be deduced that when NF-κB is chronically up-activated during aging, it leads to an equilibrium of fast-frequency NF-κB nuclear oscillation and fast-frequency pulsatile GnRH mRNA production.
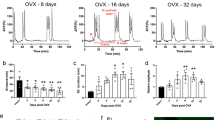
Extended Data Fig. 3 Late age physiology and lifespan of mice with castration at the age of 8 months.
(a) Schematic diagram of the experimental procedure. (b–i) Mice were assigned into subgroups for different behavioral assays at the age of 18–19 months for locomotion (b), treadmill (c), coordination (d), novel recognition (e), sociality (f), learning and memory (g), adhesive removal (h), and odor smelling (i). (j–k) Subgroups of mice were profiled for pulse frequencies (j) and peak concentrations (k) of blood LH. (l) Lifespan follow-up of these mice. P values are shown in figures for comparisons between groups as indicated (k, l) or labeled as non-significant (NS) (b–j), two-tailed Student’s t-test (b–k), log-rank (Mantel-Cox) test (l); n = 8 per group (b–i), n = 5 per group (j, k); n = 22 mice per group (l). Data are mean ± s.e.m.
Extended Data Fig. 5 GnRH-Cre in subpopulation of Sox2-positive hypothalamic cells.
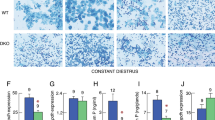
GnRH-Cre mice were crossed with Rosa 26-lox-STOP-lox-GFP mice leading to the offspring of GnRHCre:Rosa GFP mice, and the hypothalamic sections of these offspring were made for co-immunostaining of Sox2 and GnRH-Cre-dependent GFP expression (labelled as GnRH-GFP). White arrows point to representative cells which are co-positive for Sox2 and GFP. Images represent 4 independent repeats per condition. Scare bar: 100 μm.
Extended Data Fig. 6 Fast optogenetic stimulation prevents NF-κB inhibition from slowing LH pluses.
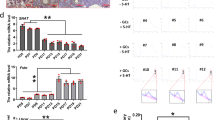
GnRH-Cre mice (12-month-old male) received hypothalamic injection of AAV together with a lentivirus containing GnRH promoter-driven DN-IκBα (labelled as DNIκBα) vs. control (labeled as Control), and subsequently these animals were treated with fast optogenetic stimulation vs. sham procedure as described in Fig. 5 and profiled for blood concentrations of LH (a–d) and FSH (e–h). (a, e) Representative blood LH (a) and FSH (e) concentrations. (b–d, f–h) Quantitative analysis of pulse frequency (b, f), peak concentration (c, g) and AUC value (d, h) of blood LH (b-d) and FSH (f–h). P values are shown in figures for comparisons between groups as indicated, one-way ANOVA with Tukey’s post hoc test (b–d), n = 5 mice per group (b–d). Comparisons of all groups with one-way ANOVA for lacking statistical significance (f–h), n = 5 mice per group (f–h). Data are mean ± s.e.m.
Extended Data Fig. 7 Fast optogenetic stimulation reduces the anti-aging effect of NF-κB inhibition.
GnRH-Cre male mice (12 months old) received hypothalamic injection of AAV together with a lentivirus containing GnRH promoter-driven DNIκBα vs. Control, and subsequently these animals were treated with fast optogenetic stimulation vs. sham as described in Fig. 5 and profiled for physiology. (a–c) Longitudinal body weight (a) and food intake (b) follow-up and body composition measured at the age of about 18 months (c). (d–m) Aging-related physiology including behavioral functions assessed when 18–19 months old (d–j) and tissue histology assessed when 24 months old (k–m). P values are shown in figures for comparisons between groups as indicated, one-way ANOVA with Tukey’s post hoc test (d–m); n = 10 mice per group (a–b), n = 7 mice per group (c), n = 8 mice per group (d–m). Data are mean ± s.e.m.
Supplementary information
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Rights and permissions
About this article

Cite this article
Wang, Z., Wu, W., Kim, M.S. et al. GnRH pulse frequency and irregularity play a role in male aging. Nat Aging 1, 904–918 (2021). https://doi.org/10.1038/s43587-021-00116-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s43587-021-00116-5
This article is cited by
-
GnRH pulses affect ageing in male mice
Nature Reviews Endocrinology (2022)
